Influence of Culture Conditions and Medium Compositions on the Production of Bacteriocin-Like Inhibitory Substances by Lactococcus lactis Gh1


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Microorganisms and Maintenance
2.3. Preparation of Inoculum and Culture Condition
2.4. Factors Influencing the Production of Bacteriocin-Like Inhibitory Substances (BLIS)
2.5. Selection of Culture Media for BLIS Production
2.6. Effects of Initial pH of Culture Media on BLIS Production
2.7. Effects of Inoculum Age, Size and Cultivation Conditions on BLIS Production
2.8. Screening of Medium Composition for BLIS Production
2.9. Analytical Procedures
2.10. Statistical Analysis
3. Results
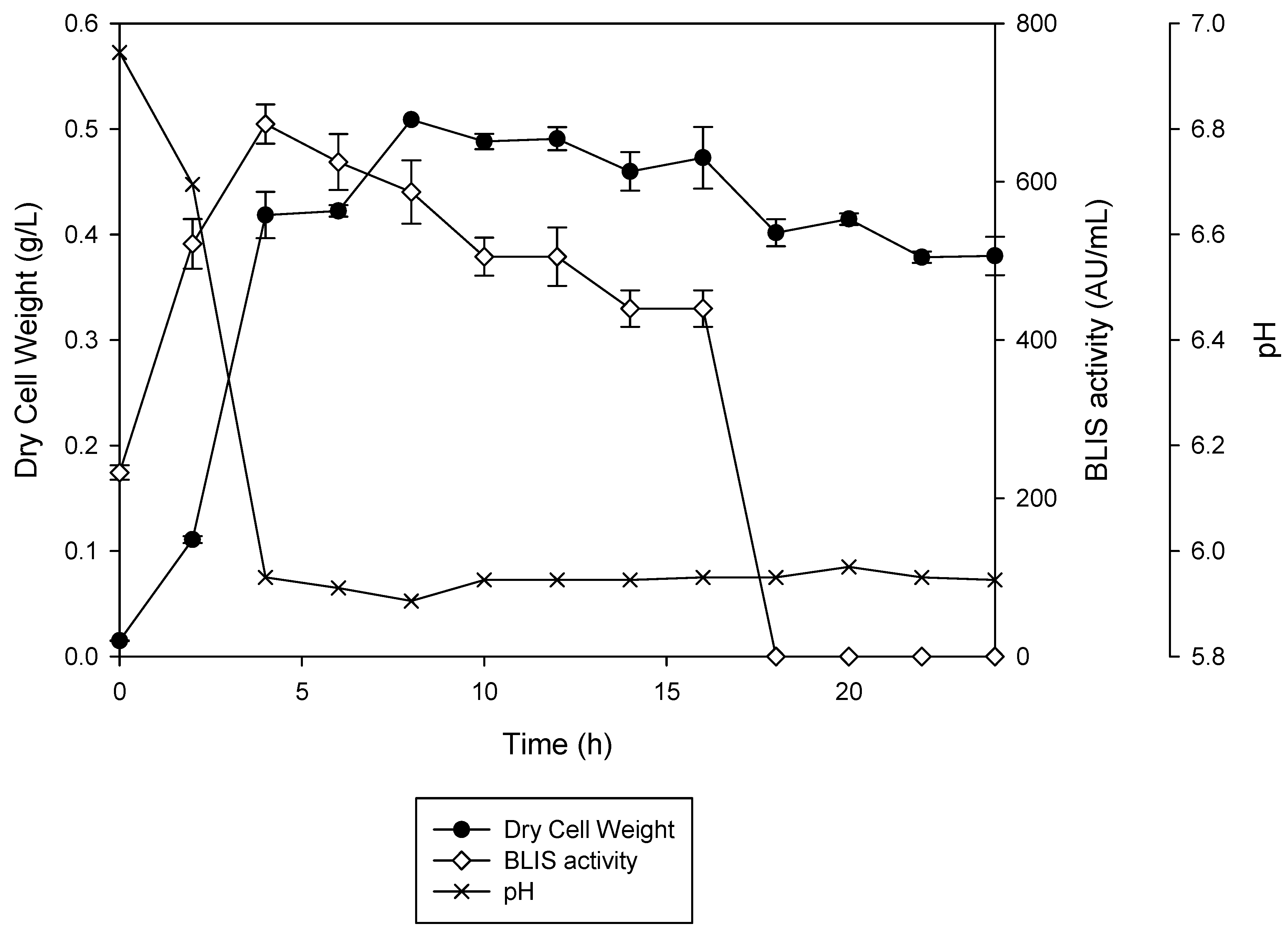
3.1. Effects of Culture Media on the Production of BLIS
3.2. Effects of Initial pH on BLIS Production
3.3. Effects of Inoculum Age, Size and Sub-Culture Frequency on BLIS Production
3.4. Effects of Different Organic and Inorganic Nitrogen Sources on BLIS Production
3.5. Effects of Different Types of Carbon Source on BLIS Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Singh, V.P. Recent approaches in food bio-preservation—A review. Open Vet. J. 2018, 8, 104–111. [Google Scholar] [CrossRef] [Green Version]
- Özogul, F.; Hamed, I. The importance of lactic acid bacteria for the prevention of bacterial growth and their biogenic amines formation: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1660–1670. [Google Scholar] [CrossRef]
- Abbasiliasi, S.; Ramanan, R.N.; Tengku Azmi, T.I.; Shuhaimi, M.; Mohammad, R.; Ariff, A.B. Partial characterization of antimicrobial compound produced by Lactobacillus paracasei LA 07, a strain isolated from Budu. Minerva Biotecnol. 2010, 22, 75–82. [Google Scholar]
- Silva, C.C.G.; Silva, S.P.M.; Ribeiro, S.C. Application of bacteriocins and protective cultures in dairy food preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef]
- Mills, S.; Serrano, L.; Griffin, C.; O’connor, P.M.; Schaad, G.; Bruining, C.; Hill, C.; Ross, R.P.; Meijer, W.C. Inhibitory activity of Lactobacillus plantarum LMG P-26358 against Listeria innocua when used as an adjunct starter in the manufacture of cheese. Microb. Cell Fact. 2011, 10, S7. [Google Scholar] [CrossRef] [Green Version]
- Grazia, S.E.; Sumayyah, S.; Haiti, F.S.; Sahlan, M.; Heng, N.C.K.; Malik, A. Bacteriocin-like inhibitory substance (BLIS) activity of Streptococcus macedonicus MBF10-2 and its synergistic action in combination with antibiotics. Asian Pac. J. Trop. Med. 2017, 10, 1140–1145. [Google Scholar] [CrossRef]
- Loh, J.Y.; Lim, Y.Y.; Ting, A.S.Y. Bacteriocin-like substances produced by Lactococcus lactis subsp. lactis CF4MRS isolated from fish intestine: Antimicrobial activities and inhibitory properties. Int. Food Res. J. 2017, 24, 394–400. [Google Scholar]
- Chakchouk-Mtibaa, A.; Smaoui, S.; Ktari, N.; Sellem, I.; Najah, S.; Karray-Rebai, I.; Mellouli, L. Biopreservative efficacy of bacteriocin BacFL31 in raw ground Turkey meat in terms of microbiological, physicochemical, and sensory qualities. Biocontrol Sci. 2017, 22, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Leelavatcharamas, V.; Arbsuwan, N.; Apiraksakorn, J.; Laopaiboon, P.; Kishida, M. Thermotolerant bacteriocin-producing lactic acid bacteria isolated from Thai local fermented foods and their bacteriocin productivity. Biocontrol Sci. 2011, 16, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Settanni, L.; Corsetti, A. Application of bacteriocins in vegetable food biopreservation. Int. J. Food Microbiol. 2008, 121, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.S.; Todorov, S.D.; Ivanova, I.; Chobert, J.-M.; Thomas Haertlé, T.; de Melo Franco, B.D.G. Improving safety of salami by application of bacteriocins produced by an autochthonous Lactobacillus curvatus isolate. Food Microbiol. 2015, 46, 254–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontana, C.; Cocconcelli, P.S.; Vignolo, G.; Saavedra, L. Occurrence of antilisterial structural bacteriocins genes in meat borne lactic acid bacteria. Food Control 2015, 47, 53–59. [Google Scholar] [CrossRef]
- Todorov, S.D.; Vaz-Velho, M.; de Melo Franco, B.D.G.; Holzapfel, W.H. Partial characterization of bacteriocins produced by three strains of Lactobacillus sakei, isolated from salpicao, a fermented meat product from North-West of Portugal. Food Control 2013, 30, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Szabóová, R.; Lauková, A.; Simonová, M.P.; Strompfová, V.; Chrastinova, L. Bacteriocin-producing enterococci’ from rabbit meat. Malays. J. Microbiol. 2012, 8, 211–218. [Google Scholar]
- Cintas, L.M.; Casaus, M.P.; Herranz, C.; Nes, I.F.; Hernández, P.E. Review: Bacteriocins of lactic acid bacteria. Food Sci. Technol. Int. 2001, 7, 281–305. [Google Scholar] [CrossRef]
- Abbasiliasi, S.; Ramanan, R.N.; Ibrahim, T.A.T.; Mustafa, S.; Mohamad, R.; Daud, H.M.; Ariff, A.B. Effect of medium composition and culture condition on the production of bacteriocin-like inhibitory substances (BLIS) by Lactobacillus Paracasei LA07, a strain isolated from Budu. Biotechnol. Biotechnol. Equip. 2011, 25, 2652–2657. [Google Scholar] [CrossRef] [Green Version]
- Abbasiliasi, S.; Tan, J.S.; Tengku Ibrahim, T.A.; Bashokouh, F.; Ramakrishnan, N.R.; Mustafa, S.; Ariff, A. Fermentation factors influencing the production of bacteriocins by lactic acid bacteria: A review. Rsc Adv. 2017, 7, 29395–29420. [Google Scholar] [CrossRef]
- Abbasiliasi, S.; Tan, J.S.; Kadkhodaei, S.; Nelofer, R.; Tengku Ibrahim, T.A.; Shuhaimi, M.; Ariff, A.B. Enhancement of BLIS production by Pediococcus acidilactici kp10 in optimized fermentation conditions using an artificial neural network. RSC Adv. 2016, 6, 6342–6349. [Google Scholar] [CrossRef]
- Castro, M.P.; Palavecino, N.Z.; Herman, C.; Garro, O.A.; Campos, C.A. Lactic acid bacteria isolated from artisanal dry sausages: Characterization of antibacterial compounds and study of the factors affecting bacteriocin production. Meat Sci. 2010, 87, 321–329. [Google Scholar] [CrossRef]
- Motta, A.S.; Brandelli, A. Influence of growth conditions on bacteriocin production by Brevibacterium linens. Appl. Microbiol. Biotechnol. 2003, 62, 163–167. [Google Scholar] [CrossRef]
- Dinarvand, M.; Rezaee, M.; Masomian, M.; Jazayeri, S.; Zareian, M.; Abbasi, S.; Ariff, A.B. Effect of C/N ratio and media optimization through response surface methodology on simultaneous productions of intra- and extracellular inulinase and invertase from Aspergillus niger ATCC 20611. Biomed. Res. Int. 2013, 2013, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.M.; Kim, J.S.; Kim, W.J. Optimization for the maximum bacteriocin production of Lactobacillus brevis DF01 using response surface methodology. Food Sci. Biotechnol. 2012, 21, 653–659. [Google Scholar] [CrossRef]
- Guerra, N.; Pastrana, L. Enhanced Nisin and Pediocin production on whey supplemented with different nitrogen sources. Biotechnol. Lett. 2001, 23, 609–612. [Google Scholar] [CrossRef]
- Arakawa, K.; Kawai, Y.; Fujitani, K.; Nishimura, J.; Kitizawa, H.; Komine, K.; Kai, K.; Saito, T. Bacteriocin production of probiotic Lactobacillus gasseri LA39 isolated from human feces in milk-based media. Anim. Sci. J. 2008, 79, 634–640. [Google Scholar] [CrossRef]
- Abbasiliasi, S.; Tan, J.S.; Ibrahim, T.A.T.; Ramanan, R.N.; Vakhshiteh, F.; Mustafa, S.; Ariff, A. Isolation of Pediococcus acidilactici Kp10 with ability to secrete bacteriocin-like inhibitory substance from milk products for applications in food industry. BMC Microbiol. 2012, 12, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jawan, R.; Abbasiliasi, S.; Mustafa, S.; Kapri, M.R.; Halim, M. In vitro evaluation of potential probiotic strain Lactococcus lactis Gh1 and its bacteriocin-like inhibitory substances for potential use in the food industry. Probiotics Antimicrob. Proteins 2020, 1–19. [Google Scholar] [CrossRef]
- Ünlü, G.; Nielsen, B.; Ionita, C. Production of antilisterial bacteriocins from lactic acid bacteria in dairy-based media: A comparative study. Probiotics Antimicrob. Proteins 2015, 7, 259–274. [Google Scholar] [CrossRef]
- Li, C.; Bai, J.; Cai, Z.; Ouyang, F. Optimization of a cultural medium for bacteriocin production by Lactococcus lactis using response surface methodology. J. Biotechnol. 2002, 93, 27–34. [Google Scholar] [CrossRef]
- Schirru, S.; Favaro, L.; Mangia, N.; Basaglia, M.; Casella, S.; Comunian, R.; Fancello, F.M.F.B.; de Souza Oliveira, R.; Todorov, S. Comparison of bacteriocins production from Enterococcus faecium strains in cheese whey and optimized commercial MRS medium. Ann. Microbiol. 2014, 64, 321–331. [Google Scholar] [CrossRef]
- Mahrous, H.; Mohamed, A.; El-Mongy, M.; El-Batal, A.; Hamza, H. Study of bacteriocin production and optimization using new isolates of Lactobacillus ssp. isolated from some dairy products under different culture conditions. Food Nutr. Sci. 2013, 4, 342–356. [Google Scholar]
- Liu, C.; Liu, Y.; Liao, W.; Wen, Z.; Chen, S. Application of statistically-based experimental designs for the optimization of Nisin production from whey. Biotechnol. Lett. 2003, 25, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Mataragas, M.; Metaxopoulos, J.; Galiotou, M.; Drosinos, E.H. Influence of pH and temperature by Leuconostoc mesenteroides L124 and Lactobacillus curvatus L442. Meat Sci. 2003, 64, 265–271. [Google Scholar] [CrossRef]
- Leroy, F.; Vankrunkelsven, S.; De Greef, J.; De Vuyst, L. The stimulating effect of a harsh environment on the bacteriocin activity by Enterococcus faecium RZS C5 and dependency on the environmental stress factor used. Int. J. Food Microbiol. 2003, 83, 27–38. [Google Scholar] [CrossRef]
- Taheri, P.; Samadi, N.; Ehsani, M.R.; Khoshayand, M.R.; Jamalifar, H. An evaluation and partial characterization of a bacteriocin produced by Lactococcus lactis subsp. lactis ST1 isolated from goat milk. Braz. J. Microbiol. 2012, 43, 1452–1462. [Google Scholar] [CrossRef]
- Lv, W.; Zhang, X.; Cong, W. Modelling the production of nisin by Lactococcus lactis in fed-batch culture. Appl. Microbiol. Biotechnol. 2006, 68, 322–326. [Google Scholar] [CrossRef]
- Cheigh, C.I.; Choi, H.J.; Park, H.; Kim, S.B.; Kook, M.C.; Kim, T.S.; Hwang, J.K.; Pyun, Y.R. Influence of growth conditions on the production of a nisin-like bacteriocin by Lactococcus lactis subsp. lactis A164 isolated from kimchi. J. Biotechnol. 2002, 95, 225–235. [Google Scholar] [CrossRef]
- Biswas, S.R.; Ray, P.; Johnson, M.C.; Ray, B. Influence of growth conditions on the production of a bacteriocin, pediocin AcH, by Pediococcus acidilactici H. Appl. Environ. Microbiol. 1991, 57, 1265–1267. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Johnson, M.C.; Ray, B. Novel method to extract large amounts of bacteriocins from lactic acid bacteria. Appl. Environ. Microbiol. 1992, 58, 3355–3359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbasiliasi, S.; Tan, J.S.; Ibrahim, T.A.T.; Ramanan, R.N.; Kadkhodaei, S.; Mustafa, S.; Ariff, A.B. Kinetic modeling of bacteriocin-like inhibitory substance secretion by Pediococcus acidilactici Kp10 and its stability in food manufacturing conditions. J. Food Sci. Technol. 2018, 55, 1270–1284. [Google Scholar] [CrossRef]
- Parente, E.; Brienza, C.; Ricciardi, A.; Addario, G. Growth and bacteriocin production by Enterococcus faecium DPC1146 in batch and continuous culture. J. Ind. Microbiol. Biotechnol. 1997, 18, 62–67. [Google Scholar] [CrossRef]
- Abriouel, H.E.; Valdivia, E.; Martinez-Bueno, M.; Maqueda, M.; Gallvez, A. A simple method for semi-preparative-scale production and recovery of enterocin AS-48 derived from Enterococcus faecalis subsp. liquefaciens A-48-32. J. Microbiol. Methods 2003, 55, 599–605. [Google Scholar] [CrossRef]
- Callewaert, R.; Vuyst, L.D. Bacteriocin production with Lactobacillus amylovorus DCE 471 is improved and stabilized by fed-batch fermentation. Appl. Environ. Microbiol. 2000, 66, 606–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coetzee, J.C.J. Increased Production of bacST4SA by Enterococcus Mundtii in an Industrial-Based Medium with pH-Control; University of Stellenbosch: Stellenbosch, South Africa, 2007. [Google Scholar]
- Guerra, N.P.; Rua, M.L.; Pastrana, L. Nutritional factors affecting the production of two bacteriocins from lactic acid bacteria on whey. Int. J. Food Microbiol. 2001, 70, 267–281. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Bacteriocin production by Enterococcus faecium RZS C5 is cell density limited and occurs in the very early growth phase. Int. J. Food Microbiol. 2002, 72, 155–164. [Google Scholar] [CrossRef]
- Kim, M.-H.; Kong, Y.-J.; Baek, H.; Hyun, H.-H. Optimization of culture conditions and medium composition for the production of micrococcin GO5 by Micrococcus sp. GO5. J. Biotechnol. 2006, 121, 54–61. [Google Scholar] [CrossRef]
- Motta, A.S.; Brandelli, A. Evaluation of environmental conditions for production of bacteriocin-like substance by Bacillus sp. strain P34. World J. Microbiol. Biotechnol. 2008, 24, 641–646. [Google Scholar] [CrossRef]
- Yang, E.; Fan, L.; Yan, J.; Jiang, Y.; Doucette, C.; Fillmore, S.; Walker, B. Influence of culture media, pH and temperature on growth and bacteriocin production of bacteriocinogenic lactic acid bacteria. AMB Express 2018, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, J.G.; Garro, M.S.; Savoy de Giori, G. Effect of pH on Lactobacillus fermentum growth, raffinose removal, α-galactosidase activity and fermentation products. Appl. Microbiol. Biotechnol. 2004, 65, 119–123. [Google Scholar] [CrossRef]
- Poolman, B.; Konings, W.N. Relation of growth of Streptococcus lactis and Streptococcus cremoris to amino acid transport. J. Bacteriol. 1988, 170, 700–707. [Google Scholar] [CrossRef] [Green Version]
- Bibal, B.; Goma, G.; Vayssier, Y.; Pareilleux, A. Influence of pH, lactose and lactic acid on the growth of Streptococcus cremoris: A kinetic study. Appl. Microbiol. Biotechnol. 1988, 23, 340–344. [Google Scholar] [CrossRef]
- Gonçalves, L.M.D.; Ramos, A.; Almeida, J.S.; Xavier, A.M.R.B.; Carrondo, M.J.T. Elucidation of the mechanism of lactic acid growth inhibition and production in batch cultures of Lactobacillus rhamnosus. Appl. Microbiol. Biotechnol. 1997, 48, 346–350. [Google Scholar] [CrossRef]
- Charlebois, D.A.; Balázsi, G. Modeling cell population dynamics. Silico Biol. 2019, 13, 21–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lajis, A.F. Biomanufacturing process for the production of bacteriocins from Bacillaceae family. Bioresour. Bioprocess. 2020, 7, 2020. [Google Scholar] [CrossRef]
- López-González, M.J.; Escobedo, S.; Rodríguez, A.; Neves, A.R.; Janzen, T.; Martínez, B. Adaptive evolution of industrial Lactococcus lactis under cell envelope stress provides phenotypic diversity. Front. Microbiol. 2018, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Morrissey, I.; Smith, J.T. The importance of oxygen in the killing of bacteria by ofloxacin and ciprofloxacin. Microbios 1994, 79, 43–53. [Google Scholar] [PubMed]
- Ramachandran, B.; Srivathsan, J.; Sivakami, V.; Harish, J.; Ravi Kumar, M.; Mukesh Kumar, D.J. Production and optimization of bacteriocin from Lactococcus lactis. J. Acad. Ind. Res. 2012, 1, 306–309. [Google Scholar]
- Lechiancole, T.; Ricciardi, A.; Parente, E. Optimization of media and fermentation conditions for the growth of Lactobacillus sakei. Ann. Microbiol. 2002, 52, 257–274. [Google Scholar]
- Dimov, S.; Peykov, S.; Raykova, D.; Ivanova, P. Influence of diverse sugars on BLIS production by three different Enterococcus strains. Trakia J. Sci. 2008, 6, 54–59. [Google Scholar]
- Delgado, A.; Arroyo López, F.N.; Brito, D.; Peres, C.; Fevereiro, P.; Garrido-Fernández, A. Optimum bacteriocin production by Lactobacillus plantarum 17.2b requires absence of NaCl and apparently follows a mixed metabolite kinetics. J. Biotechnol. 2007, 130, 193–201. [Google Scholar] [CrossRef]
- Todorov, S.D.; Dicks, L.M. Effect of growth medium on bacteriocin production by Lactobacillus plantarum ST194BZ, a strain isolated from boza. Food Technol. Biotechnol. 2005, 43, 165–173. [Google Scholar]
- Todorov, S.D.; Dicks, L.M. Effect of medium components on bacteriocin production by Lactobacillus plantarum strains ST23LD and ST341LD, isolated from spoiled olive brine. Microbiol. Res. 2006, 161, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Stolz, P.; Vogel, R.F.; Hammes, W.P. Utilization of electron acceptors by lactobacilli isolated from sourdough. II. Lactobacillus pontis, L. reuteri, L. amylovorus, and L. fermentum. Z. Lebensm. Unters. Forsch. 1995, 201, 402–410. [Google Scholar] [CrossRef]
- Messens, W.; Neysens, P.; Vansieleghem, W.; Vanderhoeven, J.; De Vuyst, L. Modeling growth and bacteriocin production by Lactobacillus amylovorus DCE 471 in response to temperature and pH values used for sourdough fermentations. Appl. Environ. Microbiol. 2002, 68, 1431–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vuyst, L.; Leroy, F. Bacteriocins from lactic acid bacteria: Production, purification, and food applications. J. Mol. Microbiol. Biotechnol. 2007, 13, 194–199. [Google Scholar] [CrossRef]
- De Vuyst, L.; Callewaert, R.; Crabbe, K. Primary metabolite kinetics of bacteriocin biosynthesis by Lactobacillus amylovorus and evidence for stimulation ofbacteriocin production under un- favourable growth conditions. Microbiology 1996, 142, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Matsusaki, H.; Endo, N.; Sonomoto, K.; Ishizaki, A. Lantibiotic nisin Z fermentative production by Lactococcus lactis IO-1: Relationship between production of the lantibiotic and lactate and cell growth. Appl. Microbiol. Biotechnol. 1996, 45, 36–40. [Google Scholar] [CrossRef]


{kind=link}
| Component | Amount (g/L) |
|---|---|
| Nutrient substrate (Extract of brain and heart, and peptones) | 27.5 |
| Sodium chloride | 5 |
| Di-Sodium hydrogen phosphate | 2.5 |
| D (+) glucose | 2 |
| Media | Time for PmX (h) | pH | Maximum BLIS Activity PmX (AU. mL−1) | Maximum Cell Concentration XmX (g·L−1) | Specific Growth Rate μmax (h−1) | |
|---|---|---|---|---|---|---|
| Initial | Final | |||||
| MRS | 16 | 5.43 ± 0.01 | 4.27 ± 0.04 | 375.48 ± 35.43 b | 0.49 ± 0.002 a | 0.09 ± 0.0057 b |
| M17 | 4 | 6.98 ± 0.01 | 6.49 ± 0.02 | 640.23 ± 21.06 a | 0.43 ± 0.020 b | 0.08 ± 0.0022 b |
| BHI | 4 | 6.95 ± 0.02 | 5.95 ± 0.00 | 672.86 ± 24.76 a | 0.46 ± 0.020 ab | 0.15 ± 0.0126 a |
| TSB | 4 | 6.92 ± 0.03 | 6.72 ± 0.02 | 637.84 ± 24.73 a | 0.48 ± 0.040 ab | 0.09 ± 0.0024 b |
| LB | - | 6.01 ± 0.01 | 3.66 ± 0.02 | 0.00 ± 0.00 c | 0.31 ± 0.002 c | 0.09 ± 0.0042 b |
| Initial pH | Time for PmX (h) | pH | Maximum BLIS Activity PmX (AU. mL−1) | Maximum Cell Concentration XmX (g·L−1) | Specific Growth Rate μmax (h−1) | |
|---|---|---|---|---|---|---|
| Initial | Final | |||||
| 2 | - | 2.10 ± 0.00 | 2.10 ± 0.00 | 0.00 ± 0.00 d | 0.00 ± 0.000 e | 0.000 ± 0.0000 c |
| 3 | - | 3.02 ± 0.00 | 3.02 ± 0.00 | 0.00 ± 0.00 d | 0.00 ± 0.000 e | 0.000 ± 0.0000 c |
| 4 | 8 | 4.13 ± 0.01 | 3.95 ± 0.01 | 464.42 ± 7.58 c | 0.09 ± 0.000 d | 0.016 ± 0.0000 c |
| 5 | 6 | 5.00 ± 0.04 | 4.48 ± 0.02 | 475.17 ± 5.45 c | 0.29 ± 0.002 b | 0.050 ± 0.0005 b |
| 6 | 6 | 6.37 ± 0.00 | 5.13 ± 0.02 | 548.67 ± 22.85 b | 0.30 ± 0.000 b | 0.051 ± 0.0003 b |
| 7.04 * | 6 | 7.10 ± 0.05 | 6.37 ± 0.00 | 599.45 ± 11.77 a | 0.36 ± 0.013 a | 0.104 ± 0.0025 a |
| 8 | 6 | 7.66 ± 0.01 | 7.10 ± 0.02 | 581.99 ± 26.38 ab | 0.27 ± 0.022 b | 0.046 ± 0.0166 b |
| 9 | 6 | 8.54 ± 0.02 | 7.89 ± 0.01 | 572.28 ± 26.64 ab | 0.22 ± 0.020 c | 0.043 ± 0.0142 b |
| Growth Phase | Time for PmX (h) | pH | Maximum BLIS Activity PmX (AU. mL−1) | Maximum Cell Concentration XmX (g·L−1) | Specific Growth Rate μmax (h−1) | |
|---|---|---|---|---|---|---|
| Initial | Final | |||||
| Mid-exponential | 6 | 6.91 ± 0.01 | 6.31 ± 0.00 | 665.96 ± 1.83 a | 0.28 ± 0.033 a | 0.113 ± 0.010 a |
| Late-exponential | 6 | 6.90 ± 0.01 | 6.28 ± 0.01 | 615.73 ± 5.34 b | 0.33 ± 0.017 a | 0.110 ± 0.004 a |
| Stationary | 8 | 6.90 ± 0.02 | 6.33 ± 0.01 | 627.98 ± 16.74 b | 0.31 ± 0.033 a | 0.100 ± 0.020 a |
| Inoculum Size % (v/v) | Time for PmX (h) | pH | Maximum BLIS Activity PmX (AU. mL−1) | Maximum Cell Concentration XmX (g L−1) | Specific Growth Rate μmax (h−1) | |
|---|---|---|---|---|---|---|
| Initial | Final | |||||
| 0.5 | 8 | 7.15 ± 0.01 | 6.33 ± 0.00 | 593.02 ± 57.36 b | 0.33 ± 0.004 d | 0.071 ± 0.001 d |
| 1.0 | 6 | 7.13 ± 0.01 | 6.33 ± 0.01 | 700.24 ± 55.44 a | 0.34 ± 0.002 c | 0.071 ± 0.001 d |
| 2.0 | 6 | 7.10 ± 0.00 | 6.29 ± 0.02 | 538.32 ± 72.05 bc | 0.42 ± 0.002 b | 0.095 ± 0.003 c |
| 4.0 | 4 | 7.06 ± 0.01 | 6.25 ± 0.00 | 532.83 ± 7.69 bc | 0.44 ± 0.005 a | 0.151 ± 0.001 a |
| 6.0 | 4 | 7.03 ± 0.04 | 6.22 ± 0.04 | 535.39 ± 17.98 bc | 0.44 ± 0.004 ab | 0.096 ± 0.002 c |
| 8.0 | 8 | 7.00 ± 0.02 | 6.21 ± 0.03 | 566.08 ± 18.36 b | 0.43 ± 0.002 ab | 0.092 ± 0.010 c |
| 10.0 | 10 | 6.95 ± 0.01 | 6.18 ± 0.02 | 479.60 ± 32.10 c | 0.42 ± 0.016 ab | 0.112 ± 0.003 b |
| Sub-Culturing Frequency | Time for PmX (h) | pH | Maximum BLIS Activity PmX (AU. mL−1) | Maximum Cell Concentration XmX (g·L−1) | Specific Growth Rate μmax (h−1) | |
|---|---|---|---|---|---|---|
| Initial | Final | |||||
| One time | 10 | 7.15 ± 0.01 | 6.55 ± 0.01 | 553.45 ± 4.58 | 0.27 ± 0.004 | 0.05 ± 0.001 |
| Two times | 6 | 7.12 ± 0.01 | 6.49 ± 0.01 | 728.83 ± 37.80 | 0.21 ± 0.002 | 0.05 ± 0.001 |
| Nitrogen Source | Time for PmX (h) | pH | Maximum BLIS Activity PmX (AU. mL−1) | Maximum Cell Concentration XmX (g·L−1) | Specific Growth Rate μmax (h−1) | |
|---|---|---|---|---|---|---|
| Initial | Final | |||||
| BHI | 6 | 6.94 ± 0.01 | 6.28 ± 0.00 | 567.05 ± 47.01 bc | 0.35 ± 0.004 d | 0.074 ± 0.0012 b |
| No nitrogen sources | - | 6.86 ± 0.00 | 6.29 ± 0.02 | 0.00 ± 0.00 e | 0.00 ± 0.00 i | 0.000 ± 0.0000 d |
| Organic Nitrogen | ||||||
| Yeast extract | 4 | 6.46 ± 0.01 | 4.98 ± 0.01 | 619.41 ± 26.02 b | 0.45 ± 0.005 b | 0.107 ± 0.0040 c |
| Meat extract | 4 | 6.51 ± 0.02 | 5.38 ± 0.01 | 594.87 ± 8.69 b | 0.42 ± 0.011 c | 0.1062 ± 0.0050 a |
| Peptone | 4 | 6.82 ± 0.03 | 6.58 ± 0.00 | 580.00 ± 14.14 b | 0.36 ± 0.016 d | 0.0547 ± 0.0025 bc |
| Soytone | 4 | 6.80 ± 0.00 | 4.78 ± 0.01 | 764.71 ± 15.15 a | 0.69 ± 0.026 a | 0.0542 ± 0.0274 c |
| Tryptone | 4 | 7.02 ± 0.01 | 6.71 ± 0.02 | 515.59 ± 33.57 cd | 0.20 ± 0.001 e | 0.0429 ± 0.0005 c |
| Inorganic Nitrogen | ||||||
| (NH4)2SO4 | 6 | 6.86 ± 0.02 | 5.40 ± 0.00 | 599.08 ± 35.88 b | 0.07 ± 0.0013 f | 0.0120 ± 0.0004 d |
| NH6PO4 | 6 | 5.08 ± 0.02 | 5.10 ± 0.01 | 509.07 ± 26.23 d | 0.01 ± 0.0007 hi | 0.0007 ± 0.0005 d |
| NH4NO3 | - | 6.82 ± 0.01 | 6.25 ± 0.01 | 0.00 ± 0.00 e | 0.03 ± 0.0004 gh | 0.0060 ± 0.0010 d |
| NH4Cl | - | 6.44 ± 0.00 | 5.98 ± 0.02 | 0.00 ± 0.00 e | 0.03 ± 0.0011 g | 0.0076 ± 0.0001 d |
| Carbon Source | Time for PmX (h) | pH | Maximum BLIS Activity PmX (AU. mL−1) | Maximum Cell Concentration XmX (g·L−1) | Specific Growth Rate μmax (h−1) | |
|---|---|---|---|---|---|---|
| Initial | Final | |||||
| BHI | 8 | 7.11 ± 0.00 | 6.19 ± 0.01 | 530.12 ± 3.38 d | 0.24 ± 0.0005 g | 0.061 ± 0.0003 f |
| No carbon sources | 10 | 6.89 ± 0.01 | 5.68 ± 0.00 | 634.82 ± 26.40 bc | 0.56 ± 0.0018 f | 0.046 ± 0.0003 g |
| Monosaccharides | ||||||
| Fructose | 10 | 6.86 ± 0.01 | 4.96 ± 0.00 | 715.36 ± 13.77 a | 0.66 ± 0.0018 c | 0.108 ± 0.0007 e |
| Glucose | 10 | 6.79 ± 0.00 | 4.79 ± 0.02 | 674.03 ± 25.05 abc | 0.67 ± 0.0110 b | 0.139 ± 0.0066 c |
| Galactose | 10 | 6.86 ± 0.02 | 5.15 ± 0.02 | 678.51 ± 1.23 abc | 0.65 ± 0.0018 d | 0.214 ± 0.0011 a |
| Disaccharides | ||||||
| Sucrose | 8 | 6.98 ± 0.00 | 4.85 ± 0.01 | 661.77 ± 17.54 abc | 0.70 ± 0.0018 a | 0.136 ± 0.0010 c |
| Lactose | 8 | 6.98 ± 0.00 | 5.53 ± 0.02 | 622.99 ± 28.62 c | 0.60 ± 0.0018 e | 0.123 ± 0.0018 d |
| Maltose | 8 | 6.99 ± 0.01 | 5.24 ± 0.02 | 687.45 ± 43.19 ab | 0.67 ± 0.0018 b | 0.198 ± 0.0010 b |
| Sugar Alcohol | ||||||
| Sorbitol | 8 | 7.01 ± 0.01 | 5.68 ± 0.00 | 636.48 ± 21.62 bc | 0.56 ± 0.0018 f | 0.206 ± 0.0027 ab |
| Mannitol | 8 | 7.00 ± 0.02 | 5.16 ± 0.02 | 651.78 ± 9.69 bc | 0.57 ± 0.0037 f | 0.199 ± 0.0011 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jawan, R.; Abbasiliasi, S.; Tan, J.S.; Mustafa, S.; Halim, M.; Ariff, A.B. Influence of Culture Conditions and Medium Compositions on the Production of Bacteriocin-Like Inhibitory Substances by Lactococcus lactis Gh1. Microorganisms 2020, 8, 1454. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101454
Jawan R, Abbasiliasi S, Tan JS, Mustafa S, Halim M, Ariff AB. Influence of Culture Conditions and Medium Compositions on the Production of Bacteriocin-Like Inhibitory Substances by Lactococcus lactis Gh1. Microorganisms. 2020; 8(10):1454. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101454
Chicago/Turabian StyleJawan, Roslina, Sahar Abbasiliasi, Joo Shun Tan, Shuhaimi Mustafa, Murni Halim, and Arbakariya B. Ariff. 2020. "Influence of Culture Conditions and Medium Compositions on the Production of Bacteriocin-Like Inhibitory Substances by Lactococcus lactis Gh1" Microorganisms 8, no. 10: 1454. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101454