Use of Autochthonous Lactobacilli to Increase the Safety of Zgougou

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Selected Autochthonous Lactic Acid Bacteria for Fermentation of Aleppo Pine’s Seeds
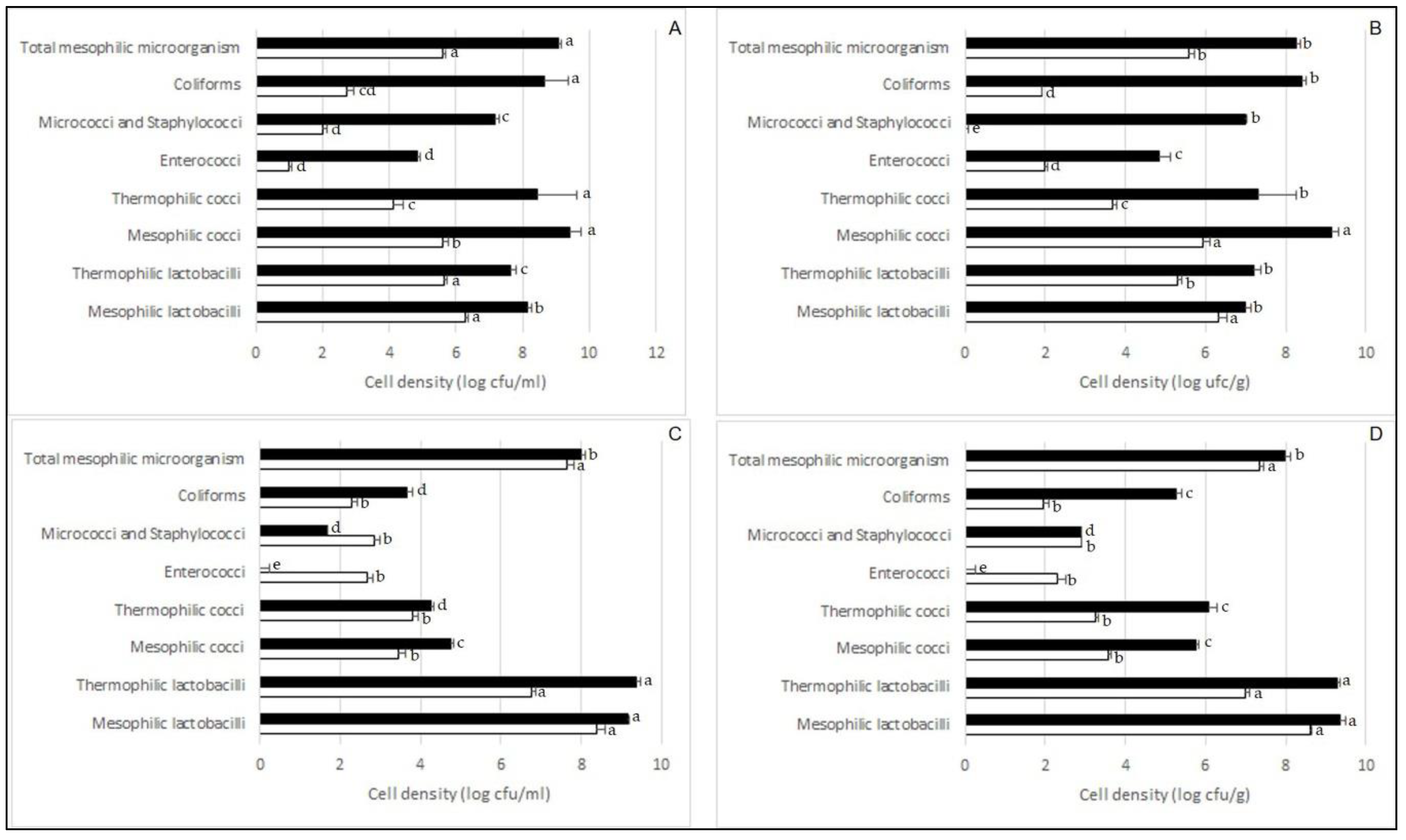
2.2. Cultivable Microbiota of Juice and Seeds from Fermented Zgougou
2.3. Culture-Independent Analysis of Microbial Community of Fermented Zgougou
2.4. VOC Profile of Zgougou
2.5. Correlations between Bacterial Biota and VOC
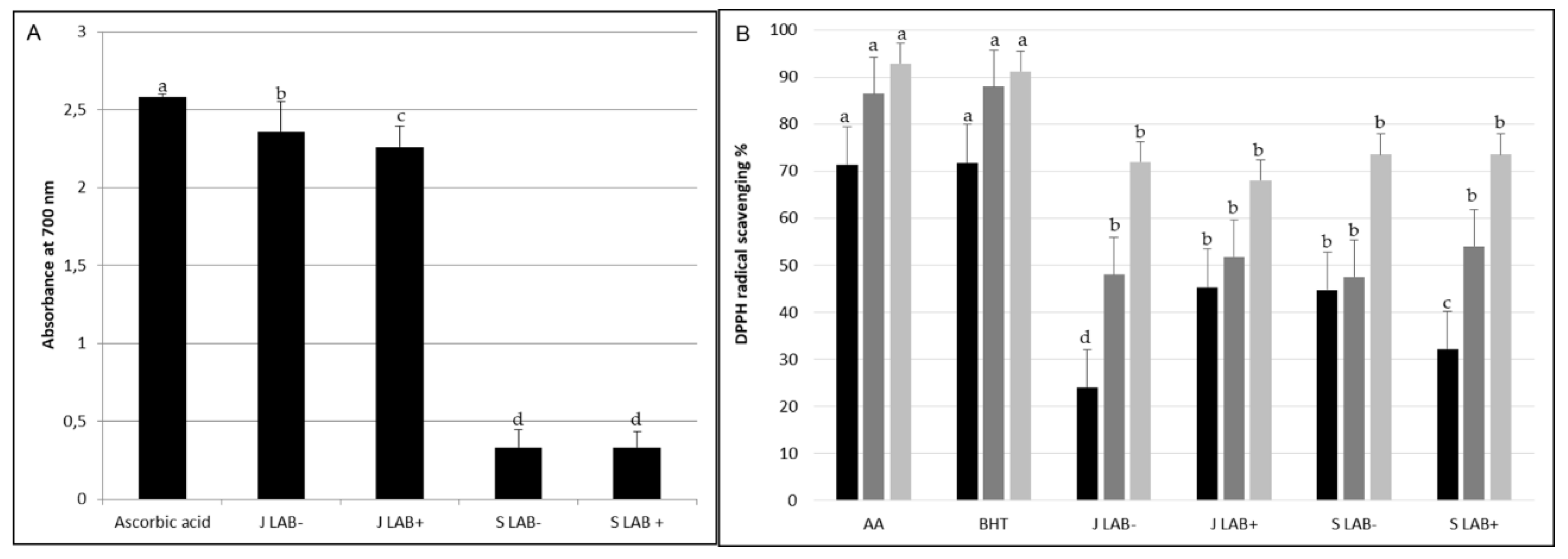
2.6. Antibacterial Activity of Zgougou Juice and Seeds
2.7. Antioxidant Activity of Zgougou
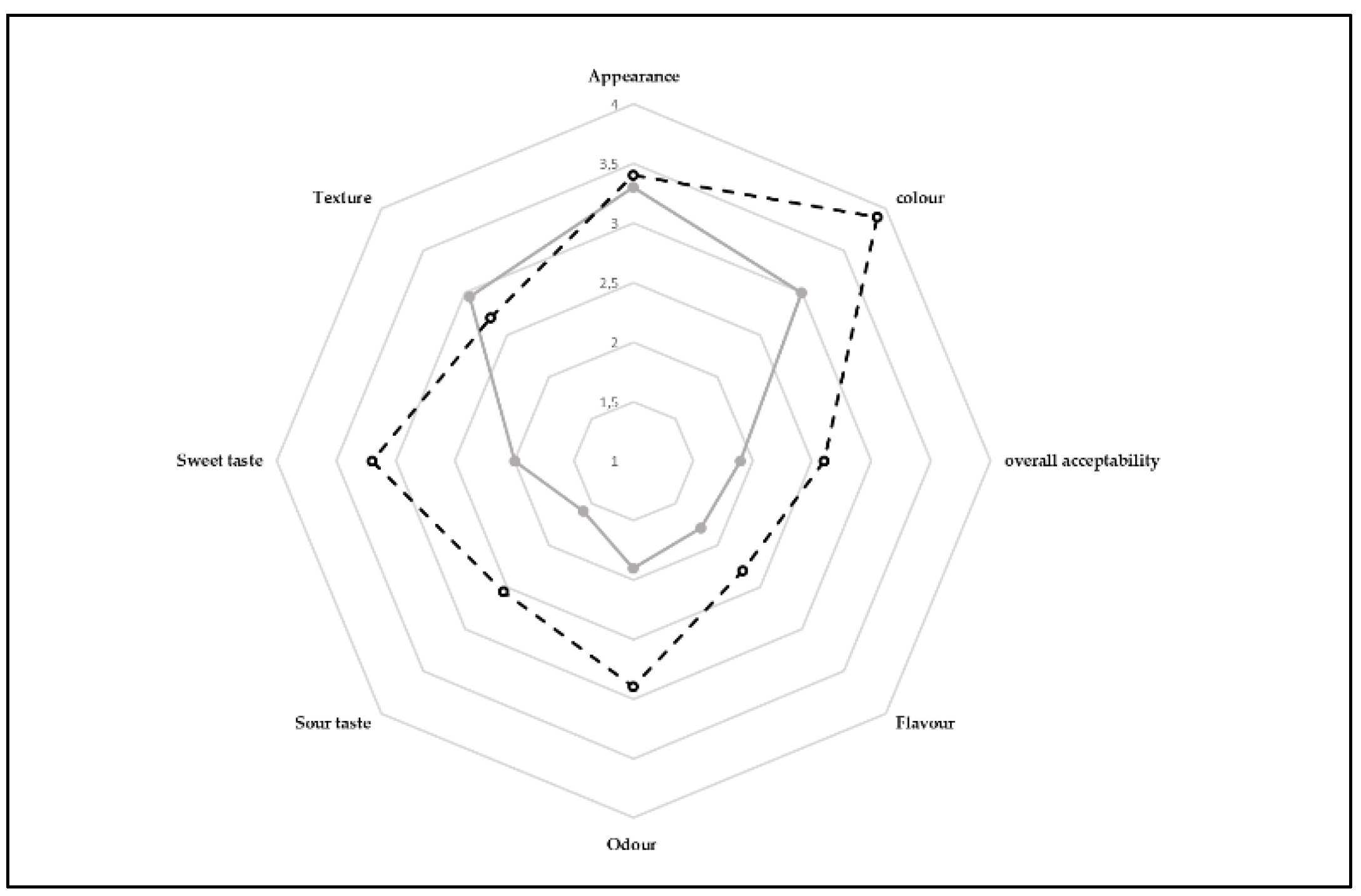
2.8. Sensory Analysis of Assidat-Zgougou Puddings
3. Discussion
4. Materials and Methods
4.1. Microorganisms, Culture Conditions and Acidification Capacity
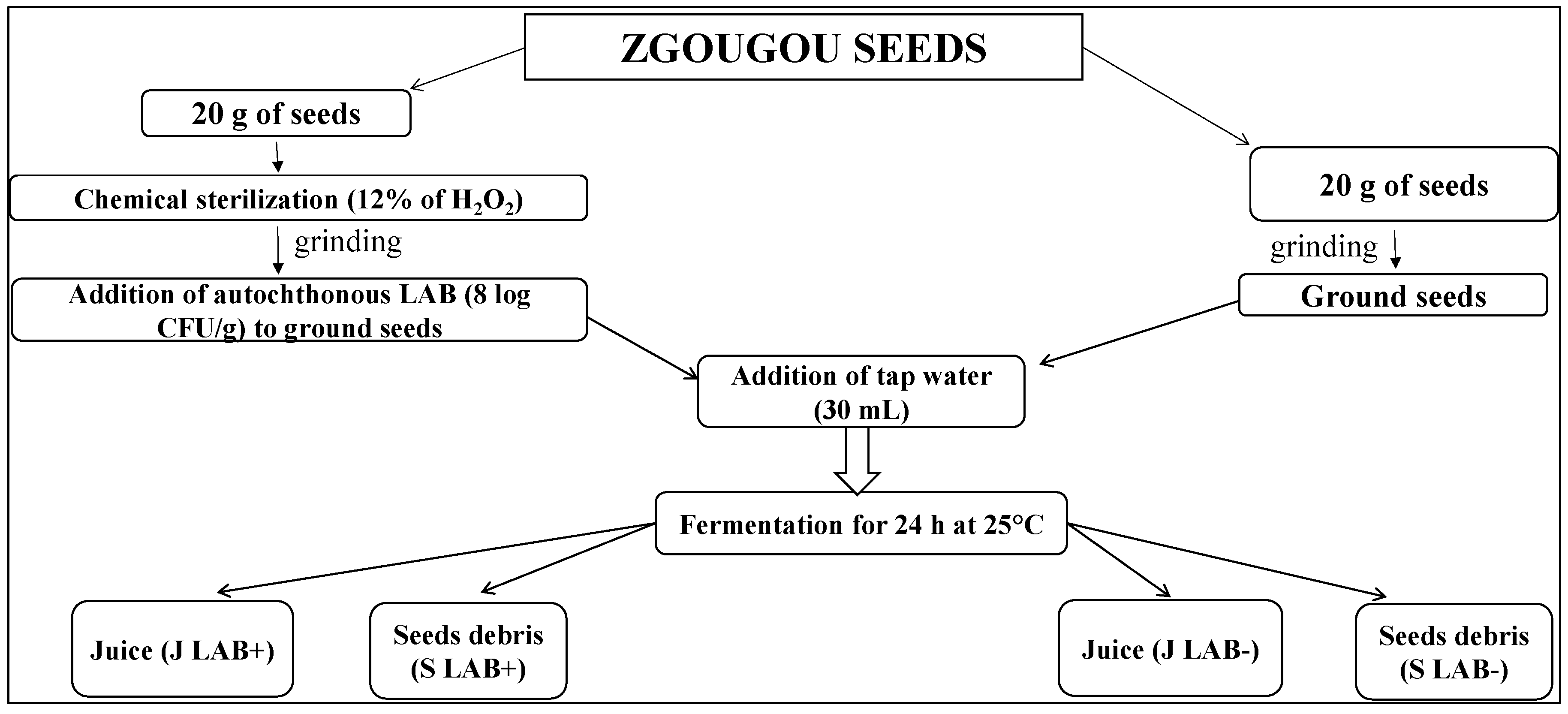
4.2. Protocol of Production of Zgougou
4.3. Cultivable Microbiota
4.4. Culture-Independent Analysis of Bacterial and Fungal Microbiota
4.5. VOCs Analyses
4.6. Antibacterial Activity
4.7. Determination of Antioxidant Activity
4.8. Sensory Analyses
4.9. Statistical Analyses
5. Conclusion and Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tsitsoni, T.K. Seed quality characteristics of Pinus halepensis–seed germination strategy and early seedling growth. Web. Ecol. 2009, 9, 72–76. [Google Scholar] [CrossRef]
- Fekih, N.; Allali, H.; Merghache, S.; Chaïb, F.; Merghache, D.; El Amine, M.; Djabou, N.; Muselli, A.; Tabti, B.; Costa, J. Chemical composition and antibacterial activity of Pinus halepensis Miller growing in West Northern of Algeria. Asian Pac. J. Trop. Dis. 2014, 4, 97. [Google Scholar] [CrossRef]
- Way, S. Strategic Management of Aleppo Pines on Lower Eyre Peninsula to Maximize Biodiversity Conservation Outcomes; Department for Environment and Heritage: Port Lincoln, Australia, 2006; p. 54. Available online: https://www.yumpu.com/en/document/read/10132669/strategic-management-of-aleppo-pines-on-lower-eyre/2 (accessed on 21 December 2019).
- Jaouadi, W.; Naghmouchi, S.; Alsubeie, M. Should the silviculture of Aleppo pine (Pinus halepensis Mill.) stands in northern Africa be oriented towards wood or seed and cone production? Diagnosis and current potentiality. IForest 2019, 12, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Ben Hamouda, M.; Ben Youssed, S.; Bedhiaf, S.; Annabi, M.; Draief, Z.; Gharbi, M.S. Status of Agricultural Innovations, Innovation Platforms, and Innovations Investment; 2015 PARI project country report: Republic of Tunisia. Forum for Agricultural Research in Africa (FARA): Accra, Ghana, 2016. Available online: https://research4agrinnovation.org/wp-content/uploads/2017/01/Tunisia_InnovationStudy.pdf (accessed on 21 December 2019).
- Nasri, N.; Triki, S. Analyse des lipides des graines de pins de Tunisie (Pinus halepensis Mill. et Pinus pinea L.). Riv. Ital. Sostanze Gr. 2004, 81, 244–247. [Google Scholar]
- Cheikh-Rouhou, S.; Hentati, B.; Besbes, S.; Blecker, C.; Deroanne, C.; Attia, H. Chemical composition and lipid fraction characteristics of Aleppo pine (Pinus halepensis Mill.) seeds cultivated in Tunisia. Food Sci. Technol. Int. 2006, 12, 407–415. [Google Scholar] [CrossRef]
- Soccol, C.R.; Vandenberghe, L.P.D.; Spier, M.R.; Medeiros, A.B.P.; Yamaguishi, C.T.; Lindner, J.D.; Pandey, A.; Thomaz-Soccol, V. The potential of probiotics: A review. Food Technol. Biotechnol. 2010, 48, 413–434. [Google Scholar]
- Hashemi, S.M.B.; Mousavi-Khaneghah, A.; Kontominas, M.G.; Eş, I.; Sant’Ana, A.S.; Martinez, R.R.; Drider, D. Fermentation of sarshir (kaymak) by lactic acid bacteria: Antibacterial activity, antioxidant properties, lipid and protein oxidation and fatty acid profile. J. Sci. Food Agric. 2017, 97, 4595–4603. [Google Scholar] [CrossRef]
- Trabelsi, I.; Ktari, N.; Slima, S.B.; Triki, M.; Bardaa, S.; Mnif, H.; Salah, R.B. Evaluation of dermal wound healing activity and in vitro antibacterial and antioxidant activities of a new exopolysaccharide produced by Lactobacillus sp. Ca6. Int. J. Biol. Macromol. 2017, 103, 194–201. [Google Scholar] [CrossRef]
- Son, S.H.; Yang, S.J.; Jeon, H.L.; Yu, H.S.; Lee, N.K.; Park, Y.S.; Paik, H.D. Antioxidant and immunostimulatory effect of potential probiotic Lactobacillus paraplantarum SC61 isolated from Korean traditional fermented food, jangajji. Microb. Pathog. 2018, 125, 486–492. [Google Scholar] [CrossRef]
- Fuchs-Tarlovsky, V.; Marquez-Barba, M.F.; Sriram, K. Probiotics in dermatologic practice. Nutr. J. 2016, 32, 289–295. [Google Scholar] [CrossRef]
- Friedrich, A.D.; Paz, M.L.; Leoni, J.; Gonzalez Maglio, D.H. Message in a bottle: Dialog be-tween intestine and skin modulated by probiotics. Int. J. Mol. Sci. 2017, 18, 1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aponte, M.; Blaiotta, G.; Croce, F.L.; Mazzaglia, A.; Farina, V.; Settanni, L.; Moschetti, G. Use of selected autochthonous lactic acid bacteria for Spanish-style table olive fermentation. Food Microbiol. 2011, 30, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Luo, D.; Bao, Y.; Liao, X.; Wu, J. Characterization of diversity and probiotic efficiency of the autochthonous lactic acid bacteria in the fermentation of selected raw fruit and vegetable juices. Front. Microbiol. 2018, 9, e2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorov, S.D.; Holzapfel, W.H. Traditional cereal fermented foods as source of functional organisms. In Advances in Food and Beverage Fermentations; Holzapfel, W.H., Ed.; Woodhead Publishing: Cambridge, UK, 2015; pp. 123–152. [Google Scholar]
- Celano, G.; De Angelis, M.; Minervini, F.; Gobbetti, M. Different flour microbial communities drive to sourdoughs characterized by diverse bacterial strains and free amino acid profiles. Front Microbiol. 2016, 7, 1770. [Google Scholar] [CrossRef] [Green Version]
- Missaoui, J.; Saidane, D.; Mzoughi, R.; Minervini, F. Fermented seeds (“Zgougou”) from Aleppo pine as novel source of potentially probiotic lactic acid bacteria. Microorganisms 2019, 7, 709. [Google Scholar] [CrossRef] [Green Version]
- Botella, L.; Diez, J.J. Phylogenic diversity of fungal endophytes in Spanish stands of Pinus halepensis. Fungal Divers. 2011, 47, 9–18. [Google Scholar] [CrossRef]
- Sanni, A.I. The need for process optimization of African fermented foods and beverages. Int. J. Food Microbiol. 1993, 18, 85–95. [Google Scholar] [CrossRef]
- Steinkraus, K.H. Classification of fermented foods: Worldwide review of household fermentation techniques. Food Control 1997, 8, 311–317. [Google Scholar] [CrossRef]
- Di Cagno, R.; Surico, R.F.; Siragusa, S.; De Angelis, M.; Paradiso, A.; Minervini, F.; De Gara, L.; Gobbetti, M. Selection and use of autochthonous mixed starter for lactic acid fermentation of carrots, French beans or marrows. Int. J. Food Microbiol. 2008, 127, 220–228. [Google Scholar] [CrossRef]
- Minervini, F.; Conte, A.; Del Nobile, M.A.; Gobbetti, M.; De Angelis, M. Dietary fibers and protective lactobacilli drive burrata cheese microbiome. Appl. Environ. Microbiol. 2017, 83, e01494-17. [Google Scholar] [CrossRef] [Green Version]
- Jackman, P.J.H.; Phillips, B.A.; Farrow, J.A.E.; Painting, K.A. National Collection of Food Bacteria Catalogue of Cultures; Agricultural and Food Research Council (AFRC): Reading, UK, 1990. [Google Scholar]
- Chaidoutis, E.; Migdanis, A.; Keramydas, D.; Papalexis, P. Biogenic amines in food as a public health concern; An outline of histamine food poisoning. Arch. Hell. Med. 2019, 36, 419–425. [Google Scholar]
- Cooney, S.; O’Brien, S.; Iversen, C.; Fanning, S. Other pathogenic Enterobacteriaceae–Enterobacter and other genera. In Encyclopedia of Food Safety; Motarjemi, Y., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 433–441. [Google Scholar]
- Hoel, S.; Vadstein, O.; Jakobsen, A.N. The significance of mesophilic Aeromonas spp. in minimally processed ready-to-eat seafood. Microorganisms 2019, 7, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahne, J.; Isele, D.; Berning, J.; Lipski, A. The contribution of fast growing, psychrotrophic microorganisms on biodiversity of refrigerated raw cow’s milk with high bacterial counts and their food spoilage potential. Food Microbiol. 2019, 79, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Babapour, E.; Haddadi, A.; Mirnejad, R.; Angaji, S.A.; Amirmozafari, N. Biofilm formation in clinical isolates of nosocomial Acinetobacter baumannii and its relationship with multidrug resistance. Asian Pac. J. Trop. Biomed. 2016, 6, 528–533. [Google Scholar] [CrossRef] [Green Version]
- Amorim, A.M.; Nascimento, J.D. Acinetobacter: An underrated foodborne pathogen? J. Infect. Dev. Ctries. 2017, 11, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Xin, K.; Liu, H.; Cheng, J.; Shen, X.; Wang, Y.; Zhang, L. Pantoea alhagi, a novel endophytic bacterium with ability to improve growth and drought tolerance in wheat. Sci. Rep. 2017, 7, 41564. [Google Scholar] [CrossRef] [Green Version]
- Pico, J.; Bernal, J.; Gómez, M. Wheat bread aroma compounds in crumb and crust: A review. Food Res. Int. 2015, 75, 200–215. [Google Scholar] [CrossRef]
- Park, M.K.; Kim, Y.S. Distinctive formation of volatile compounds in fermented rice inoculated by different molds, yeasts, and lactic acid bacteria. Molecules 2019, 24, 2123. [Google Scholar] [CrossRef] [Green Version]
- Mukisa, I.M.; Byaruhanga, Y.B.; Muyanj, C.M.B.K.; Langsrud, T.; Narvhus, J.A. Production of organic flavor compounds by dominant lactic acid bacteria and yeasts from Obushera, a traditional sorghum malt fermented beverage. Food Sci. Nutr. 2016, 5, 702–712. [Google Scholar] [CrossRef]
- Amri, I.; Hamrouni, L.; Hanana, M.; Gargouri, S.; Fezzani, T.; Jamoussi, B. Chemical composition, phytotoxic and antifungal activities of Pinus pinea essential oil. Biol. Agric. Hortic. 2013, 29, 91–106. [Google Scholar] [CrossRef]
- Food and Drugs Administration. Code of Federal Regulations-Title 21-Food and Drugs. Available online: https://www.fda.gov/medical-devices/medical-device-databases/code-federal-regulations-title-21-food-and-drugs (accessed on 21 December 2019).
- Sidira, M.; Kandylis, P.; Kanellaki, M.; Kourkoutas, Y. Effect of immobilized Lactobacillus casei on the evolution of flavor compounds in probiotic dry-fermented sausages during ripening. Meat Sci. 2015, 100, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Axelsson, L. Lactic acid bacteria: Classification and physiology. In Lactic Acid Bacteria: Microbiological and Functional Aspects, 3rd ed.; Salminen, S., Wright, A.V., Ouwehand, A., Eds.; Marcel Dekker: New York, NY, USA, 2004; pp. 1–67. [Google Scholar]
- Reis, J.A.; Paula, A.T.; Casarotti, S.N.; Penna, A.L.B. Lactic acid bacteria antimicrobial compounds: Characteristics and applications. Food Eng. Rev. 2012, 4, 124–140. [Google Scholar] [CrossRef]
- Jung, S.; Hwang, H.; Lee, J.H. Effect of lactic acid bacteria on phenyllactic acid production in kimchi. Food Control 2019, 106, 106701. [Google Scholar] [CrossRef]
- da Silva-Sabo, S.; Vitolo, M.; Domínguez-González, J.M.; de Souza-Oliveira, R.P. Overview of Lactobacillus plantarum as a promising bacteriocin producer among lactic acid bacteria. Food Res. Int. 2014, 64, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Djerrad, Z.; Kadik, L.; Djouahri, A. Chemical variability and antioxidant activities among Pinus halepensis Mill. essential oils provenances, depending on geographic variation and environmental conditions. Ind. Crops Prod. 2015, 74, 440–449. [Google Scholar] [CrossRef]
- Subedi, L.; Timalsena, S.; Duwadi, P.; Thapa, R.; Paudel, A.; Parajuli, K. Antioxidant activity and phenol and flavonoid contents of eight medicinal plants from Western Nepa. J. Tradit. Chin. Med. 2014, 34, 584–590. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Hayakawa, S.; Izumori, K. Modification of ovalbumin with a rare ketohexose through the Maillard reaction: effect on protein structure and gel properties. J. Agric. Food Chem. 2004, 52, 1293–1299. [Google Scholar] [CrossRef]
- Lattanzi, A.; Minervini, F.; Di Cagno, R.; Diviccaro, A.M.; Antonielli, L.; Cardinali, G.; Cappelle, S.; De Angelis, M.; Gobbetti, M. The lactic acid bacteria and yeast microbiota of eighteen sourdoughs used for the manufacture of traditional Italian sweet leavened baked goods. Int. J. Food Microbiol. 2013, 163, 71–79. [Google Scholar] [CrossRef]
- Handl, S.; Dowd, S.E.; Garcia-Mazcorro, J.F.; Steiner, J.M.; Suchodolski, J.S. Massive parallel 16S rRNA gene pyrosequencing reveals highly diverse fecal bacterial and fungal communities in healthy dogs and cats. FEMS Microbiol. Ecol. 2011, 76, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Lucero, M.E.; Unc, A.; Cooke, P.; Dowd, S.; Sun, S. Endophyte microbiome diversity in micropropagated Atriplex canescens and Atriplex torreyi var griffithsii. PLoS ONE 2011, 6, e17693. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd merger. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Cardinali, G.; Rizzello, C.G.; Buchin, S.; De Angelis, M.; Gobbetti, M.; Di Cagno, R. Metabolic responses of Lactobacillus plantarum strains during fermentation and storage of vegetable and fruit juices. Food Microbiol. 2014, 46, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Vitellio, P.; Celano, G.; Bonfrate, L.; Gobbetti, M.; Portincasa, P.; De Angelis, M. Effects of Bifidobacterium longum and Lactobacillus rhamnosus on gut microbiota in patients with lactose intolerance and persisting functional gastrointestinal symptoms: A randomised, double-blind, cross-over study. Nutrients 2019, 11, 886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuanxia, S.; Shigeru, H.; Melin, C.; Machiko, F.; Aree, I.; Ken, I. Antioxidant effects of Maillard reaction products obtained from ovalbumin and different d-aldohexoses. Biosci. Biotechnol. Biochem. 2006, 70, 598–605. [Google Scholar]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Das, D.; Goyal, A. Antioxidant activity and γ-aminobutyric acid (GABA) producing ability of probiotic Lactobacillus plantarum DM5 isolated from Marcha of Sikkim. Technologie 2015, 61, 263–268. [Google Scholar] [CrossRef]
- Bersuder, P.; Hole, M.; Smith, G. Antioxidants from a heated histidine-glucose model system. I: Investigation of the antioxidant role of histidine and isolation of antioxidants by high-performance liquid chromatography. J. Am. Oil Chem. Soc. 1998, 75, 181–187. [Google Scholar] [CrossRef]
- Haglund, A.; Johansson, L.; Berglund, L.; Dahlstedt, L. Sensory evaluation of carrots from ecological and conventional growing systems. Food Qual. Prefer. 1998, 10, 23–29. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | pH |
|---|---|
| Lactobacillus paraplantarum A1 | 4.65 ± 0.02f |
| Lactobacillus plantarum A2 | 4.65 ± 0.01f |
| L. plantarum A3 | 4.75 ± 0.03e |
| Enterococcus faecalis A4 | 4.84 ± 0.02c |
| Ent. faecalis A5 | 4.79 ± 0.02de |
| Ent. faecalis A6 | 4.89 ± 0.01b |
| Ent. faecalis A7 | 4.96 ± 0.01a |
| Ent. faecalis A8 | 4.83 ± 0.02c |
| Ent. faecalis A9 | 4.85 ± 0.03c |
| Ent. faecalis A10 | 4.76 ± 0.02e |
| Ent. faecalis A11 | 4.92 ± 0.03ab |
| Ent. faecalis A12 | 4.80 ± 0.01d |
| Ent. faecalis A13 | 4.85 ± 0.02c |
| Ent. faecalis A14 | 4.93 ± 0.02a |
| Sample | Observed Species | Chao1 | Shannon |
|---|---|---|---|
| J LAB- | 17.0 ± 1.41c | 17.3 ± 1.65c | 1.71 ± 0.032c |
| S LAB- | 21.5 ± 0.71a | 21.5 ± 0.71a | 2.06 ± 0.004a |
| J LAB+ | 3.0 ± 1.41d | 3.0 ± 1.41d | 0.44 ± 0.021d |
| S LAB+ | 20.0 ± 0.00b | 20.0 ± 0.00b | 1.91 ± 0.030b |
| Bacterial OTUs | J LAB− | S LAB− | J LAB+ | S LAB+ |
|---|---|---|---|---|
| Bacillus cereus | 0.22 ± 0.02b | 0.55 ± 0.34b | 0c | 0.99 ± 0.06a |
| Enterococcus hirae | 0.87 ± 0.23c | 1.60 ± 0.53b | 0c | 2.75 ± 0.48a |
| Lactobacillus plantarum | 43.83 ± 1.39b | 2.21 ± 0.95d | 82.89 ± 1.15a | 6.20 ± 0.19c |
| Weissella confusa | 0b | 0b | 0b | 9.34 ± 0.40a |
| Lactococcus raffinolactis | 3.12 ± 0.34b | 34.74 ± 0.82a | 0c | 0c |
| Bacilli | 0.004 ± 0.00b | 0.02 ± 0.00a | 0b | 0b |
| Clostridiales | 0a | 0.02 ± 0.00a | 0a | 0a |
| Aeromonas sp | 1.43 ± 0.14b | 2.75 ± 0.05a | 0d | 0.38 ± 0.10c |
| Enterobacter sp | 28.43 ± 0.36b | 37.68 ± 1.45a | 0d | 25.20 ± 0.47c |
| Pantoea sp. | 0.01 ± 0.00b | 0.08 ± 0.01b | 0b | 15.55 ± 1.09a |
| Enterobacteriaceae | 0.71 ± 0.04b | 1.28 ± 0.05a | 0c | 0.68 ± 0.03b |
| Acinetobacter junii | 0.06 ± 0.02b | 0.07 ± 0.00b | 0c | 0.14 ± 0.09a |
| Acinetobacter radioresistens | 0a | 0a | 0a | 0.01 ± 0.00a |
| Acinetobacter sp | 1.92 ± 0.11b | 5.31 ± 0.09a | 0d | 0.58 ± 0.18c |
| Pseudomonas sp | 1.41 ± 0.03a | 0.02 ± 0.01c | 0d | 0.64 ± 0.62b |
| Gammaproteobacteria | 0c | 0.06 ± 0.09b | 0c | 0.25 ± 0.01a |
| Other unclassified bacteria | 17.93 ± 0.93b | 13.54 ± 1.23c | 17.1 ± 1.16b | 37.22 ± 2.29a |
| Compounds | Odor | J LAB− | J LAB+ | S LAB− | S LAB+ |
|---|---|---|---|---|---|
| Alcohols | |||||
| Ethanol | Sweet, alcoholic, ripe apple | 0.32 ± 0.142a | 0.05 ± 0.001b | 0.08 ± 0.021A | 0.01 ± 0.003B |
| 1-butanol 3-methyl | Fermented, malt, wine | 0.03 ± 0.016a | 0.01 ± 0.003a | NdA | NdA |
| 3-butoxy-1-propanol | Nf | 0.02 ± 0.009a | 0.03 ± 0.004a | NdB | 0.10 ± 0.026A |
| 1-hexanol | Green grass, flowery, woody, mild, sweet | 0.04 ± 0.021b | 0.09 ± 0.007a | 0.14 ± 0.046A | 0.19 ± 0.003A |
| 3,5-octadien-2-ol | Bean-like | Nda | Nda | 0.05 ± 0.016A | NdB |
| 1-nonanol | Ophrys | Ndb | 0.02 ± 0.000a | NdB | 0.03 ± 0.008A |
| Benzyl alcohol | Boiled cherries, moss, roasted bread, rose | 0.01 ± 0.007a | 0.01 ± 0.002a | NdA | NdA |
| Phenylethyl alcohol | Rose-honey-like, wilted rose | 0.02 ± 0.010a | 0.01 ± 0.001a | 0.04 ± 0.016A | 0.03 ± 0.007A |
| Aldehydes | |||||
| Acetaldehyde | Fruity, floral, green apple, nut, penetrating | 0.01 ± 0.005a | Ndb | 0.14 ± 0.032A | 0.01 ± 0.001B |
| Octane, 4-methyl- | Nf | 0.01 ± 0.008b | 0.02 ± 0.002a | NdB | 0.02 ± 0.003A |
| Butanal, 3-methyl- | Malty, roasty cucumber-like | Nda | Nda | 0.02 ± 0.006A | NdB |
| Pentanal | Nf | Nda | Nda | 0.03 ± 0.011A | 0.03 ± 0.014A |
| Hexanal | Green, grassy, tallow | Nda | Nda | 0.26 ± 0.103A | 0.46 ± 0.288A |
| Heptanal | Fatty, rancid, citrus, malty | Nda | Nda | 0.02 ± 0.010A | 0.03 ± 0.008A |
| Octanal | Aglaia, Cymbidium, Hydnora, Ophry | Nda | Nda | 0.10 ± 0.052A | 0.08 ± 0.037A |
| Nonanal | Fat, floral, green, lemon, paint | Nda | Nda | 0.21 ± 0.096A | 0.14 ± 0.059A |
| 2-octenal | Fat, fish oil, green, nut, plastic | 0.02 ± 0.010a | 0.05 ± 0.001a | 0.22 ± 0.080A | 0.26 ± 0.067A |
| 2-furaldehyde | Almond-like | 0.03 ± 0.013a | 0.04 ± 0.005a | 0.22 ± 0.062A | 0.29 ± 0.068A |
| Decanal | Stewed, burnt, green, waxy, floral, lemon | 0.04 ± 0.017a | 0.02 ± 0.002a | 0.09 ± 0.032A | 0.05 ± 0.013A |
| Benzaldehyde | Almond, caramel | 0.02 ± 0.027a | 0.03 ± 0.010a | 0.10 ± 0.042A | 0.10 ± 0.022A |
| 2-nonenal | Fatty, tallowy, green | 0.02 ± 0.009a | 0.03 ± 0.004a | 0.07 ± 0.028A | 0.05 ± 0.012A |
| Decenal | tallow | Nda | Nda | 0.07 ± 0.020A | 0.07 ± 0.016A |
| 2-undecenal | sweet | Nda | Nda | 0.01 ± 0.005A | 0.01 ± 0.002A |
| 2,4-decadienal | seaweed | Ndb | 0.04 ± 0.010a | 0.09 ± 0.024A | 0.11 ± 0.033A |
| Benzeneacetaldehyde | Berry, geranium, honey, nut, pungent | 0.01 ± 0.007a | Ndb | NdB | 0.01 ± 0.003A |
| Alkanes | |||||
| Decane | Nf | 0.02 ± 0.009b | 0.06 ± 0.017a | 0.04 ± 0.007B | 0.07 ± 0.015A |
| Undecane | Nf | 0.08 ± 0.029a | 0.08 ± 0.006a | NdA | NdA |
| Nonane, 4,5-dimethyl- | Nf | 0.03 ± 0.009a | 0.03 ± 0.004a | NdA | NdA |
| Decane 2,4,6 dimethyl | Nf | 0.19 ± 0.074a | 0.24 ± 0.044a | 0.15 ± 0.072A | 0.15 ± 0.037A |
| Tricyclo[3.2.1.0(2,4)]octane, | Nf | 0.03 ± 0.024a | 0.04 ± 0.005a | NdA | NdA |
| Dodecane | Nf | 0.01 ± 0.016a | 0.01 ± 0.007a | NdA | NdA |
| Dodecane,4,6-dimethyl- | Nf | 0.14 ± 0.062a | 0.17 ± 0.021a | NdB | 0.03 ± 0.004A |
| Heptadecane, 2,6,10,15-tetramethyl- | Nf | 0.01 ± 0.004a | 0.01 ± 0.001a | NdA | NdA |
| Cyclopentane,1-ethenyl-3-ethyl-2-methyl | Nf | 0.034 ± 0.013b | 0.06 ± 0.007a | 0.13 ± 0.039A | 0.15 ± 0.038A |
| Aromatic Compounds | |||||
| Furan, 2-pentyl- | Butter, green bean, floral, fruity, mushroom, raw nuts | 0.02 ± 0.007a | 0.02 ± 0.003a | 0.09 ± 0.034A | 0.06 ± 0.016A |
| O-cymene | Citrus-like, solvent, gasoline | 0.04 ± 0.018a | 0.04 ± 0.005a | 0.09 ± 0.016A | 0.05 ± 0.024A |
| Estragole | Tarragon | Nda | Nda | 0.03 ± 0.014A | 0.01 ± 0.004A |
| Carboxylic Acids | |||||
| Acetic acid | Sour, nutty | Ndb | 0.20 ± 0.023a | 0.16 ± 0.030B | 0.24 ± 0.055A |
| Butanoic acid | cheese, unpleasant | Ndb | 0.01 ± 0.004a | NdA | NdA |
| Pentanoic acid | Sweet, rancid | Nda | Nda | 0,03 ± 0.007B | 0.05 ± 0.002A |
| Hexanoic acid (caproic acid) | Sweaty, cheesy, fatty, goat-like | 0.03 ± 0.008b | 0.19 ± 0.021a | 0.63 ± 0.184A | 0.81 ± 0.185A |
| Heptanoic acid | Cheese, fatty, sweaty | Ndb | 0.02 ± 0.005a | 0.07 ± 0.016A | 0.08 ± 0.018A |
| Octanoic acid | Cheese, fatty, sweaty, soapy, chocolate-like | 0.02 ± 0.00b | 0.06 ± 0.015a | 0.05 ± 0.056A | 0.16 ± 0.040A |
| Nonanoic acid | Cheese, fatty, sweaty | 0.05 ± 0.038 a | 0.09 ± 0.046a | 0.16 ± 0.007B | 0.26 ± 0.036A |
| Esters | |||||
| Ethyl acetate | Caramel, sweet, fruity, acid, buttery, pineapple | 0.04 ± 0.018a | 0.02 ± 0.002b | NdA | NdA |
| Hexanoic acid, ethyl ester | Apple peel-like, fruity | 0.04 ± 0.000a | Ndb | NdA | NdA |
| Butanoic acid, pentyl ester | Nf | Nda | Nda | NdB | 0.02 ± 0.003A |
| Sulfurous acid, nonyl pentyl ester | Nf | Nda | Nda | 0.07 ± 0.023A | NdB |
| Octanoic acid, ethyl ester | Alcohol-like, Fruity, citrus-like | 0.03 ± 0.010a | Ndb | NdA | NdA |
| Hexanoic acid, etenhyl ester | Nf | 0.02 ± 0.010a | 0.0 ± 0.003a | 0.08 ± 0.025A | 0.10 ± 0.019A |
| Ketones | |||||
| 2-heptanone | Soapy, fruity, cinnamon | Nda | Nda | 0.02 ± 0.004B | 0.04 ± 0.004A |
| Acetoin | flowery, wet, butter, cream | 0.15 ± 0.025a | 0.11 ± 0.000b | 0.89 ± 0.259A | 0.07 ± 0.012B |
| 2-octanone | Gasoline, mould, soap | Ndb | 0.03 ± 0.009a | 0.04 ± 0.008A | 0.03 ± 0.005A |
| 2-undecanone | fresh, green | Nda | Nda | NdB | 0.03 ± 0.007A |
| Acetophenone | Cheesy, sweet, almond, floral | 0.02 ± 0.017 | Ndb | NdA | NdA |
| Others | |||||
| 1,3,5,7-cyclooctatetraene | Nf | 0.01 ± 0.002a | Nda | NdA | NdA |
| Indole | mothball, burnt | 0.01 ± 0.018a | Ndb | NdA | NdA |
| Phenols | |||||
| Phenol, 2-methoxy- | Phenol, | 0.43 ± 0.018a | 0.02 ± 0.008b | 0.54 ± 0.203A | 0.06 ± 0.016B |
| Phenol | Phenol | 0.05 ± 0.002a | 0.01 ± 0.003b | 0.08 ± 0.029A | NdB |
| P-cresol | medicine, phenol, smoke | 0.01 ± 0.000a | Ndb | NdA | NdA |
| Vanillin | Vanilla | Nda | Nda | NdB | 0.01 ± 0.004A |
| Terpenes | |||||
| α-pinene | Woody-spicy, oily, pine-like | 3.45 ± 1.392a | 3.21 ± 0.373a | 2.63 ± 0.707A | 2.56 ± 0.470A |
| β-pinene | resinous-piney, dry-terpenous | 0.01 ± 0.008a | 0.01 ± 0.005a | 0.04 ± 0.023A | 0.02 ± 0.005A |
| Camphene | Camphor | 0.04 ± 0.017a | 0.04 ± 0.003a | 0.03 ± 0.009A | 0.03 ± 0.004A |
| 3-carene | lemon, resin | 0.09 ± 0.036a | 0.09 ± 0.007a | 0.07 ± 0.017A | 0.07 ± 0.013A |
| β-myrcene | weak citrus and lime-like | 3.05 ± 1.216a | 3.07 ± 0.497a | 3.60 ± 0.898A | 3.63 ± 0.730A |
| D-limonene | Citrus, Licorice, citrus, green, fruity | 0.33 ± 0.145a | 0.28 ± 0.027a | 0.18 ± 0.061A | 0.22 ± 0.040A |
| β-phellandrene | citrus-like, weak herbal-spicy | 0.02 ± 0.035a | 0.03 ± 0.006a | NdB | 0.06 ± 0.007A |
| 4-methylisopropenyl-benzene | Nf | 0.02 ± 0.010a | 0.02 ± 0.003a | 0.03 ± 0.016A | 0.03 ± 0.006A |
| 1,7,7-trimethylbicyclo [2.2.1]heptane-2,5-diol | Nf | Ndb | 0.02 ± 0.001a | NdA | NdA |
| (+)-α-gurjunene | wood, balsamic | Ndb | 0.01 ± 0.014a | 0.05 ± 0.012A | NdB |
| Bornyl acetate | Nf | Ndb | 0.01 ± 0.001a | 0.01 ± 0.004B | 0.03 ± 0.008A |
| Caryophyllene | Dry, woody-spicy | 0.28 ± 0.000a | 0.05 ± 0.006b | 0.30 ± 0.104A | 0.07 ± 0.016B |
| Verbenol | Nf | 0.02 ± 0.000a | 0.01 ± 0.001a | 0.03 ± 0.008A | 0.03 ± 0.007A |
| Verbenone | Nf | 0.05 ± 0.076a | 0.04 ± 0.003a | 0.12 ± 0.046A | 0.06 ± 0.007A |
| Borneol | Pungent, mint | 0.02 ± 0.018a | 0.02 ± 0.000a | 0.02 ± 0.007A | 0.02 ± 0.013A |
| (-)-carvone | Nf | 0.01 ± 0.008a | Ndb | 0.03 ± 0.010A | 0.01 ± 0.005A |
| α.-farnesene | wood, sweet | Nda | Nda | NdB | 0.05 ± 0.002A |
| Bornyl formate | Green | Ndb | 0.01 ± 0.000a | NdB | 0.02 ± 0.013A |
| Target Bacterial Species | J LAB− | J LAB+ | S LAB− | S LAB+ |
|---|---|---|---|---|
| Staphylococcus aureus | − | + | − | + |
| Staphylococcus epidermidis | − | + | − | + |
| Micrococcus luteus | + | + | − | + |
| Escherichia coli | + | + | + | + |
| Listeria monocytogenes | − | ++ | + | + |
| Enterococcus faecalis | − | ++ | + | + |
| Salmonella Typhimurium | − | + | + | + |
| Bacilus cereus | + | + | − | + |
| Vibrio parahaemolyticus | − | ++ | + | + |
| Aeromonas hydrophila | − | + | − | − |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minervini, F.; Missaoui, J.; Celano, G.; Calasso, M.; Achour, L.; Saidane, D.; Gobbetti, M.; De Angelis, M. Use of Autochthonous Lactobacilli to Increase the Safety of Zgougou. Microorganisms 2020, 8, 29. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010029
Minervini F, Missaoui J, Celano G, Calasso M, Achour L, Saidane D, Gobbetti M, De Angelis M. Use of Autochthonous Lactobacilli to Increase the Safety of Zgougou. Microorganisms. 2020; 8(1):29. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010029
Chicago/Turabian StyleMinervini, Fabio, Jihen Missaoui, Giuseppe Celano, Maria Calasso, Lotfi Achour, Dalila Saidane, Marco Gobbetti, and Maria De Angelis. 2020. "Use of Autochthonous Lactobacilli to Increase the Safety of Zgougou" Microorganisms 8, no. 1: 29. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010029