Seasonal Variations in Soil Microbiota Profile of Termite (Syntermes wheeleri) Mounds in the Brazilian Tropical Savanna

 , , ,
, , , {kind=link}
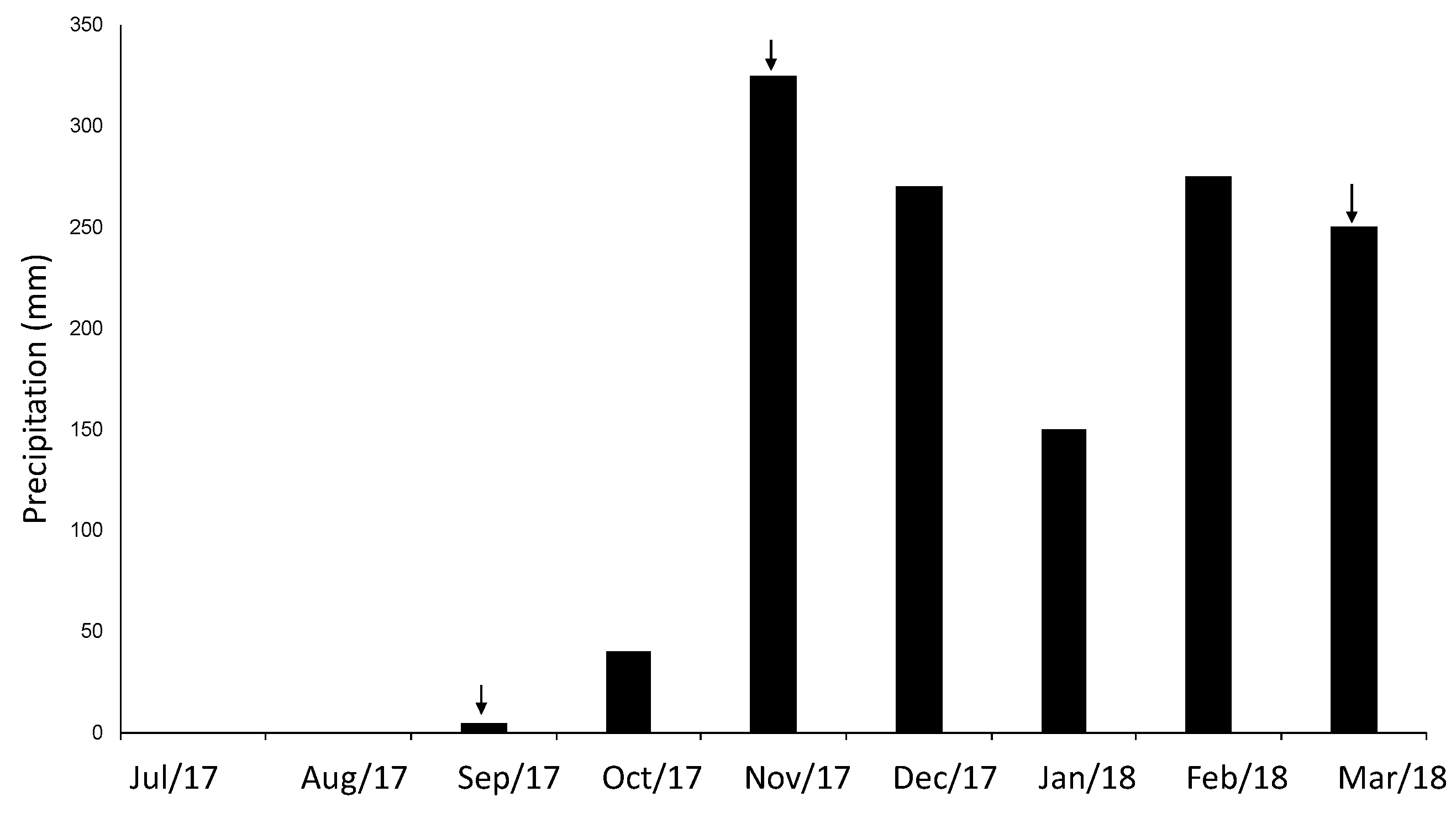{kind=link}
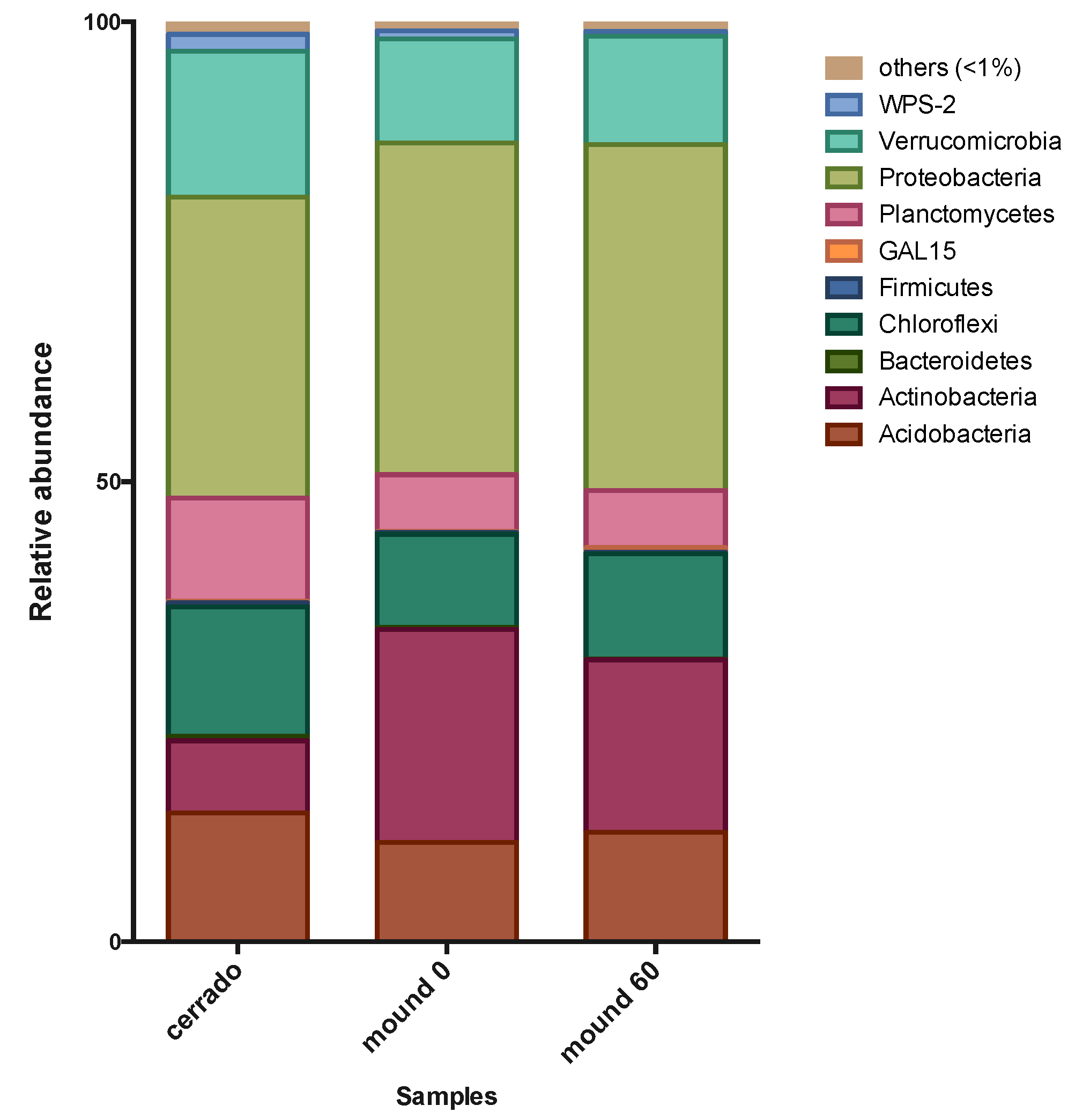{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sample Collection
2.2. Physico-Chemical Soil Parameters
2.3. Soil DNA Extraction and 16S rRNA Gene Library
3. Results
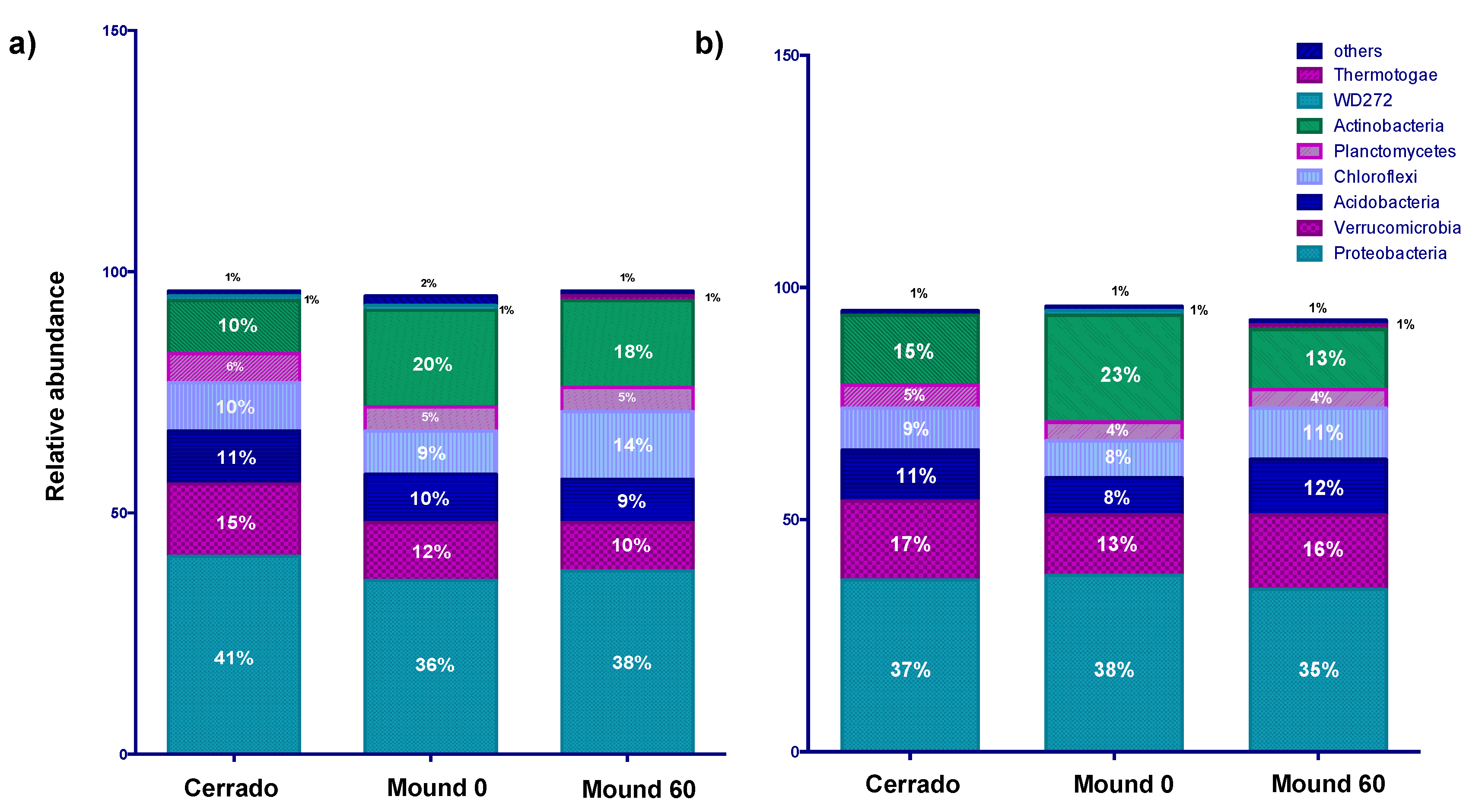
3.1. Comparison of Bacterial Community of Mound and Control Soil in the Dry Season
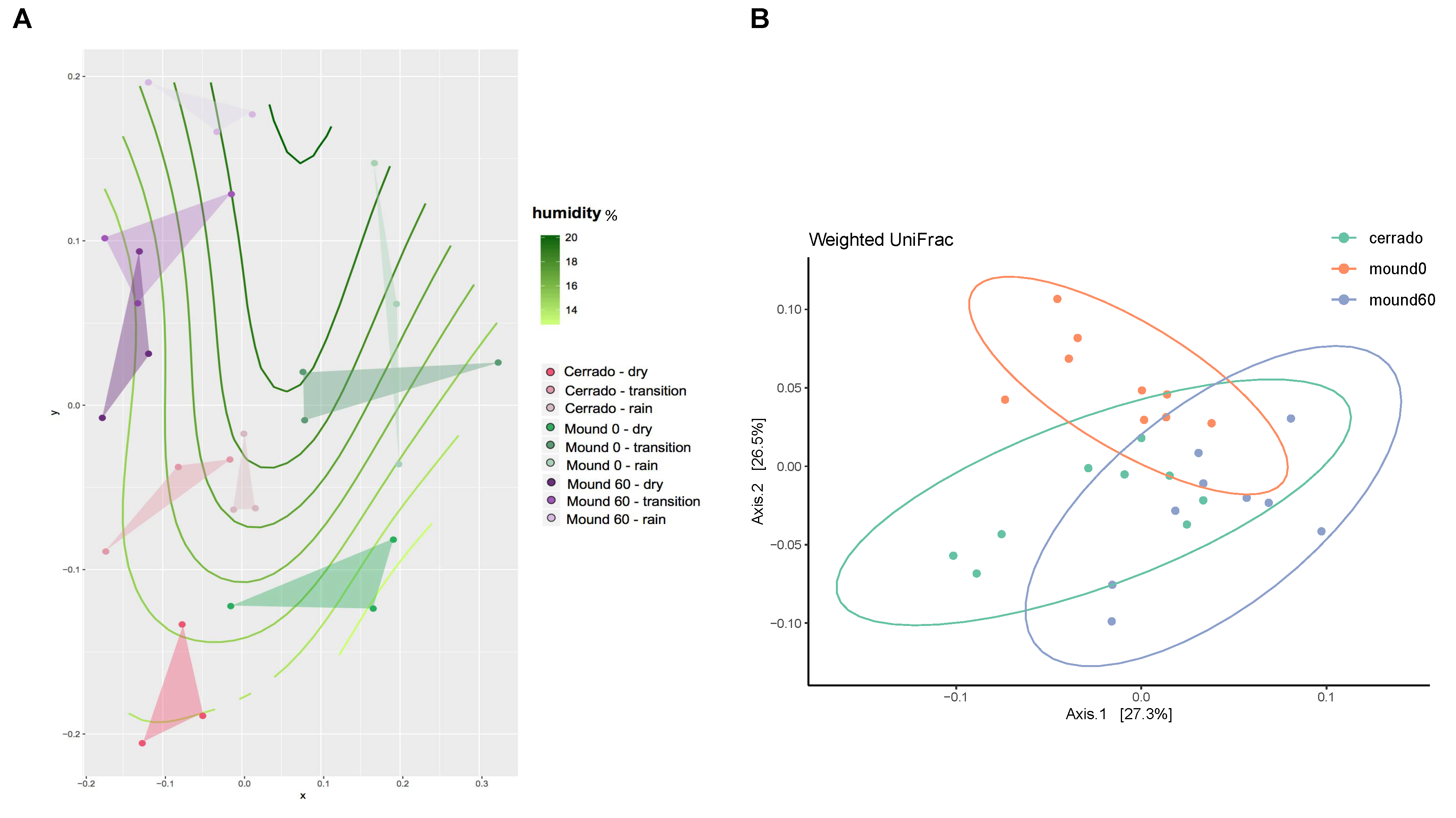
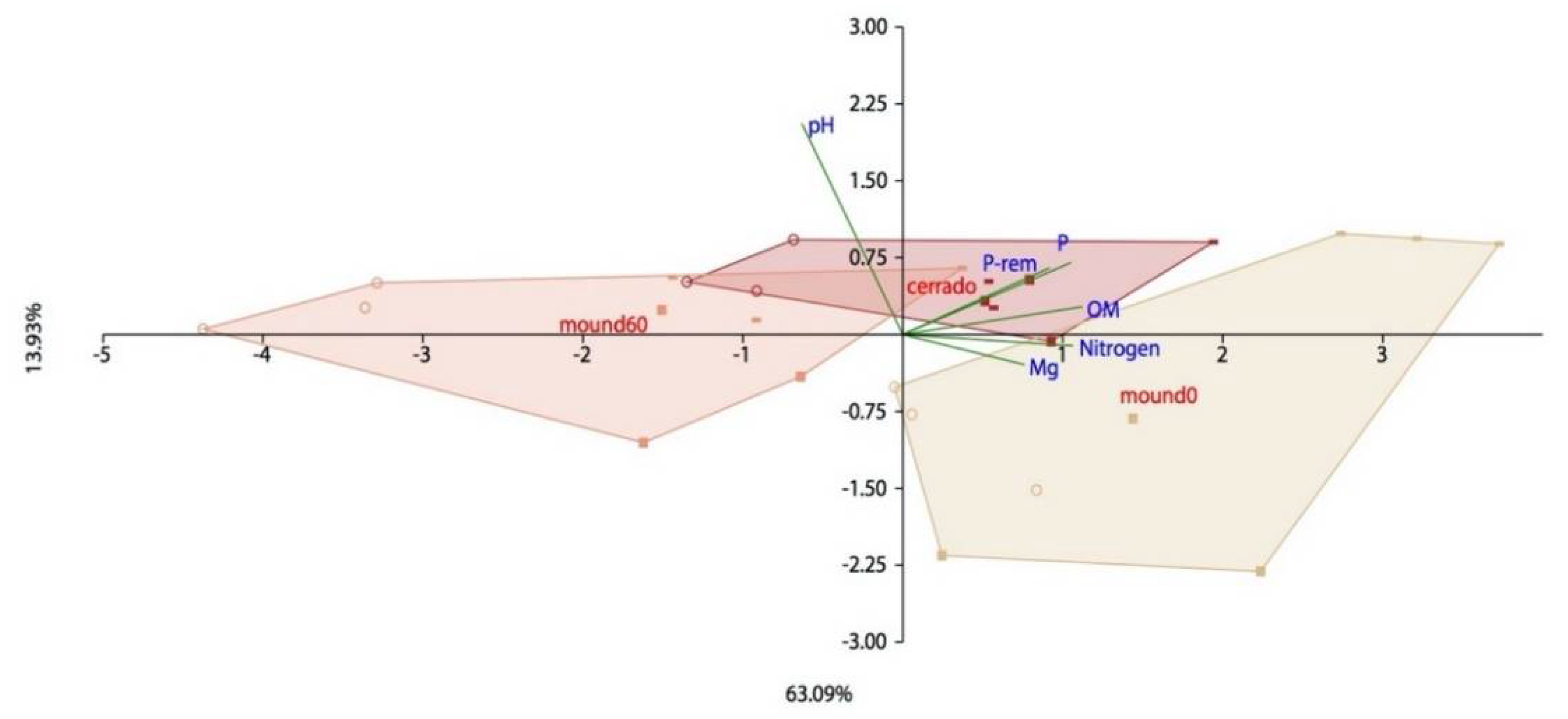
3.2. Seasonality and Chemical Parameters Effect on Bacterial Distribution in Mound and Control Soil Samples
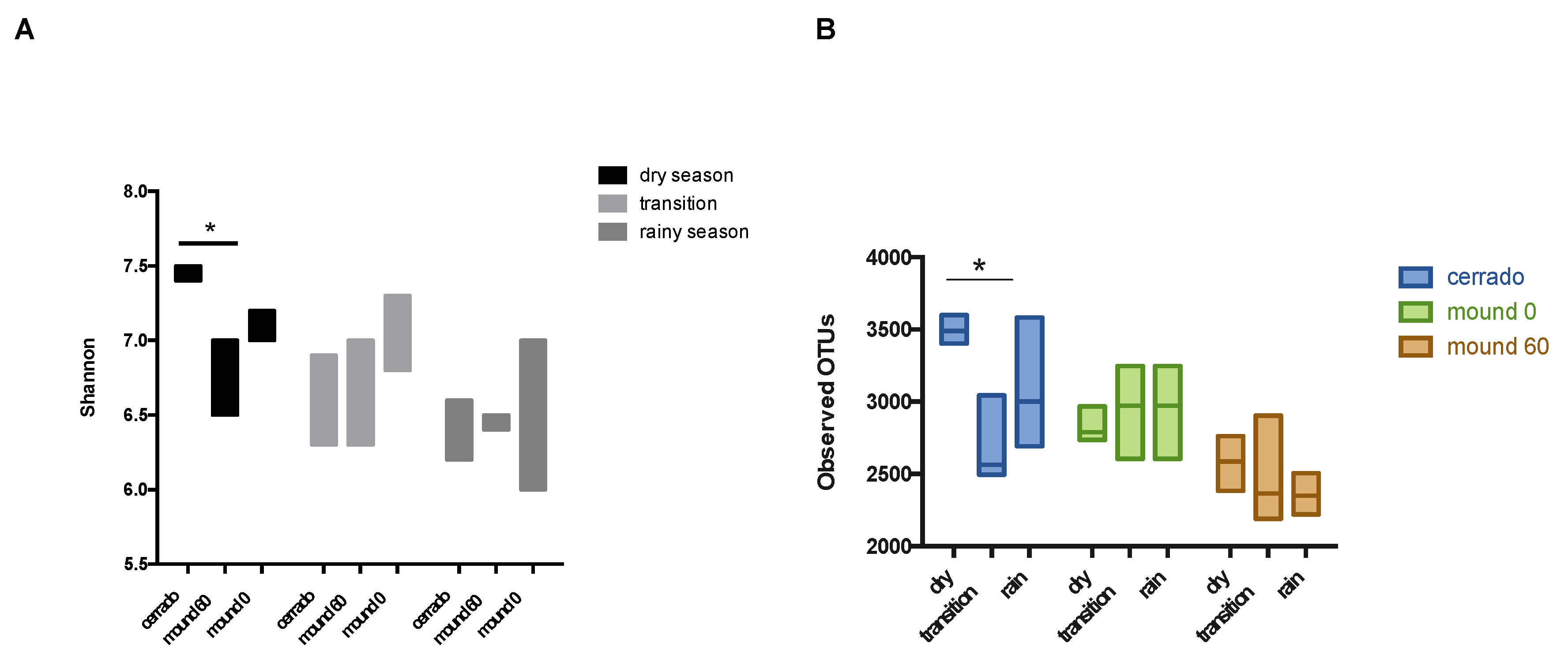
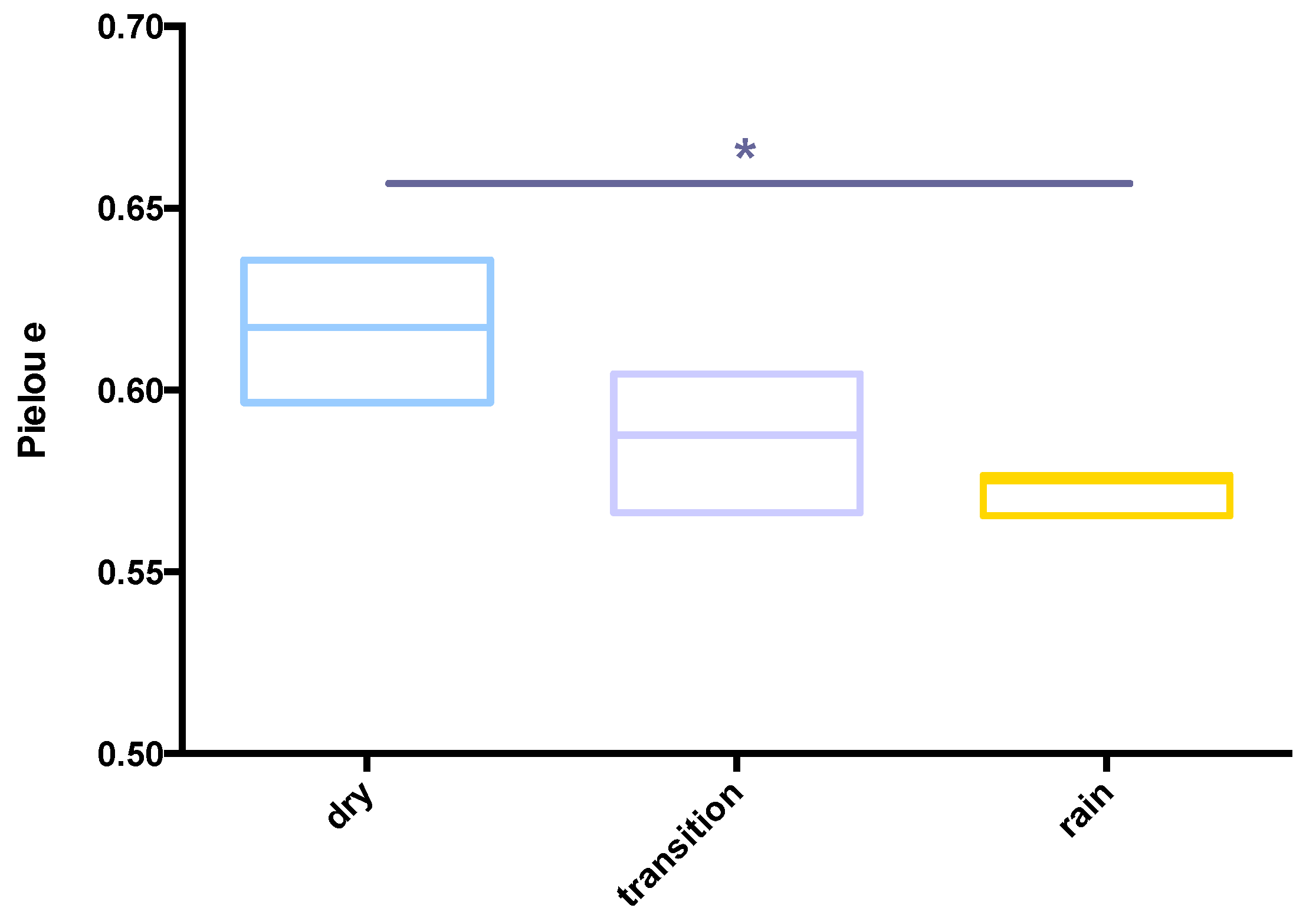
3.3. Ecological Parameters and Indexes for Soil Samples and Seasons
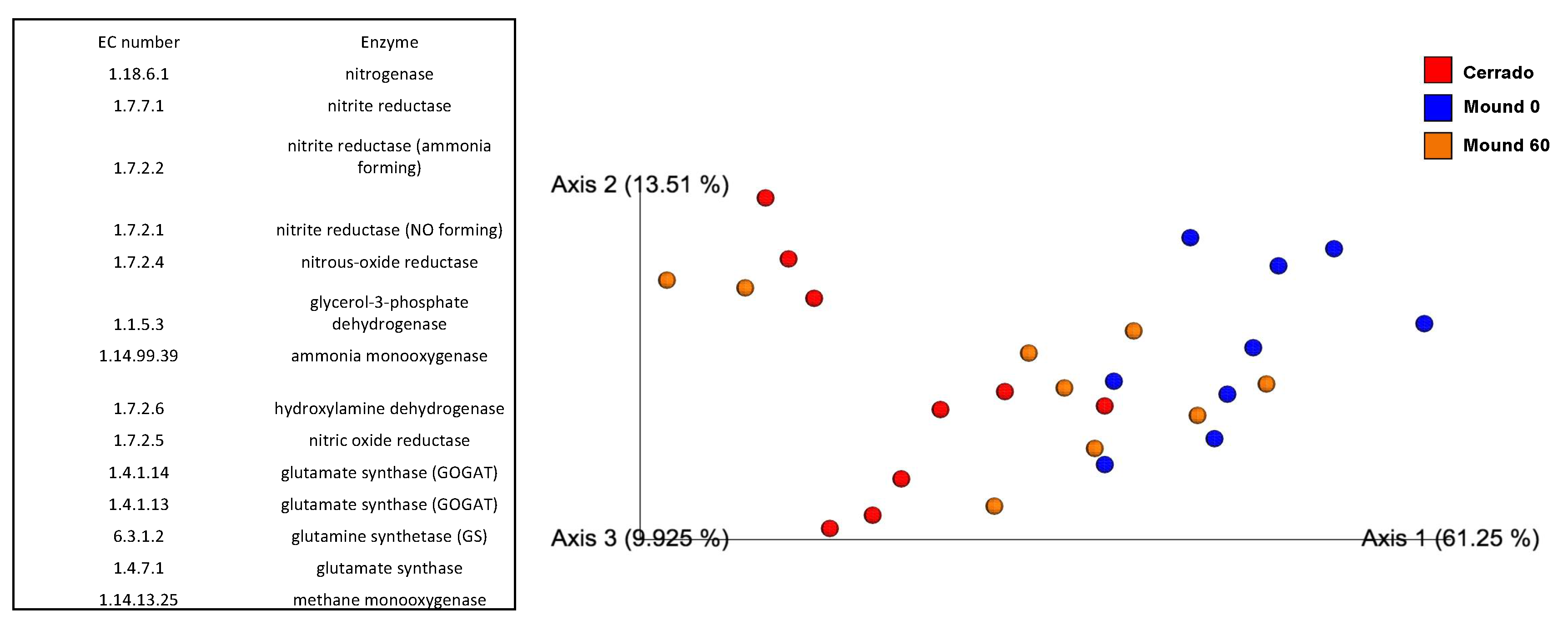
3.4. Functional Inference from Taxonomy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A





References
- Klink, C.A.; Machado, R.B. Conservation of the Brazilian Cerrado. Conserv. Biol. 2005, 19, 707–713. [Google Scholar] [CrossRef]
- Marques, J.J.G.; Schulze, D.G.; Curi, N.; Mertzman, S.A. Trace element geochemistry in Brazilian Cerrado soils. Geoderma 2004, 121, 31–43. [Google Scholar] [CrossRef]
- Furely, P.A.; Ratter, J.A. Soil resources and plant communities of the central Brazilian Cerrado and their development. J. Biogeogr. 2009, 15, 97–108. [Google Scholar] [CrossRef]
- Scalon, M.C.; Haridasan, M.; Franco, A. A comparative study of aluminium and nutrient concentrations in mistletoes on aluminium-accumulating and non-accumulating hosts. Plant Biol. 2013, 15, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Testoni, S.A.; De Almeida, J.A.; Da Silva, L.; Andrade, G.R.P. Clay mineralogy of Brazilian Oxisols with shrinkage properties. Revista Brasileira de Ciência do Solo 2017, 41, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Mendes, I.D.C.; Fernandes, M.F.; Chaer, G.M.; Junior, F.B.D.R. Biological functioning of Brazilian Cerrado soils under different vegetation types. Plant Soil 2012, 359, 183–195. [Google Scholar] [CrossRef]
- da Silva, M.R.S.S. Produção de serapilheira, biomassa e diversidade de comunidades bacterianas do solo em áreas de Cerrado sob diferentes usos e manejos; University of Brasilia: Brasilia, Brazil, 2004. [Google Scholar]
- Constantino, R.; Acioli, A.N.S. Termite diversity in Brazil (Insecta: Isoptera). In Soil Biodiversity in Amazonian and Other Brazilian Ecosystems; CABI Publishing: Wallingford, UK, 2006; pp. 117–128. [Google Scholar]
- Lee, K.E.; Wood, T.G. Termites and Soils; Academic Press: London, UK, 1971; p. 251. [Google Scholar] [CrossRef]
- Singh, K.; Muljadi, B.P.; Raeini, A.Q.; Jost, C.; Vandeginste, V.; Blunt, M.J.; Theraulaz, G.; Degond, P. The architectural design of smart ventilation and drainage systems in termite nests. Sci. Adv. 2019, 5, eaat8520. [Google Scholar] [CrossRef] [Green Version]
- Fageria, N.K.; Baligar, V.C. Properties of termite mound soils and responses of rice and bean to nitrogen, phosphorus, and potassium fertilization on such soil. Commun. Soil Sci. Plant Anal. 2005, 35, 2097–2109. [Google Scholar] [CrossRef]
- Araujo, J.F.; De Castro, A.P.; Costa, M.M.C.; Togawa, R.C.; Júnior, G.J.P.; Quirino, B.F.; Bustamante, M.M.D.C.; Williamson, L.; Handelsman, J.; Kruger, R.H. Characterization of soil bacterial assemblies in Brazilian savanna-like vegetation reveals acidobacteria dominance. Microb. Ecol. 2012, 64, 760–770. [Google Scholar] [CrossRef]
- Lavelle, P.; Spain, A. Soil Ecology Part II, Chapter IV: Functioning of the Soil System; Springer: Dordrecht, The Netherlands, 2001. [Google Scholar]
- Turner, J.S. Beyond biomimicry: What termites can tell us about realizing the living building. In Proceedings of the 1st International Conference on Industrialized, Integrated and Intelligent Construction, Loughborough, UK, 14–16 May 2008. [Google Scholar]
- Nannipieri, P.; Ascher-Jenull, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Dar, G.H. Soil Microbiology, Ecology and Biochemistry, 3rd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Bignell, D.E.; Roisin, Y.; Lo, N. Biology of Termites: A Modern Synthesis; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Moreira, E.A.; Alvarez, T.M.; Persinoti, G.F.; Paixão, D.A.A.; Menezes, L.R.; Cairo, J.P.L.F.; Squina, F.M.; Costa-Leonardo, A.M.; Carrijo, T.F.; Arab, A. Microbial Communities of the gut and nest of the humus- and litter-feeding termite Procornitermes araujoi (Syntermitinae). Curr. Microbiol. 2018, 75, 1609–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantino, R. Padrões de diversidade e endemismo de témitas no bioma Cerrado. In Biodiversidade, Ecologia e Conservação do Cerrado; Ministério do Meio Ambiente: Brasilia, Brazil, 2005; pp. 319–333. [Google Scholar]
- Santana, R.H.; Catão, E.C.P.; Lopes, F.A.C.; Constantino, R.; Barreto, C.; Kruger, R.H. The gut microbiota of workers of the litter-feeding termite Syntermes wheeleri (Termitidae: Syntermitinae): Archaeal, bacterial, and fungal communities. Microb. Ecol. 2015, 70, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Catani, R.A.; Bittencourt, V.C.; de Cangiani, A.M. Extração e determinação do íon amônio do solo. An da Esc Super Agric Luiz Queiroz 1969, 26, 57–73. [Google Scholar] [CrossRef] [Green Version]
- Claessen, M.E.C.; De Barreto, W.O. MANUAL de MÉTODOS de ANÁLISE de SOLO; EMBRAPA: Brasilia, Brazil, 1996. [Google Scholar]
- Griffiths, R.I.; Whiteley, A.S.; O’Donnell, A.G.; Bailey, M.J. Rapid method for coextraction of DNA and RNA from natural environments for analysis of ribosomal DNA- and rRNA-based microbial community composition. Appl. Environ. Microbiol. 2000, 66, 5488–5491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. FASTQC A Quality Control Tool for High Throughput Sequence Data; Babraham Bioinforma: Cambridge, UK, 2010; Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 25 September 2020).
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. Peer J. 2016, 4, e2584. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O′Hara, R.B.; Simpson, G.L.; Solymos, P.; Henry, M.; et al. Vegan Community Ecology Package. R. Package Vegan Vers. 2.2-1; World Agrofestry Centre: Nairobi, Kenya, 2015. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Shiny-phyloseq: Web application for interactive microbiome analysis with provenance tracking. Bioinform. 2014, 31, 282–283. [Google Scholar] [CrossRef]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Caspi, R.; Billington, R.; Fulcher, C.A.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Latendresse, M.; Midford, P.E.; Ong, Q.; Ong, W.K.; et al. The MetaCyc database of metabolic pathways and enzymes. Nucleic Acids Res. 2018, 46, D633–D639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Araujo, A.S.F.; Mendes, L.W.; Lemos, L.N.; Antunes, J.E.L.; Beserra, J.E.A., Jr.; de Lyra, M.D.C.C.P.; Figueiredo, M.D.V.B.; Lopes, A.C.D.A.; Gomes, R.L.F.; Bezerra, W.M.; et al. Protist species richness and soil microbiome complexity increase towards climax vegetation in the Brazilian Cerrado. Commun. Biol. 2018, 1, 135. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.; Fierer, N. A global atlas of the dominant bacteria found in soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappé, M.S.; Giovannoni, S.J. The uncultured microbial majority. Annu. Rev. Microbiol. 2003, 57, 369–394. [Google Scholar] [CrossRef] [Green Version]
- Sujada, N.; Sungthong, R.; Lumyong, S. Termite Nests as an abundant source of cultivable actinobacteria for biotechnological purposes. Microbes Environ. 2014, 29, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Enagbonma, B.J.; Aremu, B.R.; Babalola, O.O. Profiling the functional diversity of termite mound soil bacteria as revealed by shotgun sequencing. Genes 2019, 10, 637. [Google Scholar] [CrossRef] [Green Version]
- Herve, V.; Liu, P.; Dietrich, C.; Sillam-Dussès, D.; Stiblik, P.; Šobotník, J.; Brune, A. Phylogenomic analysis of 589 metagenome-assembled genomes encompassing all major prokaryotic lineages from the gut of higher termites. Peer J. 2020, 8, e8614. [Google Scholar] [CrossRef] [Green Version]
- Hongoh, Y. Toward the functional analysis of uncultivable, symbiotic microorganisms in the termite gut. Cell. Mol. Life Sci. 2011, 68, 1311–1325. [Google Scholar] [CrossRef]
- Warnecke, F.; Luginbühl, P.; Ivanova, N.N.; Ghassemian, M.; Richardson, T.H.; Stege, J.T.; Cayouette, M.; McHardy, A.C.; Djordjevic, G.; Aboushadi, N.; et al. Metagenomic and functional analysis of hindgut microbiota of a wood-feeding higher termite. Nature 2007, 450, 560–565. [Google Scholar] [CrossRef]
- Manjula, A.; Pushpanathan, M.; Sathyavathi, S.; Gunasekaran, P.; Rajendhran, J. Comparative Analysis of Microbial Diversity in Termite Gut and Termite Nest Using Ion Sequencing. Curr. Microbiol. 2015, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Minkley, N.; Fujita, A.; Brune, A.; Kirchner, W.H. Nest specificity of the bacterial community in termite guts (Hodotermes mossambicus). Insectes Sociaux 2006, 53, 339–344. [Google Scholar] [CrossRef]
- Kirchner, W.H.; Minkley, N. Nestmate discrimination in the harvester termite Hodotermes mossambicus. Insectes Sociaux 2003, 50, 222–225. [Google Scholar] [CrossRef]
- Robertson, L.A.; Kuenen, J.G. The Prokaryotes; Springer: New York, NY, USA, 2006; Volume 5. [Google Scholar]
- Brown, C.M.; Dilworth, M.J. Ammonia assimilation by rhizobium cultures and bacteroids. J. Gen. Microbiol. 1975, 86, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, I.R. Primary products of symbiotic nitrogen fixation. II. Pulse-labelling of serradella nodules with 15N2. Biochem. Biophys. Acta Gen. Subj. 1966, 130, 295–303. [Google Scholar] [CrossRef]
- WoolFolk, C.; Shapiro, B.; Stadtman, E. Regulation of glutamine synthetase. Arch. Biochem. Biophys. 1966, 116, 177–192. [Google Scholar] [CrossRef]
- Fildes, P.; Gladstone, G.P. Glutamine and the Growth of Bacteria. Br. J. Exp. Pathol. 1939, 20, 334–341. [Google Scholar]
- Vanoni, M.A.; Curti, B. Glutamate synthase: A complex iron-sulfur flavoprotein. Cell. Mol. Life Sci. 1999, 55, 617–638. [Google Scholar] [CrossRef]
- Feehily, C.; Karatzas, K.-A. Role of glutamate metabolism in bacterial responses towards acid and other stresses. J. Appl. Microbiol. 2012, 114, 11–24. [Google Scholar] [CrossRef]
- Baveye, P.C.; Otten, W.; Kravchenko, A.; Balseiro-Romero, M.; Beckers, É.; Chalhoub, M.; Darnault, C.; Eickhorst, T.; Garnier, P.; Hapca, S.; et al. Emergent properties of microbial activity in heterogeneous soil microenvironments: Different research approaches are slowly converging, yet major challenges remain. Front. Microbiol. 2018, 9, 1–48. [Google Scholar] [CrossRef]
- Zhalnina, K.; Dias, R.; De Quadros, P.D.; Davis-Richardson, A.; Camargo, F.A.O.; Clark, I.M.; McGrath, S.P.; Hirsch, P.; Triplett, E.W. Soil pH determines microbial diversity and composition in the park grass experiment. Microb. Ecol. 2014, 69, 395–406. [Google Scholar] [CrossRef]
- Reeves, J.L.; Liebig, M. Depth matters: Soil pH and dilution effects in the Northern Great Plains. Soil Sci. Soc. Am. J. 2016, 80, 1424–1427. [Google Scholar] [CrossRef]
- Da Silva, M.R.S.S.; De Castro, A.P.; Kruger, R.H.; Bustamante, M.M.D.C. Soil bacterial communities in the Brazilian Cerrado: Response to vegetation type and management. Acta Oecologica 2019, 100, 103463. [Google Scholar] [CrossRef]
- Wittebolle, L.; Marzorati, M.; Clement, L.; Balloi, A.; Daffonchio, D.; Heylen, K.; De Vos, P.; Verstraete, W.; Boon, N. Initial community evenness favours functionality under selective stress. Nature 2009, 458, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Bell, T.; Newman, J.A.; Silverman, B.W.; Turner, S.L.; Lilley, A.K. The contribution of species richness and composition to bacterial services. Nature 2005, 436, 1157–1160. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.M.; Marques, J.P.; Pinto-Ribeiro, I.; Costa, J.–L.; Carneiro, F.; Machado, J.C.; Figueiredo, C. Gastric microbial community profiling reveals a dysbiotic cancer-associated microbiota. Gut 2017, 67, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Yuan, T.; Chen, L.; Xie, Z.; Shen, Z. Occurrence of potentially pathogenic bacteria on shared bicycles. Int. J. Hyg. Environ. Heal. 2020, 224, 113442. [Google Scholar] [CrossRef]
- Boon, E.; Meehan, C.J.; Whidden, C.; Wong, D.H.-J.; Langille, M.G.I.; Beiko, R.G. Interactions in the microbiome: Communities of organisms and communities of genes. FEMS Microbiol. Rev. 2014, 38, 90–118. [Google Scholar] [CrossRef] [Green Version]
- Bill, M.; Chidamba, L.; Gokul, J.K.; Labuschagne, N.; Korsten, L. Bacterial community dynamics and functional profiling of soils from conventional and organic cropping systems. Appl. Soil Ecol. 2020, 157, 103734. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The Microbial Engines That Drive Earth’s Biogeochemical Cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guimaraes, H.I.P.; Santana, R.H.; Silveira, R.; Pinto, O.H.B.; Quirino, B.F.; Barreto, C.C.; Bustamante, M.M.d.C.; Krüger, R.H. Seasonal Variations in Soil Microbiota Profile of Termite (Syntermes wheeleri) Mounds in the Brazilian Tropical Savanna. Microorganisms 2020, 8, 1482. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101482
Guimaraes HIP, Santana RH, Silveira R, Pinto OHB, Quirino BF, Barreto CC, Bustamante MMdC, Krüger RH. Seasonal Variations in Soil Microbiota Profile of Termite (Syntermes wheeleri) Mounds in the Brazilian Tropical Savanna. Microorganisms. 2020; 8(10):1482. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101482
Chicago/Turabian StyleGuimaraes, Helena Ipe Pinheiro, Renata Henrique Santana, Rafaella Silveira, Otavio Henrique Bezerra Pinto, Betania Ferraz Quirino, Cristine Chaves Barreto, Mercedes Maria da Cunha Bustamante, and Ricardo Henrique Krüger. 2020. "Seasonal Variations in Soil Microbiota Profile of Termite (Syntermes wheeleri) Mounds in the Brazilian Tropical Savanna" Microorganisms 8, no. 10: 1482. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101482