The Dysbiosis of Gut Microbiota Caused by Low-Dose Cadmium Aggravate the Injury of Mice Liver through Increasing Intestinal Permeability

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
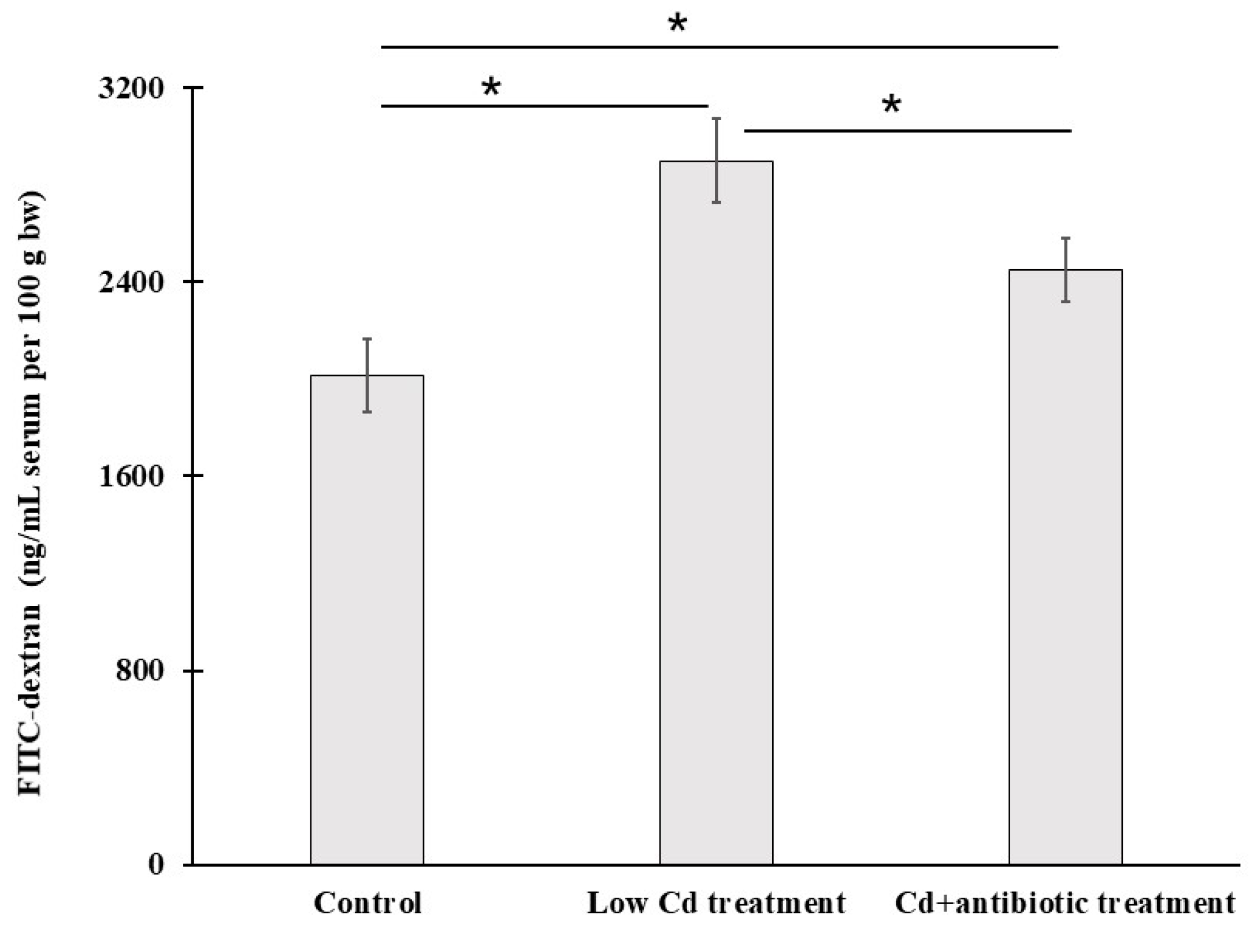
2.2. Intestinal Permeability In Vivo
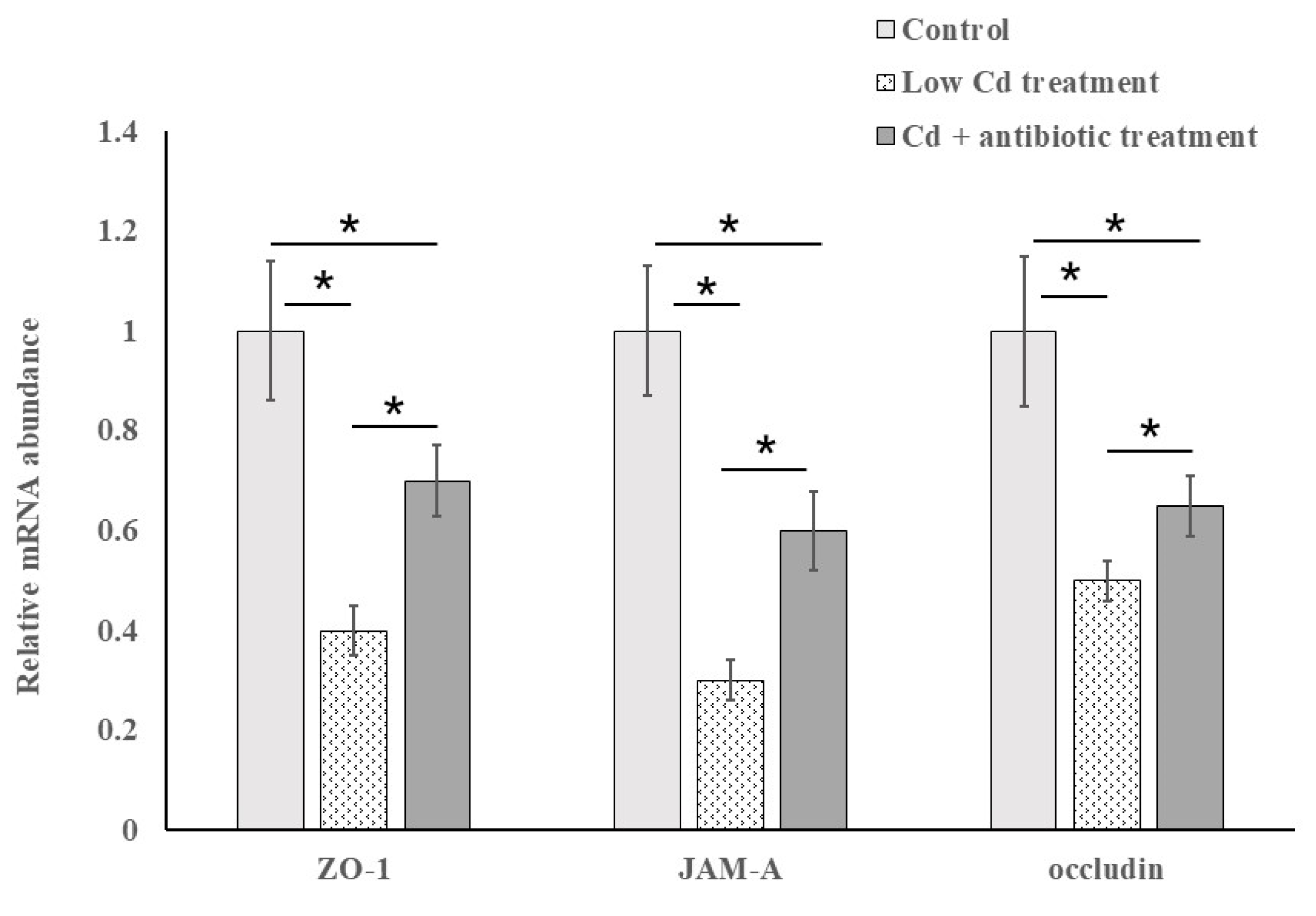
2.3. RNA Extraction and Quantitative Polymerase Chain Reaction (qPCR) Amplification of the Genes for Tight-Junction Proteins
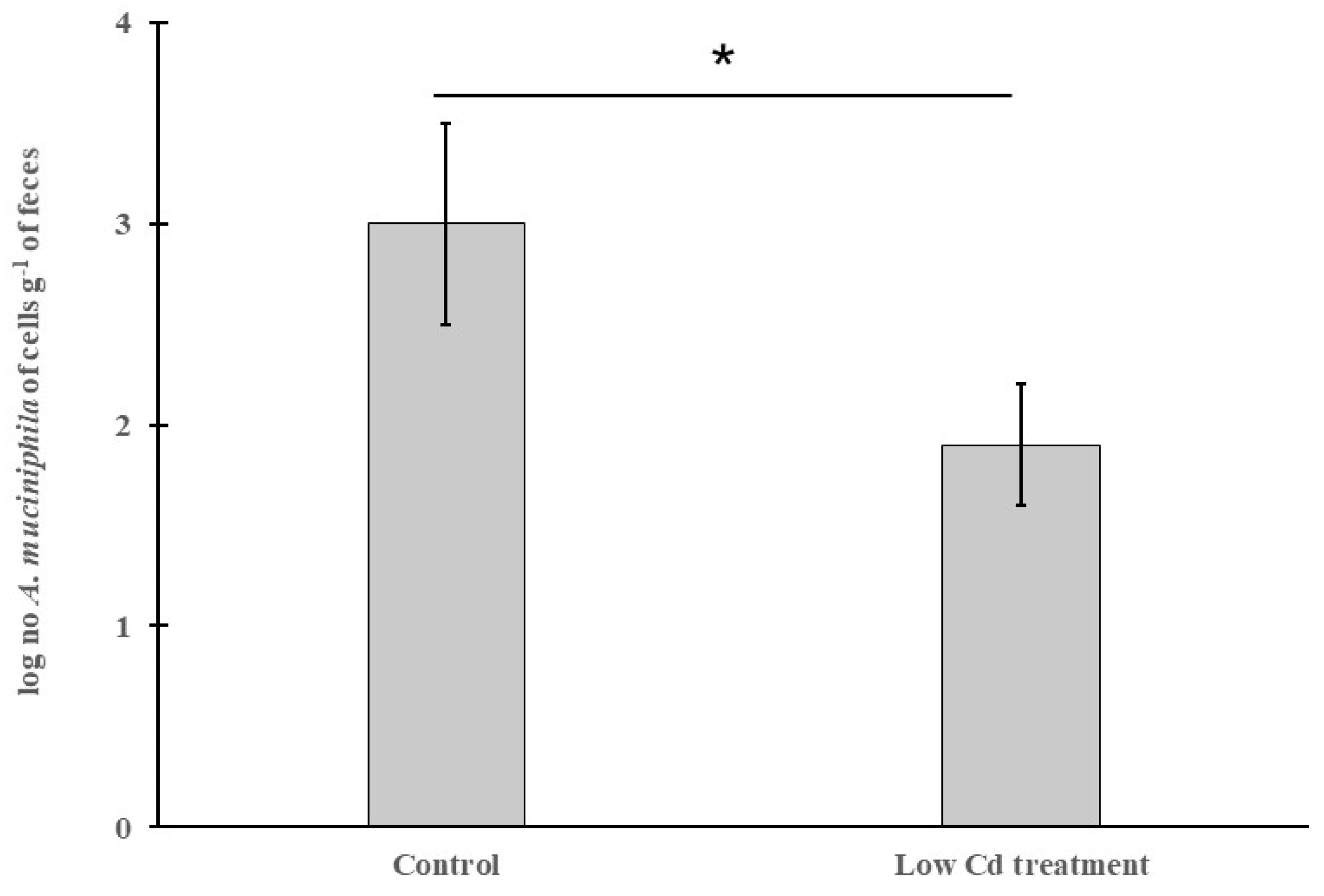
2.4. Quantitative PCR Amplification of Akkermansia Muciniphila and the Overall Microbe Census
2.5. High-Throughput RNA Sequencing
2.6. The Analyses of Gut Microbiota
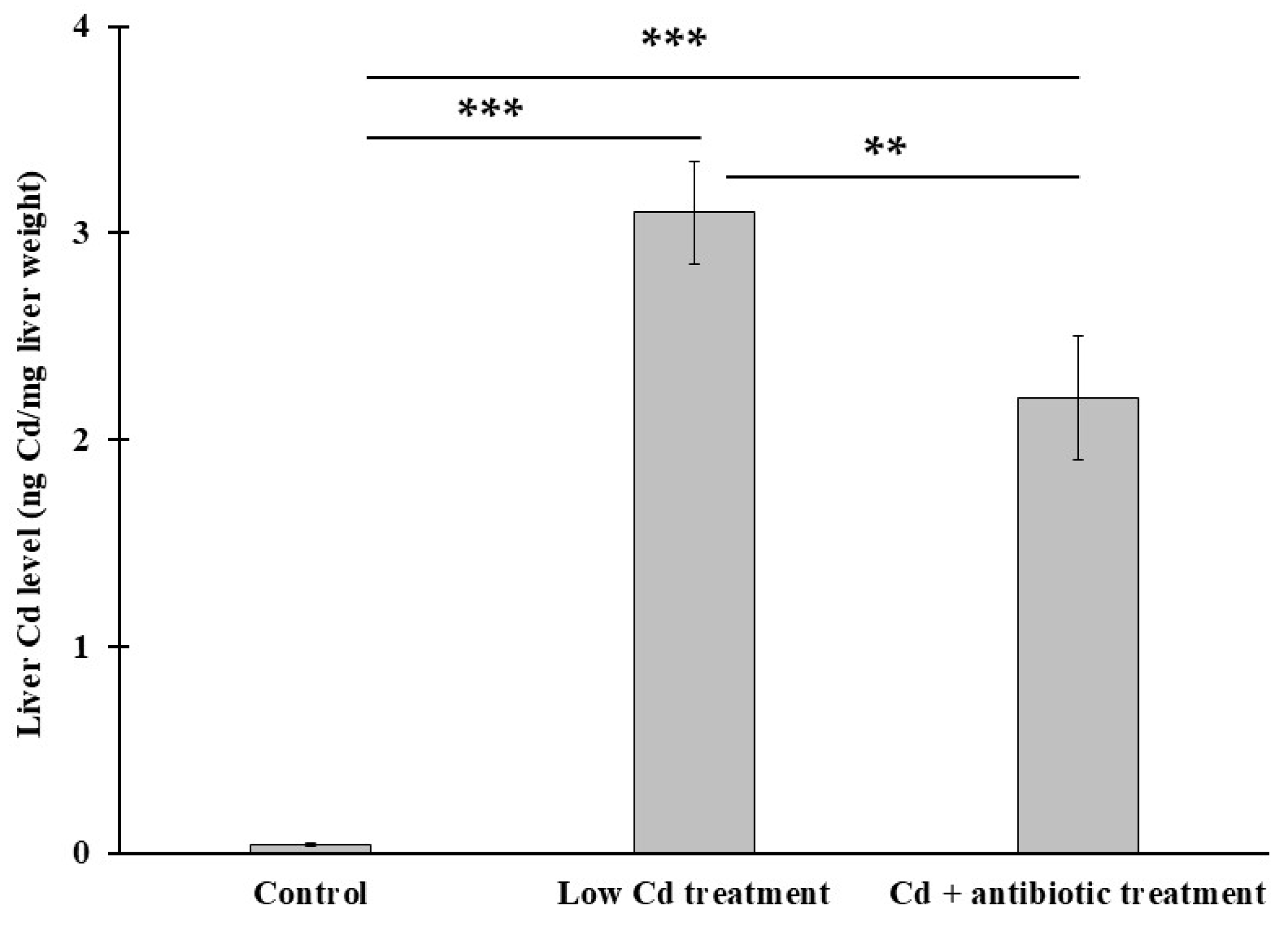
2.7. Determination of Cd Concentration in Mouse Liver
2.8. Enzyme-Linked Immunosorbent Assay (ELISA)
2.9. Statistical Analysis
3. Results
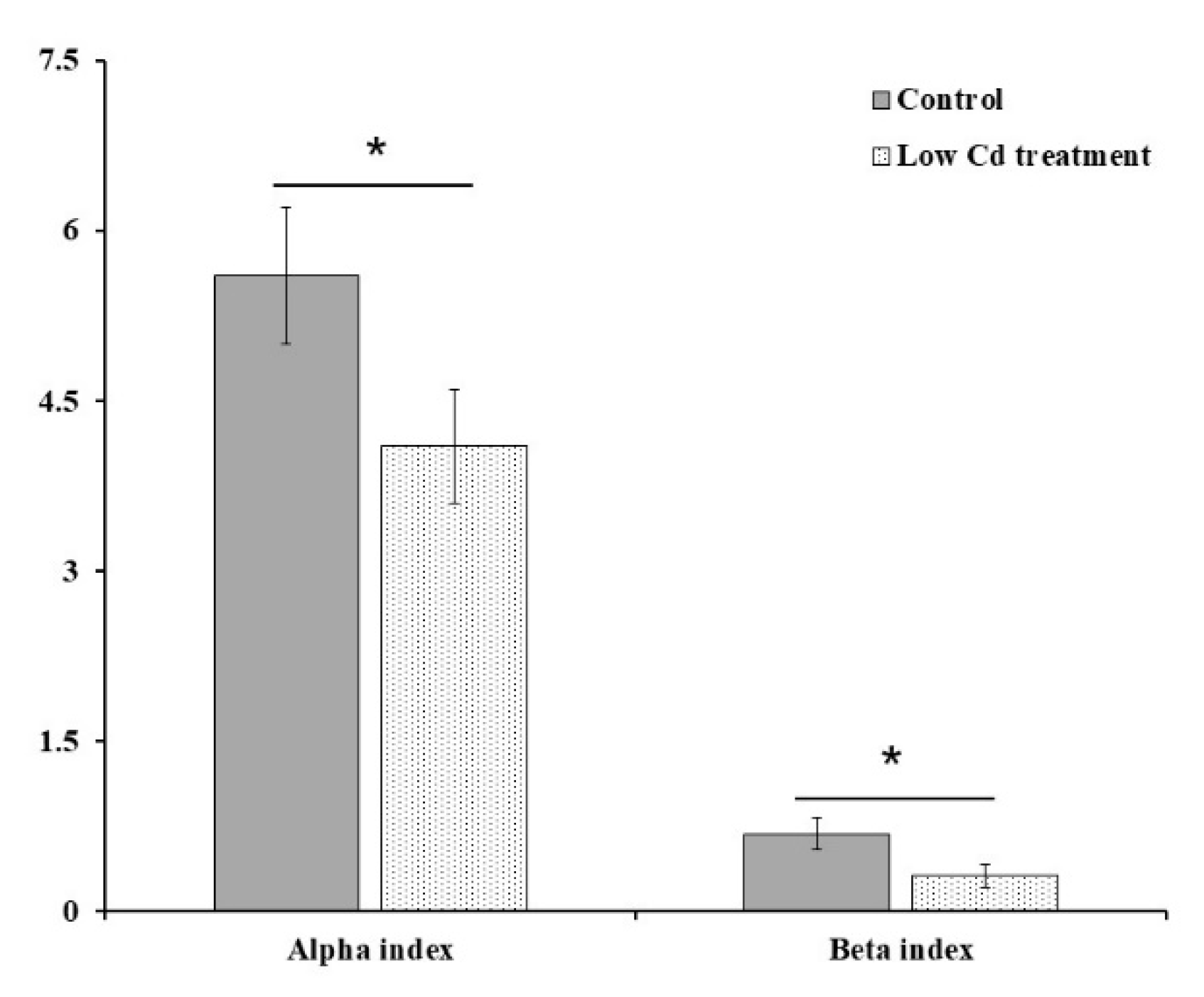
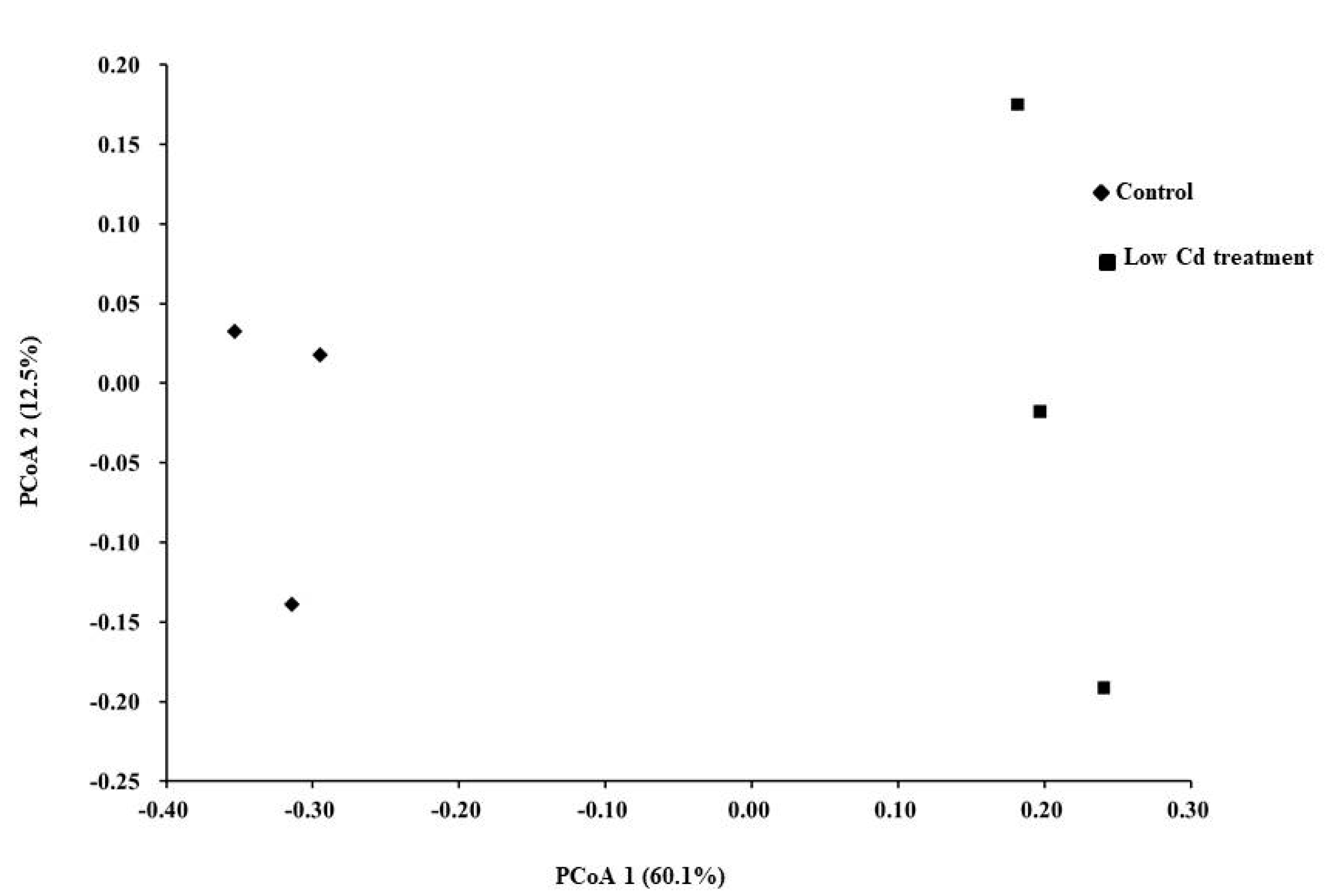
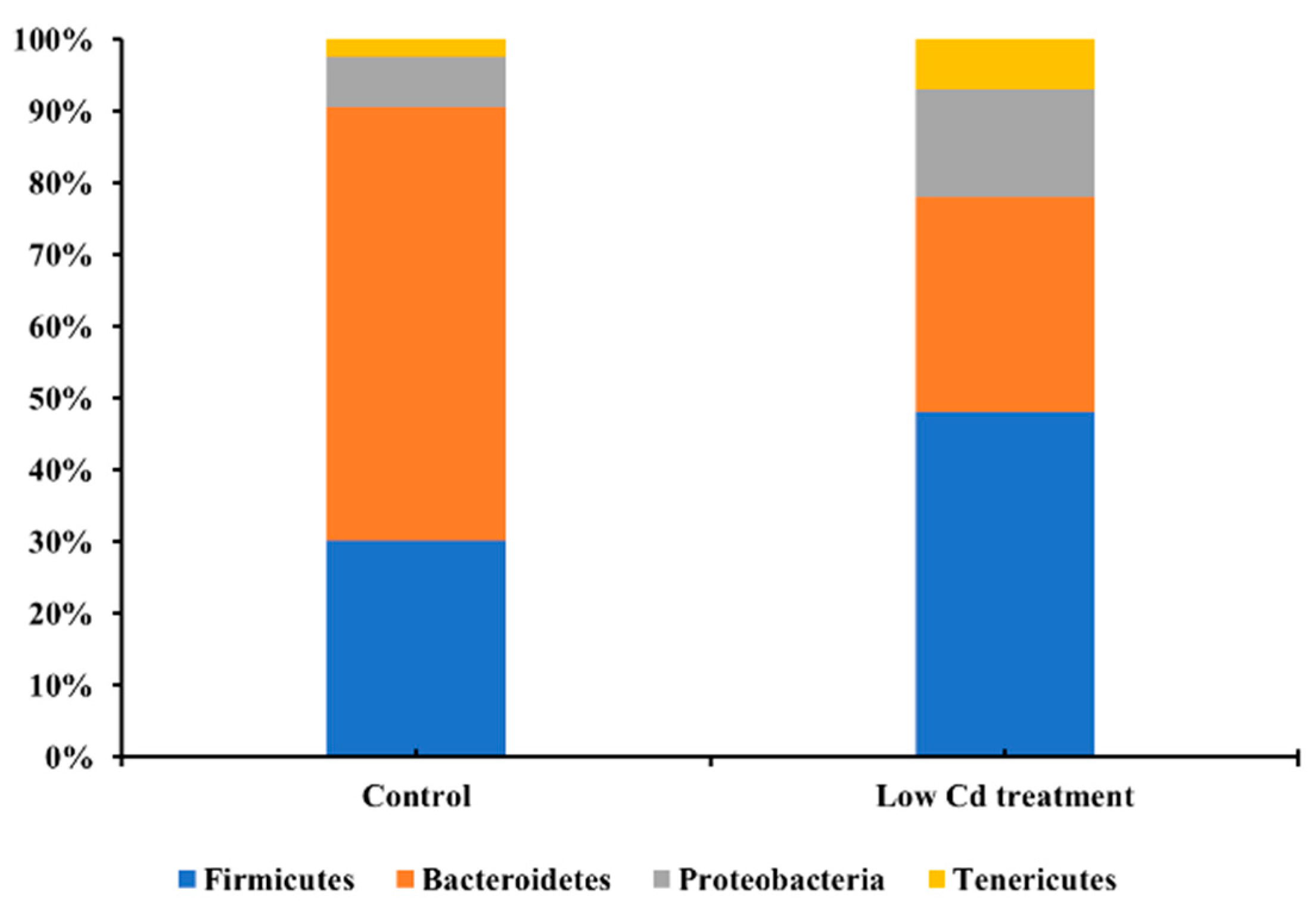
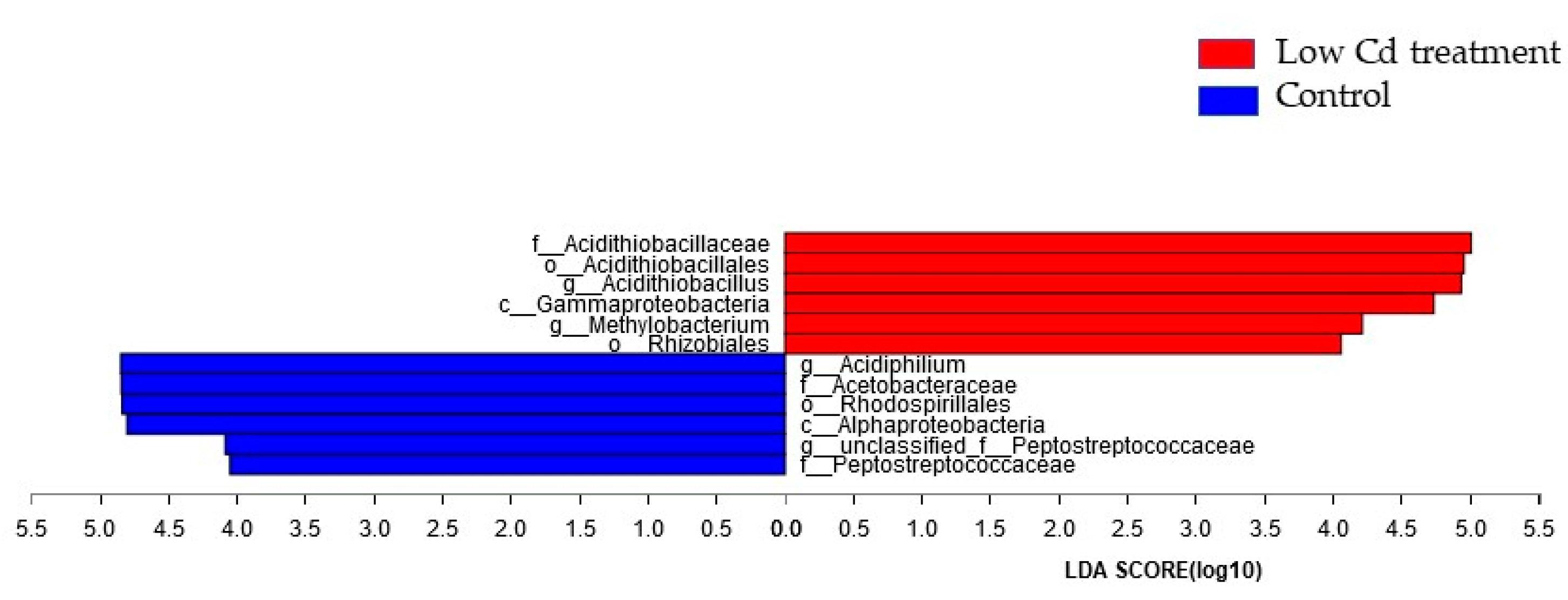
3.1. Oral Low-Level Cd Causes the Alteration of Gut Microbiota
3.2. Intestinal Permeability Is Increased after Cd Exposure
3.3. The Expression of Tight-Junction Proteins Is Closely Related to Intestinal Permeability
3.4. The Abundance of A. Mucinipbila Is Negatively Correlated to Intestinal Permeability
3.5. The Presence of Gut Microbiota Can Modulate Cd Level in the Liver
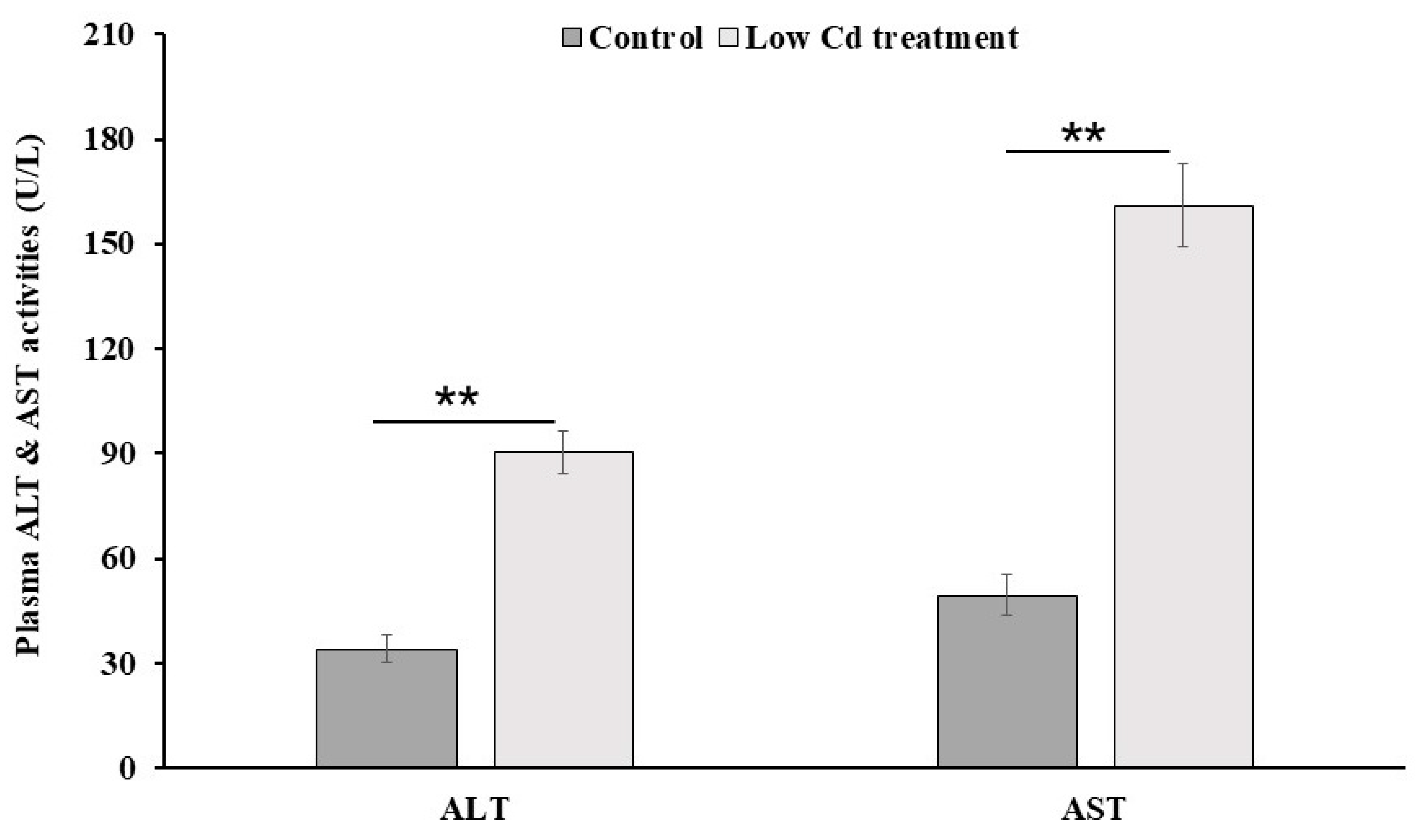
3.6. Mouse Liver Is Injured by Low-Level Cd
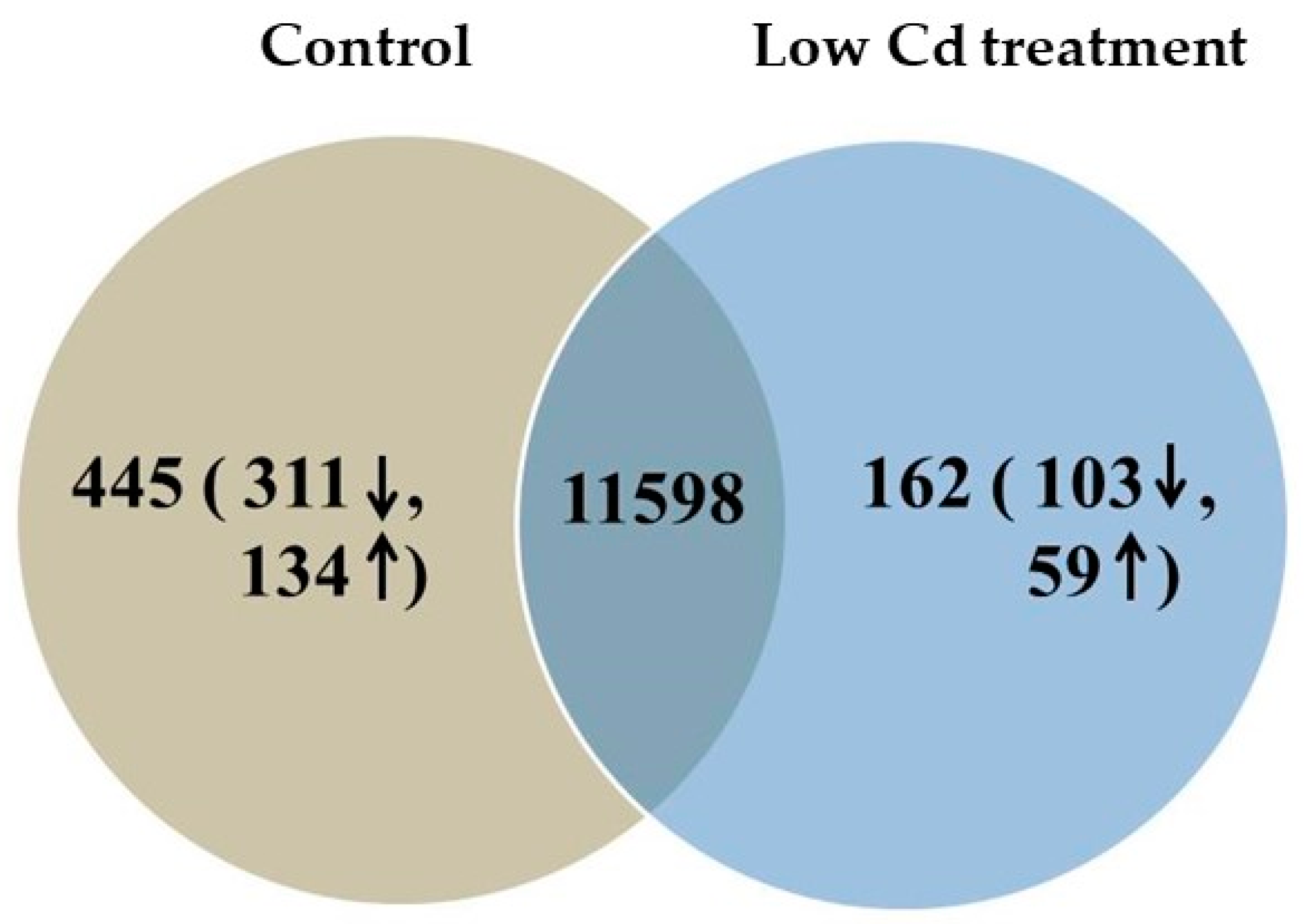
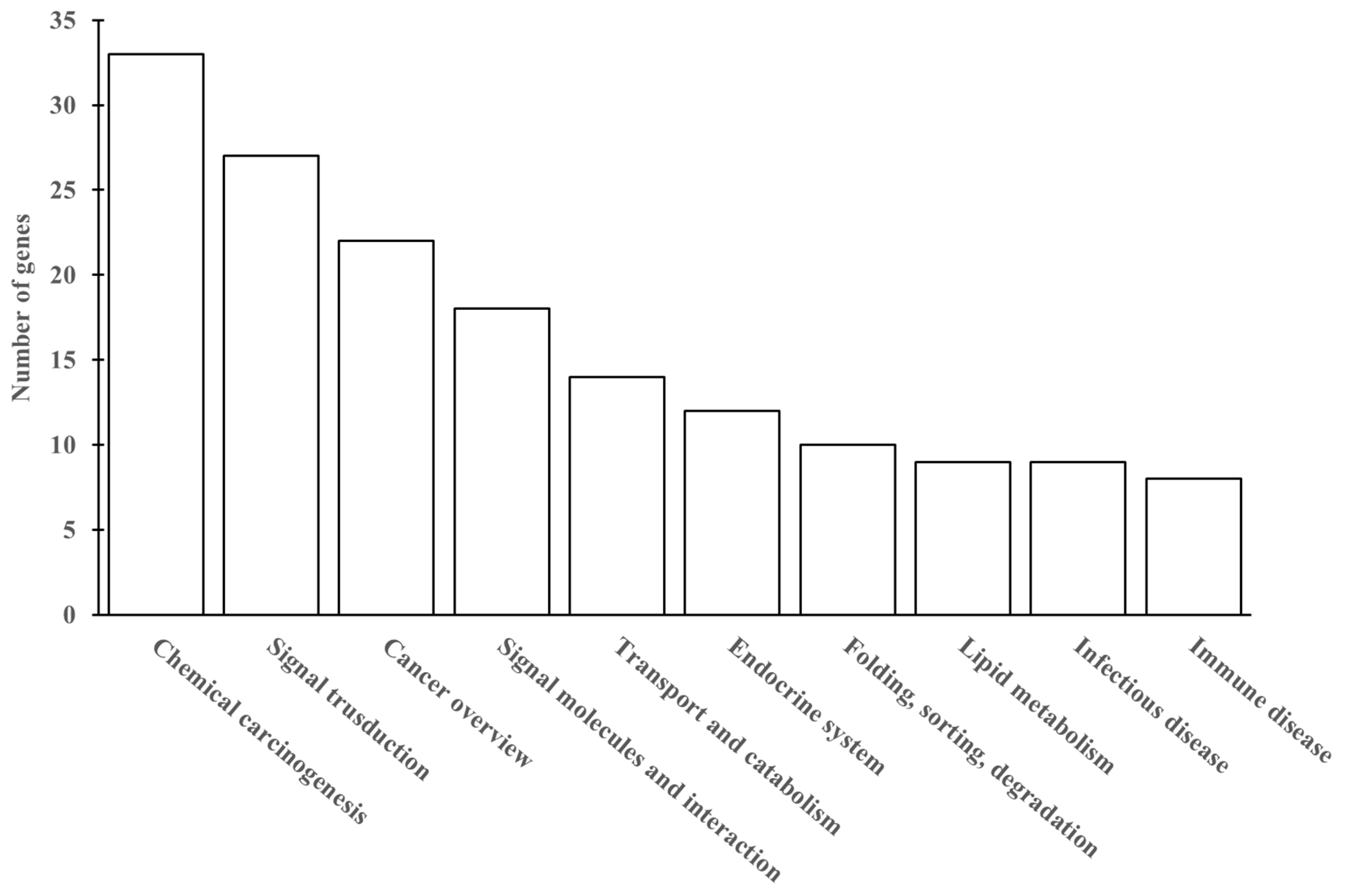
3.7. Cd Accumulation Induces Hepatic Genes Expression Differently
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Feng, W.; Guo, Z.; Xiao, X.; Peng, C.; Shi, L.; Ran, H.; Xu, W. Atmospheric deposition as a source of cadmium and lead to soil-rice system and associated risk assessment. Ecotoxicol. Environ. Saf. 2019, 180, 160–167. [Google Scholar] [CrossRef]
- Satarug, S.; Vesey, D.A.; Gobe, G.C. Current health risk assessment practice for dietary cadmium: Data from different countries. Food. Chem. Toxicol. 2017, 106, 430–445. [Google Scholar] [CrossRef] [PubMed]
- Mezynska, M.; Brzoska, M.M. Environmental exposure to cadmium—A risk for health of the general population in industrialized countries and preventive strategies. Environ. Sci. Pollut. Res. Int. 2018, 25, 3211–3232. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yang, M.; Wang, P.; Wang, Z.; Li, M.; Zhao, F.J. Dietary cadmium intake from rice and vegetables and potential health risk: A case study in Xiangtan, southern China. Sci. Total. Environ. 2018, 639, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Mu, T.; Wu, T.; Zhou, T.; Li, Z.; Ouyang, Y.; Jiang, J. Geographical variation in arsenic, cadmium, and lead of soils and rice in the major rice producing regions of China. Sci. Total. Environ. 2019, 677, 373–381. [Google Scholar] [CrossRef]
- Egger, A.E.; Grabmann, G.; Pechriggl, E.J.; Artner, C.; Hartinger, C.G.; Fritsch, H.; Keppler, B.K. Chemical imaging and assessment of cadmium distribution in the human body. Metallomics 2019, 11, 2010–2019. [Google Scholar] [CrossRef] [Green Version]
- Riazi, K.; Raman, M. Dietary patterns and components in nonalcoholic fatty liver disease (nafld): What key messages can health care providers offer? Nutrients 2019, 11, 2878. [Google Scholar] [CrossRef] [Green Version]
- Hyder, O.; Chung, M.; Cosgrove, D.; Herman, J.M.; Li, Z.; Firoozmand, A.; Gurakar, A.; Pawlik, T.M. Cadmium exposure and liver disease among US adults. J. Gastrointest. Surg. 2013, 17, 1265–1273. [Google Scholar] [CrossRef]
- Go, Y.M.; Sutliff, R.L.; Chandler, J.D.; Khalidur, R.; Kang, B.Y.; Anania, F.A.; Jones, D.P. Low-dose cadmium causes metabolic and genetic dysregulation associated with fatty liver disease in mice. Toxicol. Sci. 2015, 147, 524–534. [Google Scholar] [CrossRef] [Green Version]
- Ba, Q.; Li, M.; Chen, P.; Huang, C.; Duan, X.; Lu, L.; Wu, Y. Sex-dependent effects of cadmium exposure in early life on gut microbiota and fat accumulation in mice. Environ. Health Perspect. 2017, 125, 437–446. [Google Scholar] [CrossRef]
- Xia, Y.; Li, J.; Ren, W.K.; Feng, Z.M.; Huang, R.L.; Yin, Y.L. Transcriptomic analysis on responses of the liver and kidney of finishing pigs fed cadmium contaminated rice. J. Sci. Food Agric. 2018, 98, 2964–2972. [Google Scholar] [PubMed]
- Sanchez-Tapia, M.; Tovar, A.R.; Torres, N. Diet as regulator of gut microbiota and its role in health and disease. Arch. Med. Res. 2019, 50, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Karlund, A.; Gomez-Gallego, C.; Turpeinen, A.M.; Palo-Oja, O.M.; El-Nezami, H.; Kolehmainen, M. Protein supplements and their relation with nutrition, microbiota composition and health: Is more protein always better for sportspeople? Nutrients 2019, 11, 829. [Google Scholar] [CrossRef] [Green Version]
- Ganal-Vonarburg, S.C.; Duerr, C.U. The interaction of intestinal microbiota and innate lymphoid cells in health and disease throughout life. Immunology 2019, 159, 39–51. [Google Scholar] [CrossRef]
- Frazier, T.H.; DiBaise, J.K.; McClain, C.J. Gut microbiota, intestinal permeability, obesity-induced inflammation, and liver injury. J. Parenter. Enter. Nutr. 2011, 35 (Suppl. 5), 14S–20S. [Google Scholar] [CrossRef]
- Moreira, A.P.; Texeira, T.F.; Ferrira, A.B.; Peluzio, C.; Alfenas, C. Influence of a high-fat diet on gut microbiota, intestinal permeability and metabolic endotoxaemia. Br. J. Nutr. 2012, 108, 801–809. [Google Scholar] [CrossRef]
- Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, A.C.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Albillos, A.; Gottardi, A.; Rescigno, M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J. Hepatol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, N.; Kawada, N. Role of the Gut-Liver Axis in Liver Inflammation, Fibrosis, and Cancer: A Special Focus on the Gut Microbiota Relationship. Hepatol. Commun. 2019, 3, 456–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarrinpar, A.; Chaix, A. Antibiotic-induced microbiome depletion alters metabolic homeostasis by affecting gut signaling and colonic metabolism. Nat. Commun. 2018, 9, 2872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Liu, K.Y.; Shen, J.; Liu, Y.H. Wheat bran intake can attenuate chronic cadmium toxicity in mice gut microbiota. Food Funct. 2016, 7, 3524–3530. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Huttley, G.A. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuczynski, J.; Stombaugh, J.; Walters, W.A.; Gonzalez, A.; Caporaso, J.G.; Knight, R. Using QIIME to analyze 16S rRNA gene sequences from microbial communities. Curr. Protoc. Microbiol. 2012, 27, 1E.5.1–1E.5.20. [Google Scholar]
- Liu, Y.; Li, Y.H.; Liu, K.Y.; Shen, J. Exposing to cadmium stress cause profound toxic effect on microbiota of the mice intestinal tract. PLoS ONE 2014, 9, e85323. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Lan, T.; Yuan, B.; Chen, J.; Hu, J.; Ren, W.; Chen, Z. Cadmium-induced microsatellite instability in the kidneys and leukocytes of C57BL/6J mice. Environ. Toxicol. 2015, 30, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Qamar, N.; Rehman, Y. Arsenic-resistant and plant growth-promoting Firmicutes and gamma-Proteobacteria species from industrially polluted irrigation water and corresponding cropland. J. Appl. Microbiol. 2017, 123, 748–758. [Google Scholar] [CrossRef]
- Garavaglia, L.; Cerdeira, S.B.; Vullo, D.L. Chromium (VI) biotransformation by beta- and gamma-Proteobacteria from natural polluted environments: A combined biological and chemical treatment for industrial wastes. J. Hazard. Mater. 2010, 175, 104–110. [Google Scholar] [CrossRef]
- Cole, J.K.; Morton, B.R.; Cardamone, H.C.; Lake, H.R.; Dohnalkova, A.C.; Kim, Y.M.; Romine, M.F. Salinivirga fredricksonii gen. nov.; sp. nov.; a heterotrophic halophile isolated from a photosynthetic mat, a member of a novel lineage (Salinarimonadaceae fam. nov.) within the order Rhizobiales, and reclassification of the genus Salinarimonas. Int. J. Syst. Evol. Microbiol. 2018, 68, 1591–1598. [Google Scholar] [CrossRef]
- Chanratana, M.; Joe, M.M.; Choudhury, A.R.; Anandham, R.; Krishnamoorthy, R.; Kim, K.; Sa, T. Physiological response of tomato plant to chitosan-immobilized aggregated Methylobacterium oryzae CBMB20 inoculation under salinity stress. 3 Biotech 2019, 9, 397. [Google Scholar] [CrossRef]
- Alcantara-Martinez, N.; Figueroa-Martinez, F.; Rivera-Cabrera, F.; Gutierrez-Sanchez, G.; Volke-Sepulveda, T. An endophytic strain of Methylobacterium sp. increases arsenate tolerance in Acacia farnesiana (L.) willd: A proteomic approach. Sci. Total Environ. 2018, 625, 762–774. [Google Scholar] [CrossRef]
- Boden, R.; Hutt, L.P.; Huntemann, M.; Clum, A.; Pillay, M.; Palaniappan, K.; Ngan, C.Y. Permanent draft genome of Thermithiobaclillus tepidarius DSM 3134(T), a moderately thermophilic, obligately chemolithoautotrophic member of the Acidithiobacillia. Stand. Genom. Sci. 2016, 11, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, W.; Chin, Y.; Chen, X.; Mi, Y.; Xue, C.; Wang, Y.; Tang, Q. The role of gut microbiota in the resistance to obesity in mice fed a high fat diet. Int. J. Food Sci. Nutr. 2019, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, N.; Ma, Y.; Wen, D. Hydroxytyrosol improves obesity and insulin resistance by modulating gut microbiota in high-fat diet-induced obese mice. Front. Microbiol. 2019, 10, 390. [Google Scholar] [CrossRef] [PubMed]
- Woting, A.; Blaut, M. Small intestinal permeability and gut-transit time determined with low and high molecular weight fluorescein isothiocyanate-dextrans in C3H mice. Nutrients 2018, 10, 685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Zhang, D.; Han, W.; Guo, C. High-mobility group box-1 inhibition stabilizes intestinal permeability through tight junctions in experimental acute necrotizing pancreatitis. Inflamm. Res. 2019, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.A.; Hou, Y.; Yi, D.; Qiu, Y.; Wu, G.; Kong, X.; Yin, Y. Autophagy and tight junction proteins in the intestine and intestinal diseases. Anim. Nutr. 2015, 1, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Uchiyama, K.; Takagi, T. A next-generation beneficial microbe: Akkermansia muciniphila. J. Clin. Biochem. Nutr. 2018, 63, 33–35. [Google Scholar] [CrossRef] [Green Version]
- Ottman, N.; Geerlings, S.Y.; Aalvink, S.; de Vos, W.M.; Belzer, C. Action and function of Akkermansia muciniphila in microbiome ecology, health and disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 637–642. [Google Scholar] [CrossRef] [Green Version]
- Chelakkot, C.; Choi, Y.; Kim, D.K.; Park, H.T.; Ghim, J.; Kwon, Y.; Park, H.S. Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Exp. Mol. Med. 2018, 50, e450. [Google Scholar] [CrossRef]
- Hung, M.; Su, L.; Yang, L.; Zhu, L.; Liu, Z.; Duan, R. Effect of exogenous TGF-beta1 on the cadmium-induced nephrotoxicity by inhibiting apoptosis of proximal tubular cells through PI3K-AKT-mTOR signaling pathway. Chem. Biol. Interact. 2017, 269, 25–32. [Google Scholar] [CrossRef]
- Huff, M.O.; Todd, S.L.; Smith, A.L.; Elpers, J.T.; Smith, A.P.; Murphy, R.D.; Klinge, C.M. Arsenite and cadmium activate mapk/erk via membrane estrogen receptors and g-protein coupled estrogen receptor signaling in human lung adenocarcinoma cells. Toxicol. Sci. 2016, 152, 62–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firouzian, F.; Pourshoja, P.; Nili-Ahmadabadi, A.; Ranjbar, A. Hepatoprotective effect of N-acetylcystein loaded niosomes on liver function in paraquat-induced acute poisoning. Pestic. Biochem. Physiol. 2019, 160, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Swarup, D.; Naresh, R.; Varshney, V.; Kumar, P.; Nandi, D.; Patra, R. Changes in plasma hormones profile and liver function in cows naturally exposed to lead and cadmium around different industrial areas. Res. Vet. Sci. 2007, 82, 16–21. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Low Cd Treatment | |
|---|---|---|
| Total reads (×105) | 581.04 ± 33.84 | 563.01 ± 35.14 |
| Total mapped reads | 564.28 ± 35.34 | 545.29 ± 33.32 |
| Mapped to reference genome % | 92.76 | 93.26 |
| Mapped to gene % | 97.54 | 97.35 |
| Mapped to exon % | 95.45 | 95.61 |
| Mapped to intergene % | 2.35 | 2.71 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Li, Y.; Xia, Y.; Liu, K.; Ren, L.; Ji, Y. The Dysbiosis of Gut Microbiota Caused by Low-Dose Cadmium Aggravate the Injury of Mice Liver through Increasing Intestinal Permeability. Microorganisms 2020, 8, 211. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8020211
Liu Y, Li Y, Xia Y, Liu K, Ren L, Ji Y. The Dysbiosis of Gut Microbiota Caused by Low-Dose Cadmium Aggravate the Injury of Mice Liver through Increasing Intestinal Permeability. Microorganisms. 2020; 8(2):211. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8020211
Chicago/Turabian StyleLiu, Yehao, Yuhui Li, Yuhong Xia, Kaiyong Liu, Lingling Ren, and Yanli Ji. 2020. "The Dysbiosis of Gut Microbiota Caused by Low-Dose Cadmium Aggravate the Injury of Mice Liver through Increasing Intestinal Permeability" Microorganisms 8, no. 2: 211. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8020211