Unravelling the Features of Success of VIM-Producing ST111 and ST235 Pseudomonas aeruginosa in a Greek Hospital

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Isolates, Identification and Susceptibility Testing
2.2. Confirmation of Carbapenemase Production
2.3. Multilocus Sequence Typing
2.4. Integron Analysis
2.5. Plasmid Analysis
2.6. Whole Genome Sequencing
2.7. Nucleotide Sequence Accession Numbers
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nicolle, L.E. Complicated urinary tract infection in adults. Can. J. Infect. Dis. Med. 2005, 16, 349–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. 2017. Available online: http://www.who.int/medicines/publications/global-prioritylist-antibiotic-resistant-bacteria/en/ (accessed on 27 February 2020).
- Livermore, D.M. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: Our worst nightmare? Clin. Infect. Dis. 2002, 34, 634–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzouvelekis, L.S.; Markogiannakis, A.; Psichogiou, M.; Tassios, P.T.; Daikos, G.L. Carbapenemases in Klebsiella pneumoniae and other Enterobacteriaceae: An evolving crisis of global dimensions. Clin. Microbiol. Rev. 2012, 25, 682–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsakris, A.; Pournaras, S.; Woodford, N.; Palepou, M.F.; Babini, G.S.; Douboyas, J.; Livermore, D.M. Outbreak of infections caused by Pseudomonas aeruginosa producing VIM-1 carbapenemase in Greece. J. Clin. Microbiol. 2000, 38, 1290–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giakkoupi, P.; Petrikkos, G.; Tzouvelekis, L.S.; Tsonas, S.; Legakis, N.J.; Vatopoulos, A.C.; WHONET Greece Study Group. Spread of integron-associated VIM-type metallo-beta-lactamase genes among imipenem-nonsusceptible Pseudomonas aeruginosa strains in Greek hospitals. J. Clin. Microbiol. 2003, 41, 822–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pournaras, S.; Tsakris, A.; Maniati, M.; Tzouvelekis, L.S.; Maniatis, A.N. Novel variant (bla(VIM-4)) of the metallo-beta-lactamase gene bla(VIM-1) in a clinical strain of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2002, 46, 4026–4028. [Google Scholar] [CrossRef] [Green Version]
- Pournaras, S.; Maniati, M.; Petinaki, E.; Tzouvelekis, L.S.; Tsakris, A.; Legakis, N.J.; Maniatis, A.N. Hospital outbreak of multiple clones of Pseudomonas aeruginosa carrying the unrelated metallo-beta-lactamase gene variants blaVIM-2 and blaVIM-4. J. Antimicrob. Chemother. 2003, 51, 1409–1414. [Google Scholar] [CrossRef] [Green Version]
- Liakopoulos, A.; Mavroidi, A.; Katsifas, E.A.; Theodosiou, A.; Karagouni, A.D.; Miriagou, V.; Petinaki, E. Carbapenemase-producing Pseudomonas aeruginosa from Central Greece: Molecular epidemiology and genetic analysis of class I integrons. BMC Infect. Dis. 2013, 13, 505. [Google Scholar] [CrossRef] [Green Version]
- EUCAST. European Committee on Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID). Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth dilution. Clin. Microbiol. Infect. 2003, 9, ix–xv. [Google Scholar] [CrossRef] [Green Version]
- Wayne, P.A. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; CLSI 2013, 23rd International Supplement, CLSI document M100-S23; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2013. [Google Scholar]
- Lee, K.; Lim, Y.S.; Yong, D.; Yum, J.H.; Chong, Y. Evaluation of the Hodge test and the imipenem-EDTA double-disk synergy test for differentiating metallo-b-lactamase-producing isolates of Pseudomonas spp. and Acinetobacter spp. J. Clin. Microbiol. 2003, 41, 4623–4629. [Google Scholar] [CrossRef] [Green Version]
- Papagiannitsis, C.C.; Medvecky, M.; Chudejova, K.; Skalova, A.; Rotova, V.; Spanelova, P.; Jakubu, V.; Zemlickova, H.; Hrabak, J. Molecular Characterization of carbapenemase-producing Pseudomonas aeruginosa of Czech origin and evidence for clonal spread of extensively resistant Sequence Type 357 expressing IMP-7 metallo-β-lactamase. Antimicrob. Agents Chemother. 2017, 61, e01811-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curran, B.; Jonas, D.; Grundmann, H.; Pitt, T.; Dowson, C.G. Development of a multilocus sequence typing scheme for the opportunistic pathogen Pseudomonas aeruginosa. J. Clin. Microbiol. 2004, 42, 5644–5649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura, A.; Soares, M.; Pereira, C.; Leitão, N.; Henriques, I.; Correia, A. INTEGRALL: A database and search engine for integrons, integrases and gene cassettes. Bioinformatics 2009, 25, 1096–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, B.M.; Harding, G.P.; Zuccarelli, A.J. A general method for detecting and sizing large plasmids. Anal. Biochem. 1995, 226, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.D.; Jin, Q.; Chen, L.H.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Couvin, D.; Bernheim, A.; Toffano-Nioche, C.; Touchon, M.; Michalik, J.; Neron, B.; Rocha, E.P.C.; Vergnaud, G.; Gautheret, D.; Pourcel, C. CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res. 2018, 46, W246–W251. [Google Scholar] [CrossRef] [Green Version]
- Woodford, N.; Turton, J.F.; Livermore, D.M. Multiresistant Gram-negative bacteria: The role of high-risk clones in the dissemination of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 736–755. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Lambert, T.; Turkoglu, S.; Ronco, E.; Gaillard, J.L.; Nordmann, P. Characterization of class 1 integrons from Pseudomonas aeruginosa that contain the blaVIM-2 carbapenem-hydrolyzing b-lactamase gene and of two novel aminoglycoside resistance gene cassettes. Antimicrob. Agents Chemother. 2001, 45, 546–552. [Google Scholar] [CrossRef] [Green Version]
- Samuelsen, Ø.; Toleman, M.A.; Sundsfjord, A.; Rydberg, J.; Leegaard, T.M.; Walder, M.; Lia, A.; Ranheim, T.E.; Rajendra, Y.; Hermansen, N.O.; et al. Molecular epidemiology of metallo-β-lactamase-producing Pseudomonas aeruginosa isolates from Norway and Sweden shows import of international clones and local clonal expansion. Antimicrob. Agents Chemother. 2010, 54, 346–352. [Google Scholar] [CrossRef] [Green Version]
- Yano, H.; Genka, H.; Ohtsubo, Y.; Nagata, Y.; Top, E.M.; Tsuda, M. Cointegrate-resolution of toluene-catabolic transposon Tn4651: Determination of crossover site and the segment required for full resolution activity. Plasmid 2013, 69, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Breidenstein, E.B.M.; de la Fuente-Nunez, C.; Hancock, R.E.W. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Jalal, S.; Wretlind, B. Mechanisms of quinolone resistance in clinical strains of Pseudomonas aeruginosa. Microb. Drug Resist. 1998, 4, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Walsh, F.; Amyes, S.G. Carbapenem resistance in clinical isolates of Pseudomonas aeruginosa. J. Chemother. 2007, 19, 376–381. [Google Scholar] [CrossRef]
- Lanotte, P.; Watt, S.; Mereghetti, L.; Dartiguelongue, N.; Rastegar-Lari, A.; Goudeau, A.; Quentin, R. Genetic features of Pseudomonas aeruginosa isolates from cystic fibrosis patients compared with those of isolates from other origins. J. Med. Microbiol. 2004, 53, 73–81. [Google Scholar] [CrossRef]
- Azimi, S.; Kafil, H.S.; Baghi, H.B.; Shokrian, S.; Najaf, K.; Asgharzadeh, M.; Yousefi, M.; Shahrivar, F.; Aghazadeh, M. Presence of exoY, exoS, exoU and exoT genes, antibiotic resistance and biofilm production among Pseudomonas aeruginosa isolates in Northwest Iran. GMS Hyg. Infect. Control. 2016, 11, Doc04. [Google Scholar] [CrossRef]
- Roy-Burman, A.; Savel, R.H.; Racine, S.; Revadigar, N.S.; Fujimoto, J.; Sawa, T.; Frank, D.W.; Wiener-Kronish, J.P. Type III protein secretion is associated with death in lower respiratory and systemic Pseudomonas aeruginosa infections. J. Infect. Dis. 2001, 183, 1767–1774. [Google Scholar] [CrossRef] [Green Version]
- Finck-Barbancon, V.; Goranson, J.; Zhu, L.; Wiener-Kronish, J.P.; Fleiszig, S.M.; Wu, C.; Mende-Mueller, L.; Frank, D.W. ExoU expression by Pseudomonas aeruginosa correlates with acute cytotoxicity and epithelial injury. Mol. Microbiol. 1997, 25, 547–557. [Google Scholar] [CrossRef]
- Van’t Wout, E.F.; Van Schadewijk, A.; Van Boxtel, R.; Dalton, L.E.; Clarke, H.J.; Tommassen, J.; Marciniak, S.J.; Hiemstra, P.S. Virulence factors of Pseudomonas aeruginosa induce both the unfolded protein and integrated stress responses in airway epithelial cells. PLoS Pathog. 2015, 11, e1004946. [Google Scholar] [CrossRef] [Green Version]
- Jamunadevi, S.; Balashanmugam, P.; Muralitharan, G.; Kalaichelvan, P.T. Molecular characterization of pathogenic and non-pathogenic Pseudomonas aeruginosa with special reference to phenazine gene. J. Mod. Biotechnol. 2012, 1, 70–74. [Google Scholar]
- Karah, N.; Samuelsen, O.; Zarrilli, R.; Sahl, J.W.; Wai, S.N.; Uhlin, B.E. CRISPR-cas subtype I-Fb in Acinetobacter baumannii: Evolution and utilization for strain subtyping. PLoS ONE 2015, 10, e0118205. [Google Scholar] [CrossRef] [Green Version]
- van Belkum, A.; Soriaga, L.B.; LaFave, M.C.; Akella, S.; Veyrieras, J.B.; Barbu, E.M.; Shortridge, D.; Blanc, B.; Hannum, G.; Zambardi, G.; et al. Phylogenetic distribution of CRISPR-Cas systems in antibiotic-resistant Pseudomonas aeruginosa. mBio 2015, 6, e01796-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira Luz, A.C.; da Silva, J.M.A.; Rezende, A.M.; de Barros, M.P.S.; Leal-Balbino, T.C. Analysis of direct repeats and spacers of CRISPR/Cas systems type I-F in Brazilian clinical strains of Pseudomonas Aeruginosa. Mol. Genet. Genom. 2019, 294, 1095–1105. [Google Scholar] [CrossRef]
- Wright, L.L.; Turton, J.F.; Hopkins, K.L.; Livermore, D.M.; Woodford, N. Genetic environment of metallo-β-lactamase genes in Pseudomonas aeruginosa isolates from the UK. J. Antimicrob. Chemother. 2015, 70, 3250–3258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostria-Hernandez, M.L.; Sánchez-Vallejo, C.J.; Ibarra, J.A.; Castro-Escarpulli, G. Survey of clustered regularly interspaced short palindromic repeats and their associated Cas proteins (CRISPR/Cas) systems in multiple sequenced strains of Klebsiella pneumoniae. BMC Res. Notes 2015, 8, 332. [Google Scholar] [CrossRef] [Green Version]
- Hatoum-Aslan, A.; Marraffini, L.A. Impact of CRISPR immunity on the emergence and virulence of bacterial pathogens. Curr. Opin. Microbiol. 2014, 17, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Edelstein, M.V.; Skleenova, E.N.; Shevchenko, O.V.; D’souza, J.W.; Tapalski, D.V.; Azizov, I.S.; Sukhorukova, M.V.; Pavlukov, R.A.; Kozlov, R.S.; Toleman, M.A.; et al. Spread of extensively resistant VIM-2-positive ST235 Pseudomonas aeruginosa in Belarus, Kazakhstan, and Russia: A longitudinal epidemiological and clinical study. Lancet Infect. Dis. 2013, 13, 867–876. [Google Scholar] [CrossRef]
- Hrabak, J.; Bitar, I.; Papagiannitsis, C.C. Combination of mass spectrometry and DNA sequencing for detection of antibiotic resistance in diagnostic laboratories. Folia Microbiol. 2020, 65, 233–243. [Google Scholar] [CrossRef]



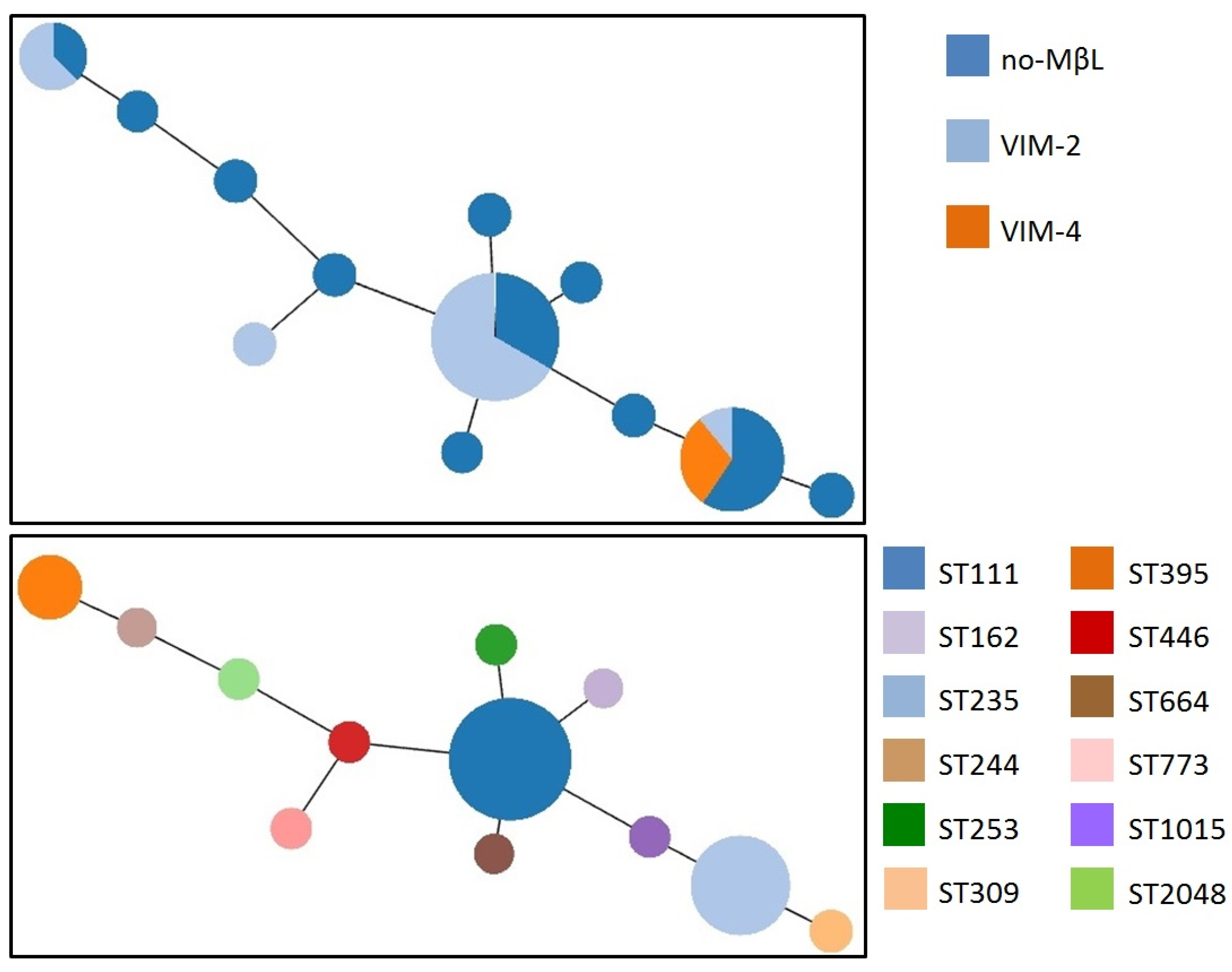{kind=link}
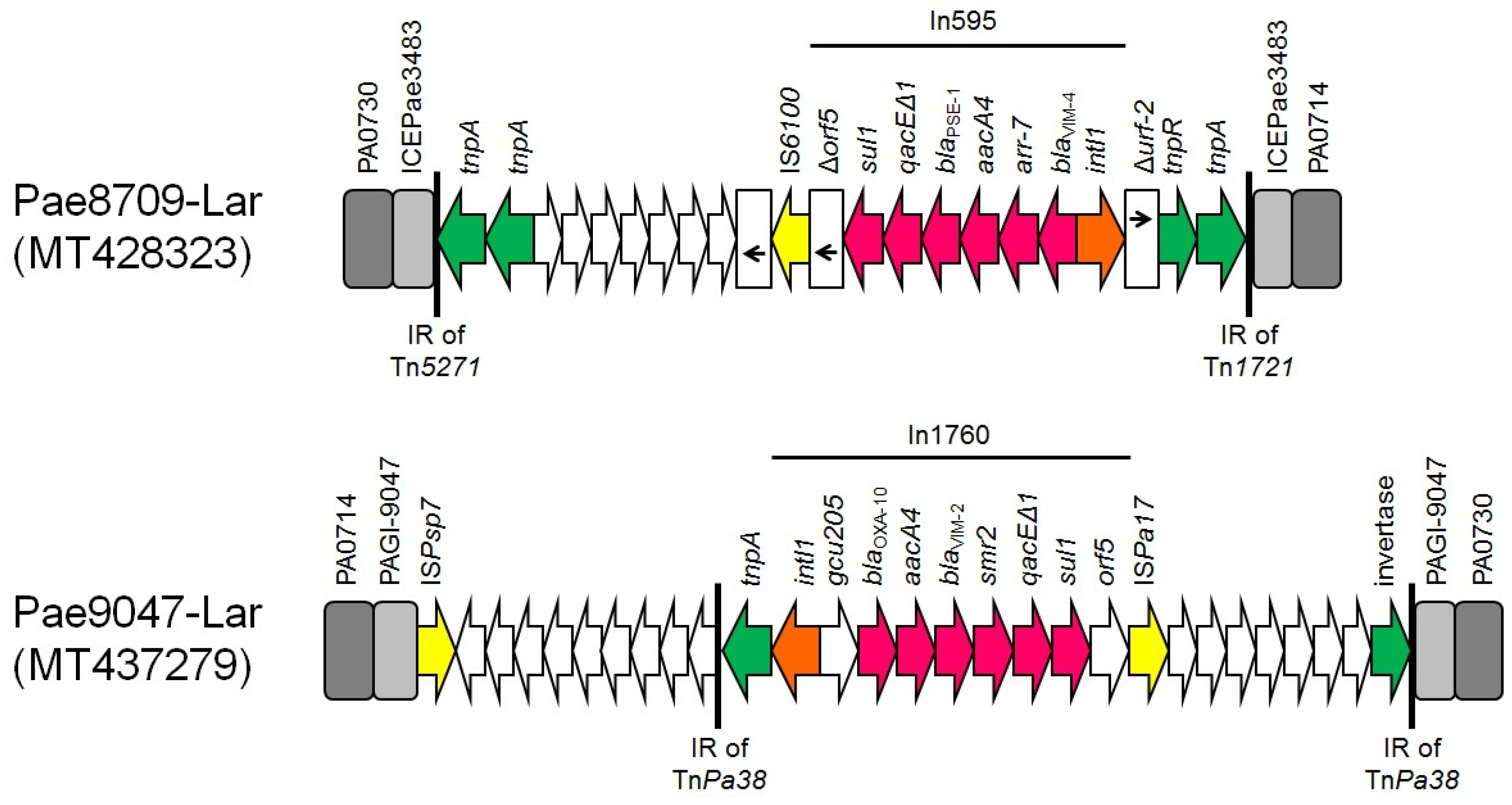{kind=link}
| ST | MBL | No. of Isolates | Integrons with blaVIM Genes |
|---|---|---|---|
| ST111 | VIM-2 | 34 | In59 |
| - | 17 | - | |
| ST162 | - | 1 | - |
| ST235 | VIM-2 | 4 | In59 |
| VIM-4 | 11 | In595 | |
| - | 22 | - | |
| ST244 | - | 1 | - |
| ST253 | - | 2 | - |
| ST309 | - | 3 | - |
| ST395 | VIM-2 | 10 | In1760 |
| - | 6 | - | |
| ST446 | - | 2 | - |
| ST664 | - | 1 | - |
| ST773 | VIM-2 | 2 | In59 |
| ST1015 | - | 2 | - |
| ST2048 | - | 2 | - |
| Isolate | ST | MBL Content | Integrons with blaVIM Genes | Additional Resistance Genes | Virulence Genes | CRISPR/Cas System a |
|---|---|---|---|---|---|---|
| Pae8397-Lar | 111 | VIM-2 | In59 | blaPAO, blaOXA-395, aph(3’)-IIb, catB7 | algD, aprA, lasB, plcH, plcN, exoS, exoT, exoY, phzM, phzS, phzH, pilA, pilB, toxA | - |
| Pae9981-Lar | 111 | VIM-2 | In59 | blaPAO, blaOXA-395, aph(3’)-IIb, catB7 | algD, aprA, lasB, plcH, plcN, exoS, exoT, exoY, phzM, phzS, phzH, pilA, pilB, toxA | - |
| Pae8709-Lar | 235 | VIM-4 | In595 | blaPAO, blaOXA-35, blaOXA-488, aph(3’)-IIb, catB7 | algD, aprA, lasB, plcH, plcN, exoT, exoU, exoY, phzM, phzS, phzH, pilB, toxA | Type I-C |
| Pae9938-Lar | 235 | VIM-4 | In595 | blaPAO, blaOXA-35, blaOXA-488, aph(3’)-IIb, catB7 | algD, aprA, lasB, plcH, plcN, exoT, exoU, exoY, phzM, phzS, phzH, pilB, toxA | Type I-C |
| Pae8414-Lar | 395 | VIM-2 | In1760 | blaPAO, blaOXA-488, aadA6, aph(3’)-IIb, aph(3’)-Via, catB7 | algD, aprA, lasB, plcH, plcN, exoS, exoT, exoY, phzM, phzS, phzH, pilB, toxA | Type I-E |
| Pae9047-Lar | 395 | VIM-2 | In1760 | blaPAO, blaOXA-488, aadA6, aph(3’)-IIb, aph(3’)-Via, catB7 | algD, aprA, lasB, plcH, plcN, exoS, exoT, exoY, phzM, phzS, phzH, pilA, pilB, toxA | Type I-E |
| Pae112-Lar | 162 | No | - | blaPAO, blaOXA-494, aacA29, aph(3’)-IIb, catB7, sul1 | algD, aprA, lasB, plcH, plcN, exoT, exoU, exoY, phzM, phzS, phzH, pilB, toxA | - |
| Pae69-Lar | 235 | No | - | blaPAO, blaOXA-2, blaOXA-488, blaPER-1, aacA4, aadA2, ant(2’’)-Ia, aph(3’)-IIb, strA, strB, sul1 | algD, aprA, lasB, plcH, plcN, exoT, exoU, exoY, phzM, phzS, phzH, pilB, toxA | - |
| Pae90-Lar | 235 | No | - | blaPAO, blaOXA-35, blaOXA-488, aacA4, aph(3’)-IIb, catB7, sul1 | algD, aprA, lasB, plcH, plcN, exoT, exoU, exoY, phzM, phzS, phzH, pilB, toxA | Type I-C |
| Pae91-Lar | 244 | No | - | blaPAO, blaOXA-494, aph(3’)-IIb, catB7 | algD, aprA, lasB, plcH, plcN, exoT, exoU, exoY, phzM, phzS, phzH, pilB, toxA | Type I-F |
| Pae462-Lar | 253 | No | - | blaPAO, blaOXA-488, aph(3’)-IIb, catB7 | algD, aprA, lasB, plcH, plcN, exoT, exoU, exoY, phzM, phzS, phzH, pilB, toxA | Type I-F |
| Pae100-Lar | 299 | No | - | blaPAO, blaOXA-494, aph(3’)-IIb, catB7 | algD, aprA, lasB, plcH, plcN, exoS, exoT, exoY, phzM, phzS, phzH, pilB, toxA | - |
| Pae86-Lar | 446 | No | - | blaPAO, blaOXA-395, aph(3’)-IIb, catB7 | algD, aprA, lasB, plcH, plcN, exoT, exoU, exoY, phzM, phzS, phzH, pilA, pilB, toxA | - |
| Pae64-Lar | 664 | No | - | blaPAO, blaOXA-50, aph(3’)-IIb, catB7 | algD, aprA, lasB, plcH, plcN, exoS, exoT, exoY, phzM, phzS, phzH, pilB, toxA | - |
| Pae475-Lar | 2048 | No | - | blaPAO, blaOXA-494, aph(3’)-IIb, catB7 | algD, aprA, lasB, plcH, plcN, exoS, exoT, exoY, phzM, phzS, phzH, pilA, pilB, toxA | - |
| Isolate | ST | MBL Content | MIC (μg/mL) 1 | Amino Acid Changes 2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Imp | Mem | Cip | GyrA 3 | GyrB 4 | ParC | ParE | MexR | NfxB | MexT | OprD 5 | |||
| Pae8397-Lar | 111 | VIM-2 | >16 | >16 | >4 | T83I, V671I, G860S, D893E, A900G, S903A, +S912, +E913 | SB1 | S87L, F254V, A346Q | T89I, I91T | V126E | E124A | L26V | * 94 |
| Pae9981-Lar | 111 | VIM-2 | >16 | >16 | >4 | T83I, V671I, G860S, D893E, A900G, S903A, +S912, +E913 | SB1 | S87L, F254V, A346Q | T89I, I91T | V126E | E124A | L26V | Dis337 |
| Pae8709-Lar | 235 | VIM-4 | >16 | >16 | >4 | T83I | SB1, E469D | S87L, F254V, A346Q | T89I, I91T, D533E | V126E | E124A | L26V | * 94 |
| Pae9938-Lar | 235 | VIM-4 | >16 | >16 | >4 | T83I | SB1 | S87L, F254V, A346Q | T89I, I91T, D533E | V126E | E124A | L26V | Non-SC |
| Pae8414-Lar | 395 | VIM-2 | >16 | >16 | >4 | T83I, +E909, +S910 | SB1 | S87L, F254V, A346Q | T89I, I91T, V200M | - | E124A | L26V | * 229 |
| Pae9047-Lar | 395 | VIM-2 | >16 | >16 | >4 | T83I, +E909, +S910 | SB1 | S87L, F254V, A346Q | T89I, I91T, V200M | - | E124A | L26V | Non-SC |
| Pae112-Lar | 162 | No | >16 | >16 | 1 | T83I | SB1 | F254V, A346Q | T89I, I91T, A473V, D533E | - | E124A | L26V | * 219 |
| Pae69-Lar | 235 | No | >16 | >16 | >4 | T83I | SB1, P750S | S87L, F254V, A346Q | T89I, I91T, D533E | V126E | E124A | L26V | FR501 |
| Pae90-Lar | 235 | No | >16 | >16 | >4 | T83I | SB1 | S87L, F254V, A346Q | T89I, I91T, D533E | V126E | E124A | L26V | * 142 |
| Pae91-Lar | 244 | No | >16 | 4 | ≤0.25 | - | SB1 | F254V, A346Q | T89I, I91T | - | E124A | L26V | Non-SC |
| Pae462-Lar | 253 | No | >16 | >16 | 1 | +E909, +S910 | SB1 | F254V, A346Q | T89I, I91T, A473V, D533E | G91E, V126E | R21H, D56G, E124A | L26V | FR501 |
| Pae100-Lar | 299 | No | >16 | 8 | ≤0.25 | - | SB1 | F254V, A346Q, V646L | T89I, I91T, D533E | - | E124A | L26V | Non-SC |
| Pae86-Lar | 446 | No | >16 | 4 | 2 | +E909, +S910 | SB1 | F254V, A346Q, A587T | T89I, I91T, D533E | V126E | E124A | L26V | Non-SC |
| Pae64-Lar | 664 | No | 2 | 12 | 1 | - | SB1 | F254V, H262Q, A346Q | T89I, I91T, E215Q | * 94 | E124A | L26V | Non-SC |
| Pae475-Lar | 2048 | No | >16 | 4 | ≤0.25 | - | SB1 | F254V, A346Q | T89I, I91T | - | R82L, E124A | L26V | * 424 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papagiannitsis, C.C.; Verra, A.; Galani, V.; Xitsas, S.; Bitar, I.; Hrabak, J.; Petinaki, E. Unravelling the Features of Success of VIM-Producing ST111 and ST235 Pseudomonas aeruginosa in a Greek Hospital. Microorganisms 2020, 8, 1884. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121884
Papagiannitsis CC, Verra A, Galani V, Xitsas S, Bitar I, Hrabak J, Petinaki E. Unravelling the Features of Success of VIM-Producing ST111 and ST235 Pseudomonas aeruginosa in a Greek Hospital. Microorganisms. 2020; 8(12):1884. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121884
Chicago/Turabian StylePapagiannitsis, Costas C., Aggeliki Verra, Vasiliki Galani, Stelios Xitsas, Ibrahim Bitar, Jaroslav Hrabak, and Efthymia Petinaki. 2020. "Unravelling the Features of Success of VIM-Producing ST111 and ST235 Pseudomonas aeruginosa in a Greek Hospital" Microorganisms 8, no. 12: 1884. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121884