Reuterin Demonstrates Potent Antimicrobial Activity Against a Broad Panel of Human and Poultry Meat Campylobacter spp. Isolates

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media and Growth Conditions
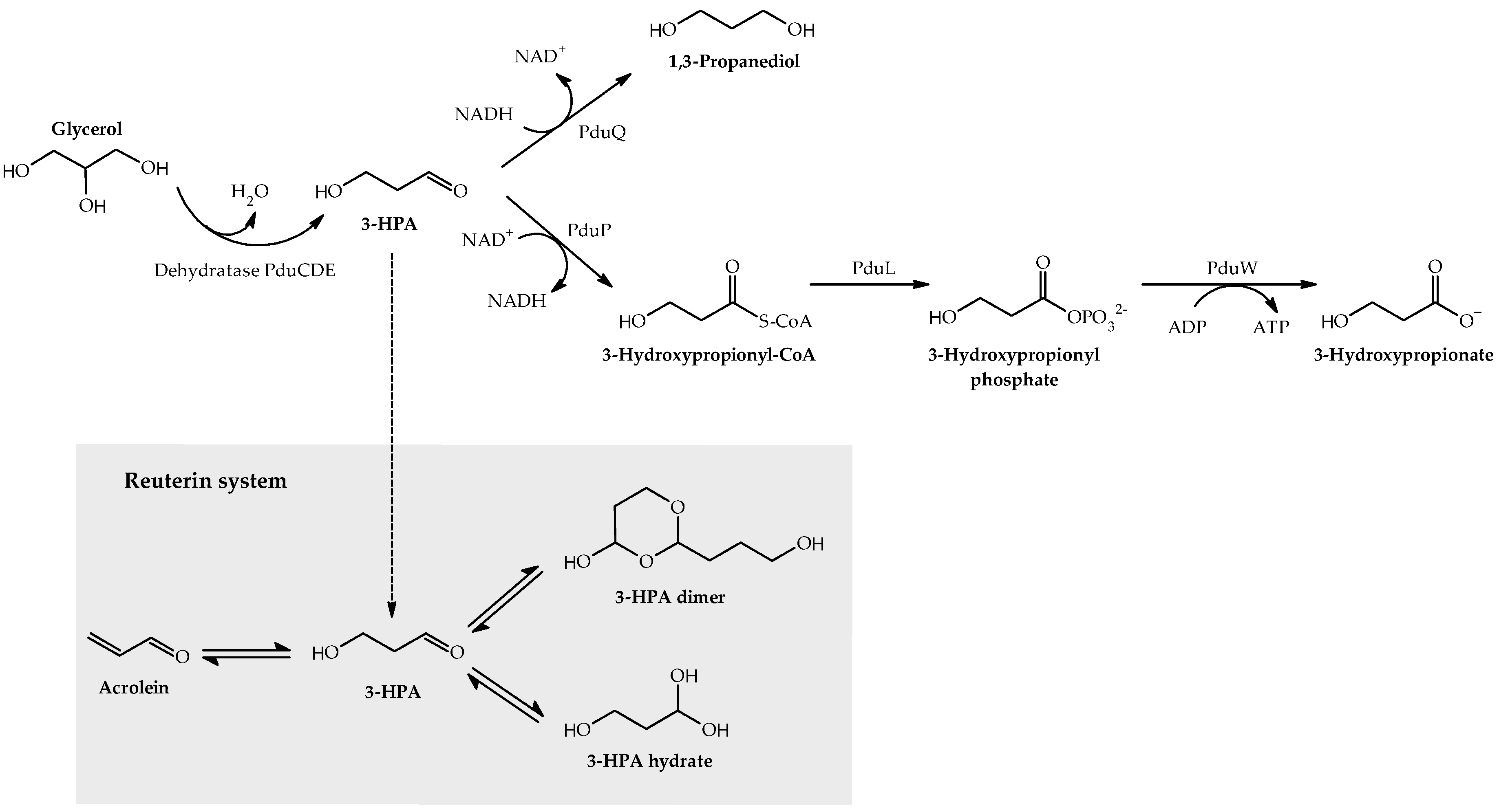
2.2. Reuterin Production
2.3. Reuterin Antimicrobial Activity Against a Panel of C. jejuni and C. coli Strains
2.4. Glycerol Metabolism by Mono-Culture Assay of L. reuteri and C. jejuni under Microaerophilic Growth Condition
2.5. Co-Culture Assays of L. reuteri and C. jejuni with Glycerol
2.6. DNA Isolation and Quantitative PCR (qPCR) Analysis
2.7. Chemical Analyses
2.8. Statistical Analysis
3. Results
3.1. C. jejuni and C. coli Strains are Highly Sensitive to Reuterin Compared to E. coli K12 Indicator
3.2. L. reuteri PTA5_F13 Produces Reuterin under Microaerophilic Growth Conditions
3.3. Reuterin Produced by L. reuteri PTA5_F13 Kills C. jejuni N16-1419 during Co-Cultures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- EFSA. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSA J. 2017, 15, 5077. [Google Scholar]
- EFSA. Scientific opinion on Campylobacter in broiler meat production: Control options and performance objectives and/or targets at different stages of the food chain. EFSA J. 2011, 9, 1–141. [Google Scholar]
- Nadeau, R.; Messier, S.; Quessy, S. Prevalence and comparison of genetic profiles of Campylobacter strains isolated from poultry and sporadic cases of campylobacteriosis in humans. J. Food Prot. 2002, 65, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehm, M.; Heimesaat, M.M. Chapter 1—Human Campylobacteriosis. In Campylobacter; Klein, G., Ed.; Academic Press: London, UK, 2017; pp. 1–25. [Google Scholar]
- Lehtopolku, M.; Nakari, U.M.; Kotilainen, P.; Huovinen, P.; Siitonen, A.; Hakanen, A.J. Antimicrobial susceptibilities of multidrug-resistant Campylobacter jejuni and C. coli strains: In vitro activities of 20 antimicrobial agents. Antimicrob. Agents Chemother. 2010, 54, 1232–1236. [Google Scholar] [CrossRef] [Green Version]
- Schiaffino, F.; Colston, J.M.; Paredes-Olortegui, M.; François, R.; Pisanic, N.; Burga, R.; Peñataro-Yori, P.; Kosek, M.N. Antibiotic resistance of Campylobacter species in a pediatric cohort study. Antimicrob. Agents Chemother. 2019, 63, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, K.; Wolkowicz, T.; Osek, J. Antimicrobial resistance and virulence-associated traits of Campylobacter jejuni isolated from poultry food chain and humans with diarrhea. Front. Microbiol. 2018, 9, 1508. [Google Scholar] [CrossRef]
- Morita, H.; Hidehiro, T.O.H.; Fukuda, S.; Horikawa, H.; Oshima, K.; Suzuki, T.; Murakami, M.; Hisamatsu, S.; Kato, Y.; Takizawa, T.; et al. Comparative genome analysis of Lactobacillus reuteri and Lactobacillus fermentum reveal a genomic island for reuterin and cobalamin production. DNA Res. 2008, 15, 151–161. [Google Scholar] [CrossRef]
- Engels, C.; Schwab, C.; Zhang, J.; Stevens, M.J.A.; Bieri, C.; Ebert, M.-O.; McNeill, K.; Sturla, S.J.; Lacroix, C. Acrolein contributes strongly to antimicrobial and heterocyclic amine transformation activities of reuterin. Sci. Rep. 2016, 6, 36246. [Google Scholar] [CrossRef] [Green Version]
- Stevens, M.; Vollenweider, S.; Lacroix, C. The potential of reuterin produced by Lactobacillus reuteri as a broad-spectrum preservative in food. In Protective Cultures, Antimicrobial Metabolites and Bacteriophages for Food and Beverage Biopreservation; Woodhead Publishing: Sawston, UK, 2011; pp. 129–160. [Google Scholar]
- Schaefer, L.; Auchtung, T.A.; Hermans, K.E.; Whitehead, D.; Borhan, B.; Britton, R.A. The antimicrobial compound reuterin (3-hydroxypropionaldehyde) induces oxidative stress via interaction with thiol groups. Microbiology 2010, 156, 1589–1599. [Google Scholar] [CrossRef] [Green Version]
- Asare, P.T.; Greppi, A.; Stettler, M.; Schwab, C.; Stevens, M.J.A.; Lacroix, C. Decontamination of minimally-processed fresh lettuce using reuterin produced by Lactobacillus reuteri. Front. Microbiol. 2018, 9, 1421. [Google Scholar] [CrossRef] [PubMed]
- Walter, J. Ecological role of Lactobacilli in the gastrointestinal tract: Implications for fundamental and biomedical research. Appl. Environ. Microbiol. 2008, 74, 4985–4996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dozier, I.W.A.; Kerr, B.J.; Branton, S.L. Apparent metabolizable energy of crude glycerin originating from different sources in broiler chickens. Poult. Sci. 2011, 90, 2528–2534. [Google Scholar] [CrossRef] [PubMed]
- Groesbeck, C.N.; McKinney, L.J.; DeRouchey, J.M.; Tokach, M.D.; Goodband, R.D.; Dritz, S.S.; Nelssen, J.L.; Duttlinger, A.W.; Fahrenholz, A.C.; Behnke, K.C. Effect of crude glycerol on pellet mill production and nursery pig growth performance. J. Anim. Sci. 2008, 86, 2228–2236. [Google Scholar] [CrossRef] [Green Version]
- Greppi, A.; Asare, P.T.; Schwab, C.; Zemp, N.; Stephan, R.; Lacroix, C. Isolation and comparative genomic analysis of reuterin-producing Lactobacillus reuteri from poultry gastrointestinal tract. bioRxiv 2019. [CrossRef] [Green Version]
- Oh, P.L.; Benson, A.K.; Peterson, D.A.; Patil, P.B.; Moriyama, E.N.; Roos, S.; Walter, J.; Lantbruksuniversitet, S. Diversification of the gut symbiont Lactobacillus reuteri as a result of host-driven evolution. ISME J. 2010, 4, 377–387. [Google Scholar] [CrossRef]
- Mota-meira, M.; LaPointe, G.; Lacroix, C.; Lavoie, M.C.; Pointe, L.E.L.A.; Lacroix, C.; Mota-meira, M.; Lavoie, M.C. MICs of mutacin B-Ny266, nisin A, vancomycin, and oxacillin against bacterial pathogens. Antimicrob. Agents Chemother. 2000, 44, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Cleusix, V.; Lacroix, C.; Vollenweider, S.; Duboux, M.; le Blay, G. Inhibitory activity spectrum of reuterin produced by Lactobacillus reuteri against intestinal bacteria. BMC Microbiol. 2007, 7, 101. [Google Scholar] [CrossRef] [Green Version]
- Davis, L.; DiRita, V. Growth and laboratory maintenance of Campylobacter jejuni. Curr. Protoc. Microbiol. 2008, 8, 1–7. [Google Scholar]
- Furet, J.-P.; Firmesse, O.; Gourmelon, M.; Bridonneau, C.; Tap, J.; Mondot, S.; Dore, J.; Corthier, G. Comparative assessment of human and farm animal faecal microbiota using real-time quantitative PCR. FEMS Microbiol. Ecol. 2009, 68, 351–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josefsen, M.H.; Jacobsen, N.R.; Hoorfar, J. Enrichment followed by quantitative PCR both for rapid detection and as a tool for quantitative risk assessment of food-borne thermotolerant Campylobacters. Appl. Environ. Microbiol. 2004, 70, 3588–3592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoddard, S.F.; Smith, B.J.; Hein, R.; Roller, B.R.K.; Schmidt, T.M. rrnDB: Improved tools for interpreting rRNA gene abundance in bacteria and archaea and a new foundation for future development. Nucleic Acids Res. 2015, 43, D593–D598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollenweider, S.; Grassi, G.; König, I.; Puhan, Z. Purification and structural characterization of 3-hydroxypropionaldehyde and its derivatives. J. Agric. Food Chem. 2003, 51, 3287–3293. [Google Scholar] [CrossRef]
- Saint-Cyr, M.J.; Guyard-Nicodème, M.; Messaoudi, S.; Chemaly, M.; Cappelier, J.M.; Dousset, X.; Haddad, N. Recent advances in screening of anti-Campylobacter activity in probiotics for use in poultry. Front. Microbiol. 2016, 7, 553. [Google Scholar] [CrossRef] [Green Version]
- Robin, D.D.; Sabrina, V.; Isabelle, D.; Christopher, M.; André, T.; Philippe, T. In vitro antagonistic activity evaluation of lactic acid bacteria (LAB) combined with cellulase enzyme against Campylobacter jejuni growth in co-culture. J. Microbiol. Biotechnol. 2011, 21, 62–70. [Google Scholar]
- Johnson, T.J.; Shank, J.M.; Johnson, J.G. Current and potential treatments for reducing Campylobacter colonization in animal hosts and disease in humans. Front. Microbiol. 2017, 8, 487. [Google Scholar] [CrossRef] [Green Version]
- Levesque, S.; Fournier, E.; Carrier, N.; Frost, E.; Arbeit, R.D.; Michaud, S. Campylobacteriosis in urban versus rural areas: A case-case study integrated with molecular typing to validate risk factors and to attribute sources of infection. PLoS ONE 2013, 8, 17–20. [Google Scholar] [CrossRef] [Green Version]
- Griekspoor, P.; Engvall, E.O.; Åkerlind, B.; Olsen, B.; Waldenström, J. Genetic diversity and host associations in Campylobacter jejuni from human cases and broilers in 2000 and 2008. Vet. Microbiol. 2015, 178, 94–98. [Google Scholar] [CrossRef]
- Arqués, J.L.; Fernández, J.; Gaya, P.; Nuñez, M.; Rodríguez, E.; Medina, M. Antimicrobial activity of reuterin in combination with nisin against food-borne pathogens. Int. J. Food Microbiol. 2004, 95, 225–229. [Google Scholar] [CrossRef]
- Vollenweider, S.; Evers, S.; Zurbriggen, K.; Lacroix, C. Unraveling the hydroxypropionaldehyde (HPA) system: An active antimicrobial agent against human pathogens. J. Agric. Food Chem. 2010, 58, 10315–10322. [Google Scholar] [CrossRef] [PubMed]
- Nunoshiba, T.; Yamamoto, K. Role of glutathione on acrolein-induced cytotoxicity and mutagenicity in Escherichia coli. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 1999, 442, 1–8. [Google Scholar] [CrossRef]
- Fouts, D.E.; Mongodin, E.F.; Mandrell, R.E.; Miller, W.G.; Rasko, D.A.; Ravel, J.; Brinkac, L.M.; Deboy, R.T.; Parker, C.T.; Daugherty, S.C.; et al. Major structural differences and novel potential virulence mechanisms from the genomes of multiple Campylobacter species. PLoS Biol. 2005, 3, e15. [Google Scholar] [CrossRef] [PubMed]
- Parkhill, J.; Wren, B.W.; Mungall, K.; Ketley, J.M.; Churcher, C.; Basham, D.; Chillingworth, T.; Davies, R.M.; Feltwell, T.; Holroyd, S.; et al. The genome sequence of the food-borne pathogen Campylobacter jejuni reveals hypervariable sequences. Nature 2000, 403, 665–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, J.; Britton, R.A.; Roos, S. Host-microbial symbiosis in the vertebrate gastrointestinal tract and the Lactobacillus reuteri paradigm. Proc. Natl. Acad. Sci. USA 2011, 108, 4645–4652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobierecka, P.A.; Wyszyńska, A.K.; Aleksandrzak-Piekarczyk, T.; Kuczkowski, M.; Tuzimek, A.; Piotrowska, W.; Górecki, A.; Adamska, I.; Wieliczko, A.; Bardowski, J.; et al. In vitro characteristics of Lactobacillus spp. strains isolated from the chicken digestive tract and their role in the inhibition of Campylobacter colonization. MicrobiologyOpen 2017, 6, e00512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neal-McKinney, J.M.; Lu, X.; Duong, T.; Larson, C.L.; Call, D.R.; Shah, D.H.; Konkel, M.E. Production of organic acids by probiotic Lactobacilli can be used to reduce pathogen load in poultry. PLoS ONE 2012, 7, e43928. [Google Scholar] [CrossRef]
- Doyle, M.P.; Roman, D.J. Growth and survival of Campylobacter fetus subsp. jejuni as a function of temperature and pH. J. Food Prot. 1981, 44, 596–601. [Google Scholar] [CrossRef]
- Reid, A.N.; Pandey, R.; Palyada, K.; Whitworth, L.; Doukhanine, E.; Stintzi, A. Identification of Campylobacter jejuni genes contributing to acid adaptation by transcriptional profiling and genome-wide mutagenesis. Appl. Environ. Microbiol. 2008, 74, 1598–1612. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Organism | Number of Strains | MIC a (µM) | MBC a (µM) |
|---|---|---|---|
| Campylobacter jejuni | 51 | 1.5–3.0 | 3.0–5.8 |
| Campylobacter coli | 20 | 1.5–3.0 | 1.5–5.8 |
| Campylobacter jejuni ATCC 33560 | 1 | 3.0 | 5.8 |
| Lactobacillus reuteri | 5 | 150.0 | 450.3 |
| Escherichia coli K12 | 1 | 16.5 | 50.1 |
| Strains | Treatment | pH 1 | Substrate Consumed | Metabolites Formed | |||||
|---|---|---|---|---|---|---|---|---|---|
| Glycerol 2 (mM) | Lactate (mM) | Acetate (mM) | Succinate (mM) | Acrolein (mM) | 1,3-PDO (mM) | 3-HP (mM) | |||
| C. jejuni N16-1419 | Glycerol | 7.2 ± 0.1 A | 0.0 | <0.1 A | 4.4 ± 0.5 A | 5.3 ± 0.4 A | ND | ND | ND |
| No glycerol | 7.3 ± 0.1 A | - | <0.1 A | 4.0 ± 0.3 A | 6.2 ± 0.6 A | ND | ND | ND | |
| L. reuteri PTA8-1 | Glycerol | 6.8 ± 0.1 A | 0.0 | 0.2 ± 0.1 A | 1.9 ± 0.1 A | ND | ND | ND | ND |
| No glycerol | 6.8 ± 0.1 A | - | 0.3 ± 0.1 A | 2.0 ± 0.1 A | ND | ND | ND | ND | |
| L. reuteri PTA5-F13 | Glycerol | 6.1 ± 0.1 A | 12.9 ± 1.1 | 0.7 ± 0.1 A | 3.0 ± 0.1 A | ND | 0.2 ± 0.0 | 5.3 ± 0.3 | 4.4 ± 0.3 |
| No glycerol | 6.8 ± 0.1 B | - | 0.2 ± 0.1 B | 1.9 ± 0.1 B | ND | ND | ND | ND | |
| C. jejuni N16-1419 + L. reuteri PTA8-1 | Glycerol | 7.2 ± 0.1 A | 0.0 | <0.1 A | 6.8 ± 0.1 A | 5.4 ± 0.6 A | ND | ND | ND |
| No glycerol | 7.3 ± 0.1 A | - | <0.1 A | 6.7 ± 0.4 A | 5.9 ± 0.8 A | ND | ND | ND | |
| C. jejuni N16-1419 + L. reuteri PTA5-F13 | Glycerol | 5.8 ± 0.1 A | 25.4 ± 0.4 | 2.2 ± 0.1A | 3.4 ± 0.1 A | 0.8 ± 0.1 A | 0.1 ± 0.0 | 6.2 ± 0.6 | 4.9 ± 0.8 |
| No glycerol | 7.3 ± 0.1 B | - | <0.1 B | 7.9 ± 0.5 B | 5.7 ± 1.3 B | ND | ND | ND | |
| Strains | Viable Cell Counts (Log CFU/mL) | 16S rRNA Gene Copy Number (Log Gene copies/mL) | ||
|---|---|---|---|---|
| Biphasic CAMH with 28 mM glycerol | Biphasic CAMH without glycerol | Biphasic CAMH with 28 mM glycerol | Biphasic CAMH without glycerol | |
| Mono-cultures | ||||
| C. jejuni N16-1419 | 8.1 ± 0.4 A | 8.4 ± 0.6 A | 9.6 ± 0.3 a | 9.8 ± 0.1 a |
| L. reuteri PTA8_1 | 7.0 ± 0.2 a | 6.6 ± 0.1 a | 7.9 ± 0.1 A | 7.3 ± 0.4 A |
| L. reuteri PTA5_F13 | 4.5 ± 0.4 A | 6.7 ± 0.1 B | 8.0 ± 0.2 a | 7.8 ± 0.2 a |
| Co-cultures * | ||||
| C. jejuni N16-1419 + L. reuteri PTA8_1 | 7.5 ± 0.4 A | 8.5 ± 0.8 A | 9.9 ± 0.2 a | 10.2 ± 0.1 a |
| C. jejuni N16-1419 + L. reuteri PTA5_F13 | BDL a | 9.3 ± 0.1 b | 6.3 ± 0.1 A | 10.2 ± 0.1 B |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asare, P.T.; Zurfluh, K.; Greppi, A.; Lynch, D.; Schwab, C.; Stephan, R.; Lacroix, C. Reuterin Demonstrates Potent Antimicrobial Activity Against a Broad Panel of Human and Poultry Meat Campylobacter spp. Isolates. Microorganisms 2020, 8, 78. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010078
Asare PT, Zurfluh K, Greppi A, Lynch D, Schwab C, Stephan R, Lacroix C. Reuterin Demonstrates Potent Antimicrobial Activity Against a Broad Panel of Human and Poultry Meat Campylobacter spp. Isolates. Microorganisms. 2020; 8(1):78. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010078
Chicago/Turabian StyleAsare, Paul Tetteh, Katrin Zurfluh, Anna Greppi, Denise Lynch, Clarissa Schwab, Roger Stephan, and Christophe Lacroix. 2020. "Reuterin Demonstrates Potent Antimicrobial Activity Against a Broad Panel of Human and Poultry Meat Campylobacter spp. Isolates" Microorganisms 8, no. 1: 78. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010078