In Vitro Probiotic Potential and Safety Evaluation (Hemolytic, Cytotoxic Activity) of Bifidobacterium Strains Isolated from Raw Camel Milk

 , , ,
, , ,
Abstract
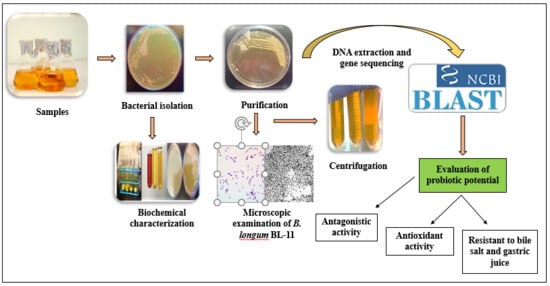
:
1. Introduction
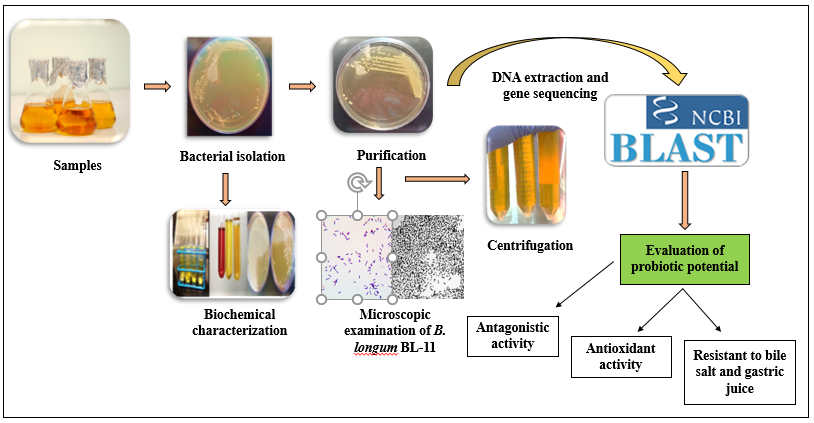
2. Materials and Methods
2.1. Chemicals
2.2. Sample Collection and Isolation
2.3. Preliminary Identification and Screening of Bifidobacterium
2.4. In Vitro Screening of Probiotic Potential of Isolates
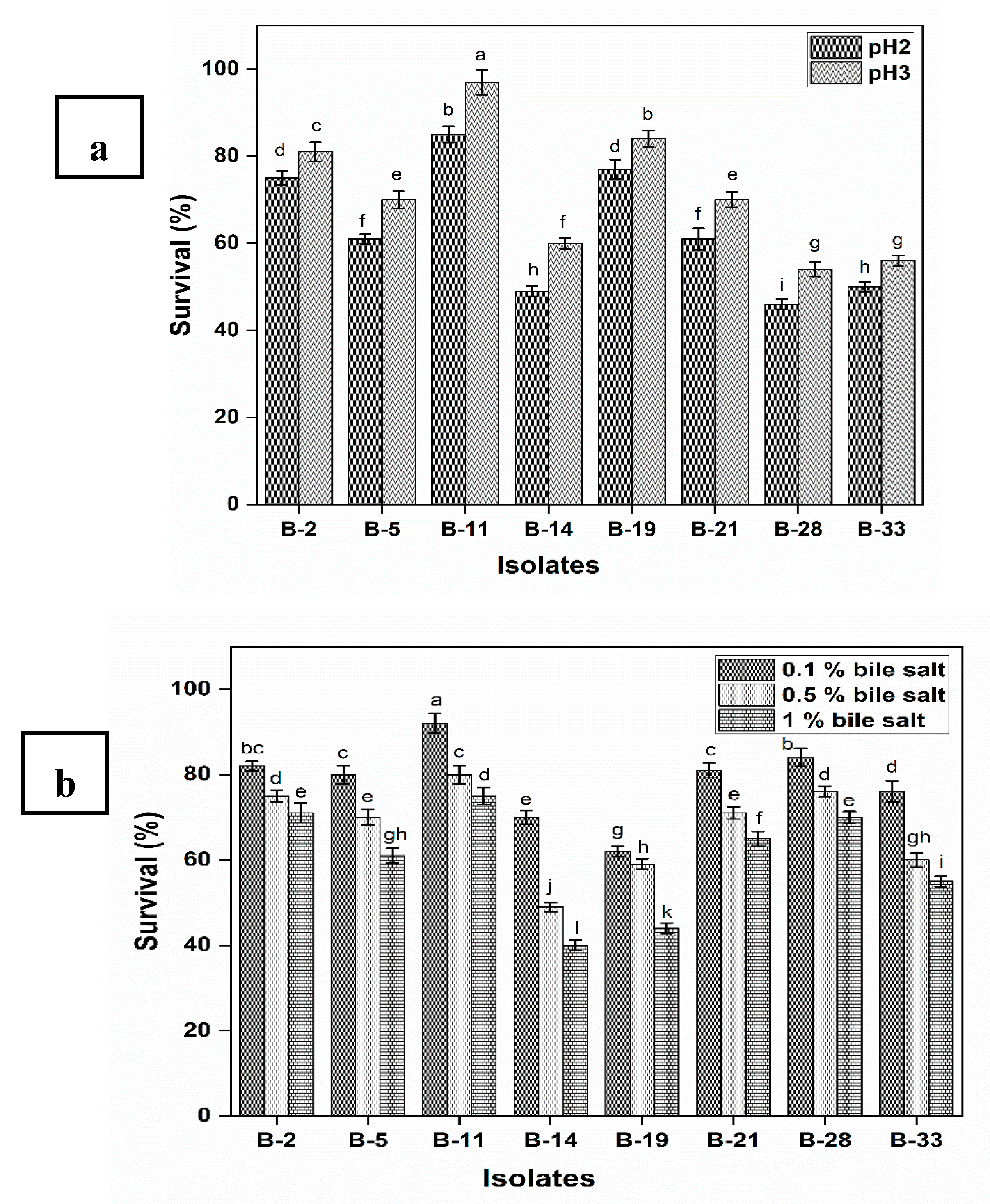
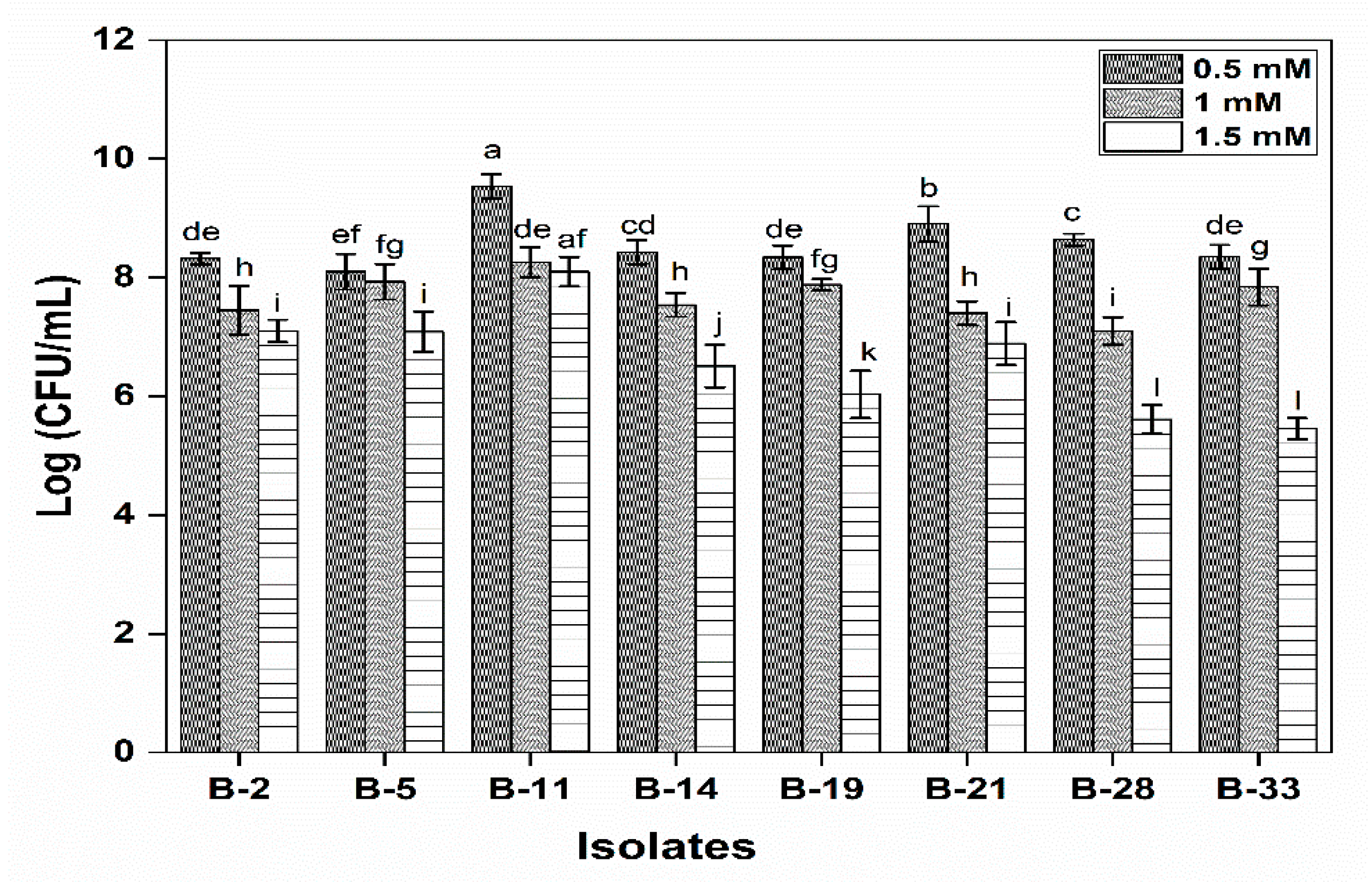
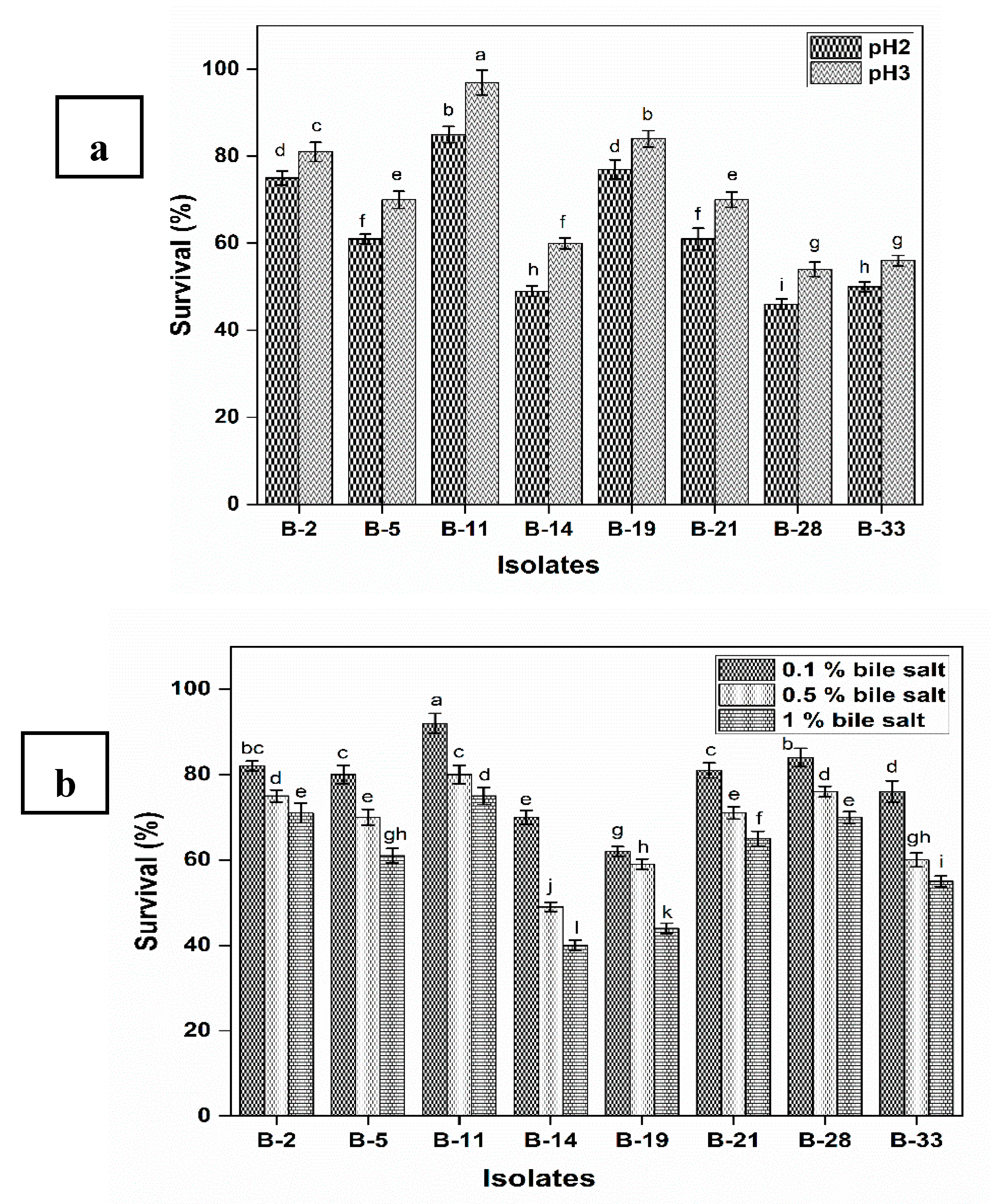
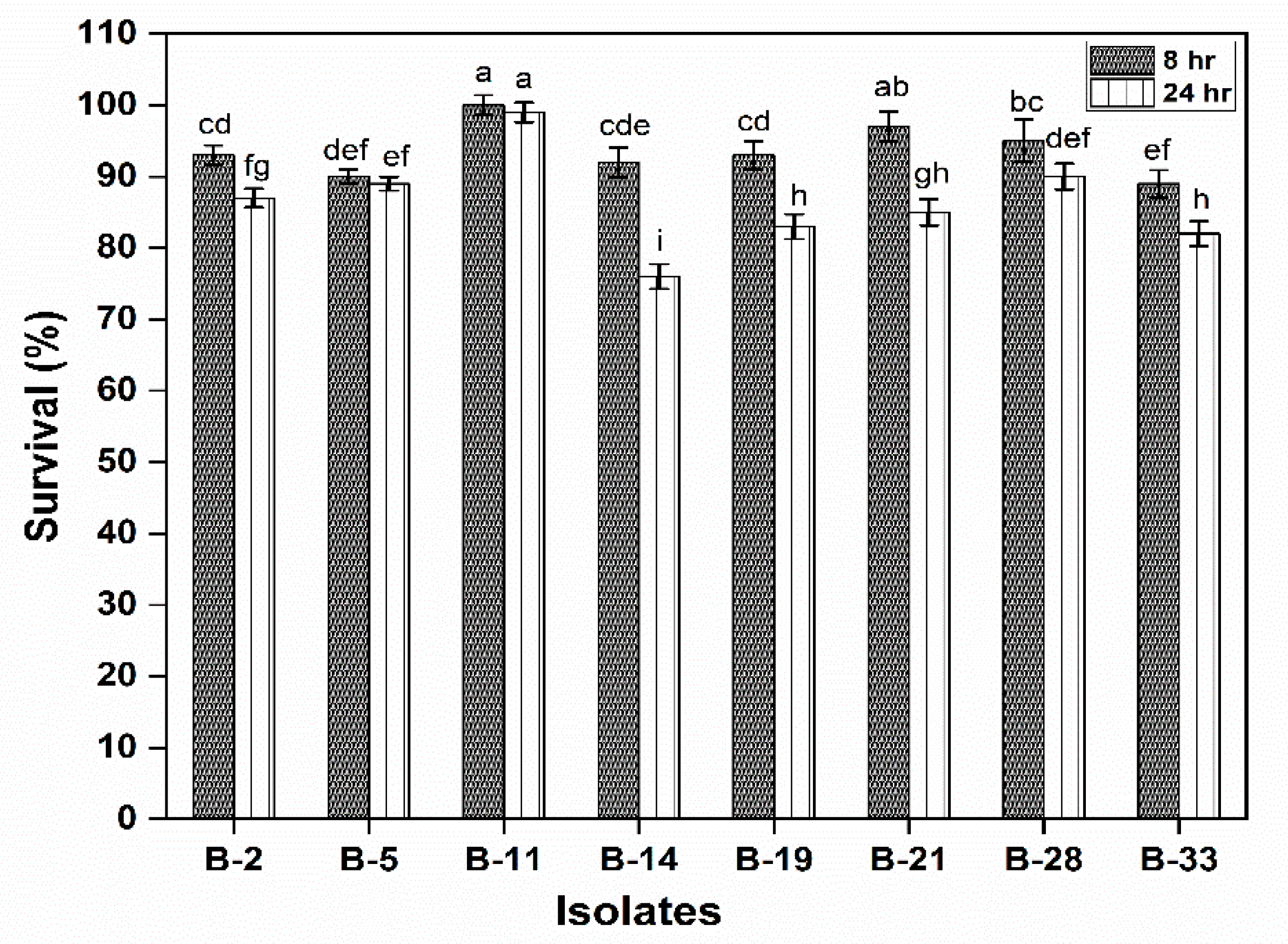
2.4.1. Survival under Gastrointestinal Conditions and Phenol Tolerance
2.4.2. Production of Exopolysaccharides (EPS)
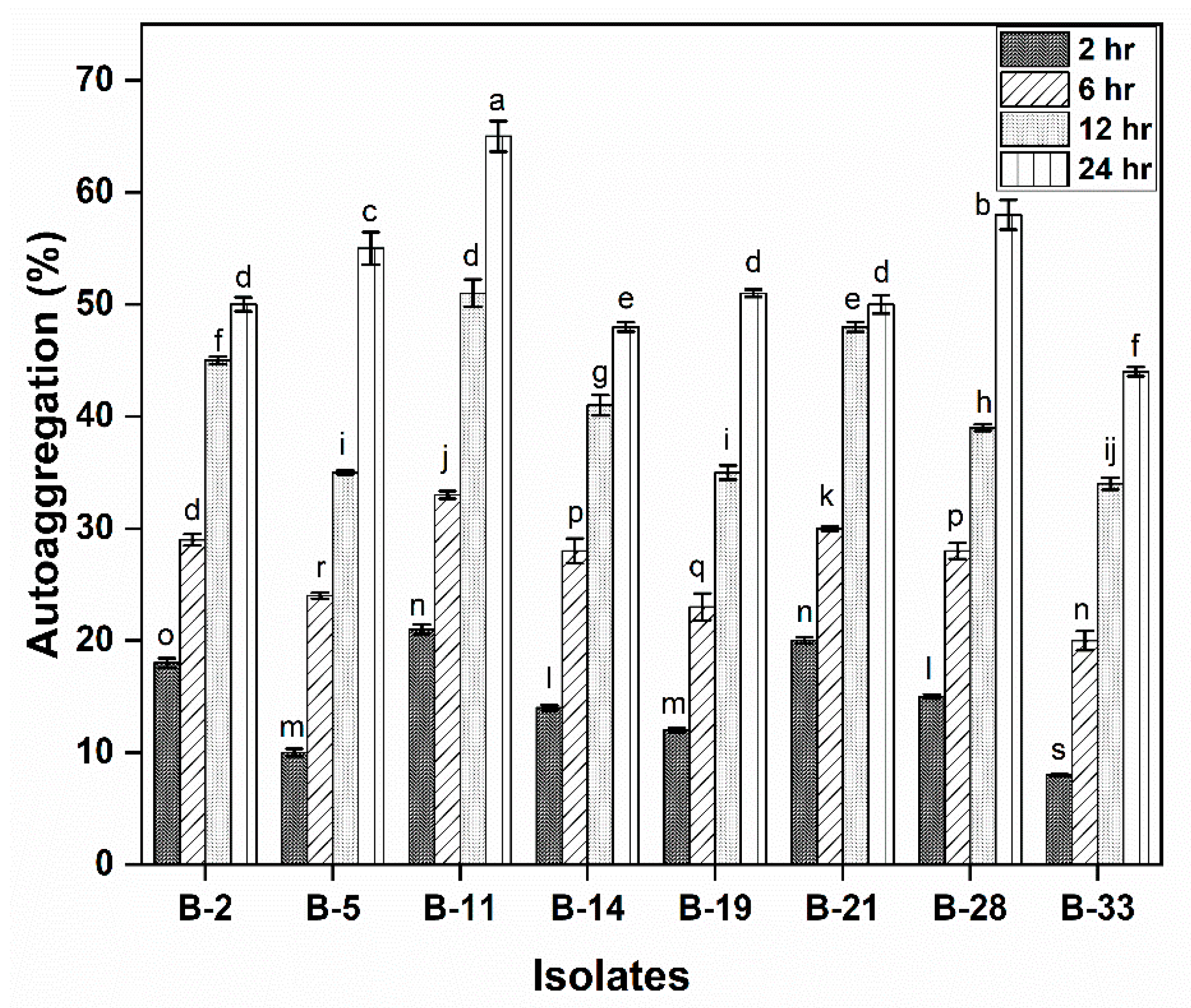
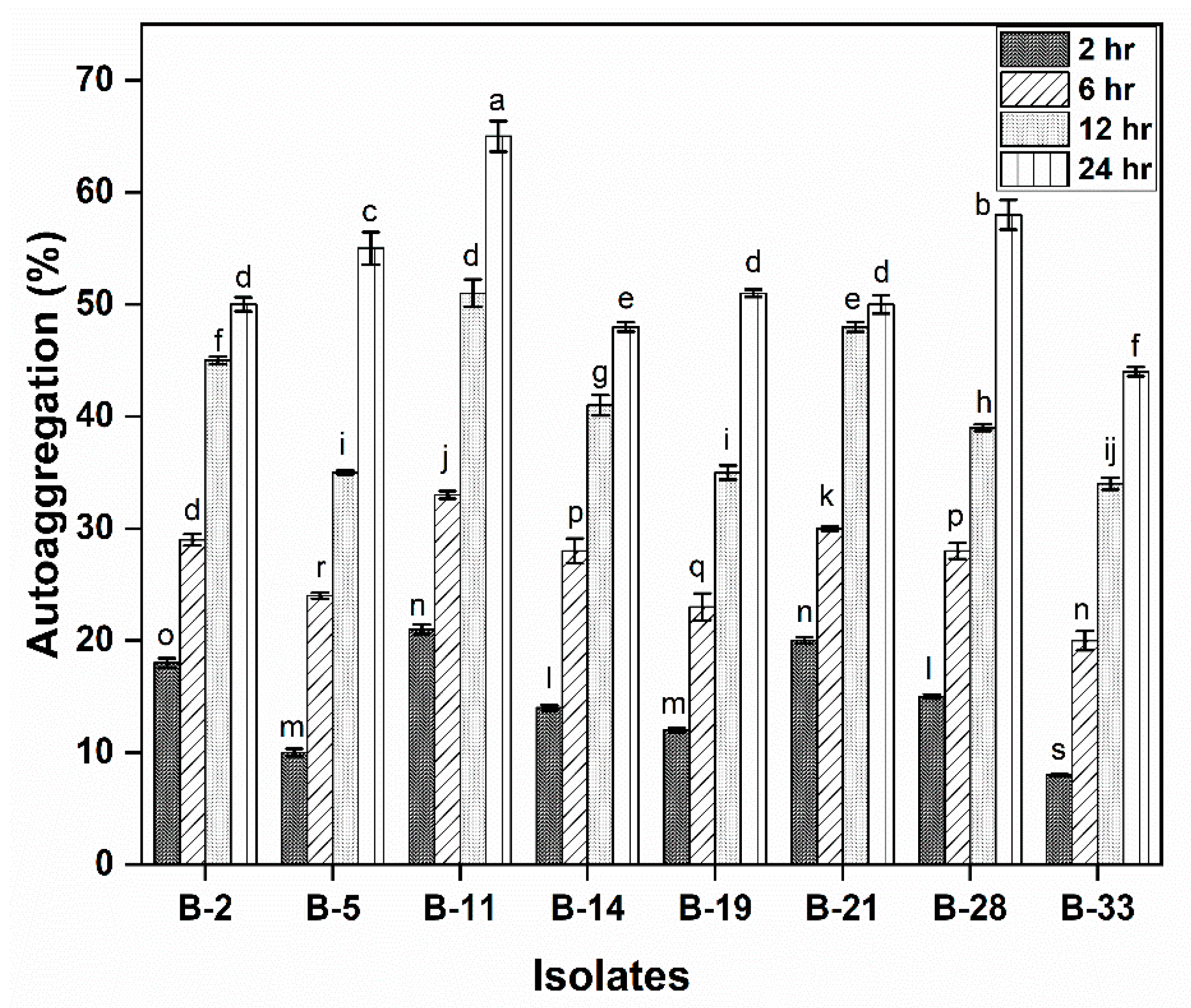
2.4.3. Auto-Aggregation Assay
2.4.4. Cell Surface Hydrophobicity
2.4.5. DPPH Free Radical Scavenging Activity
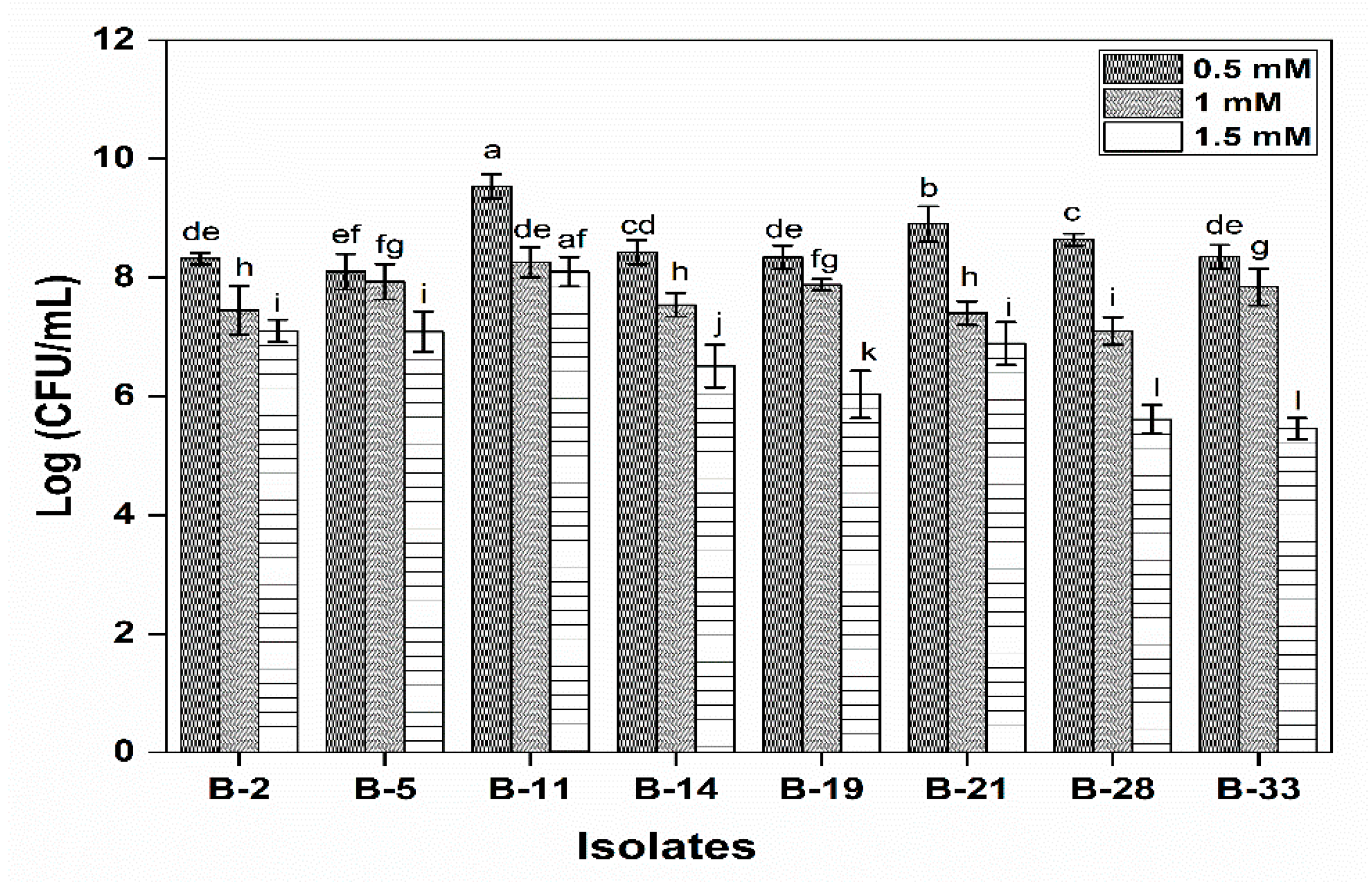
2.4.6. Resistance to Hydrogen Peroxide
2.4.7. Depletion of Sodium Nitrite
2.4.8. Antibacterial Activity
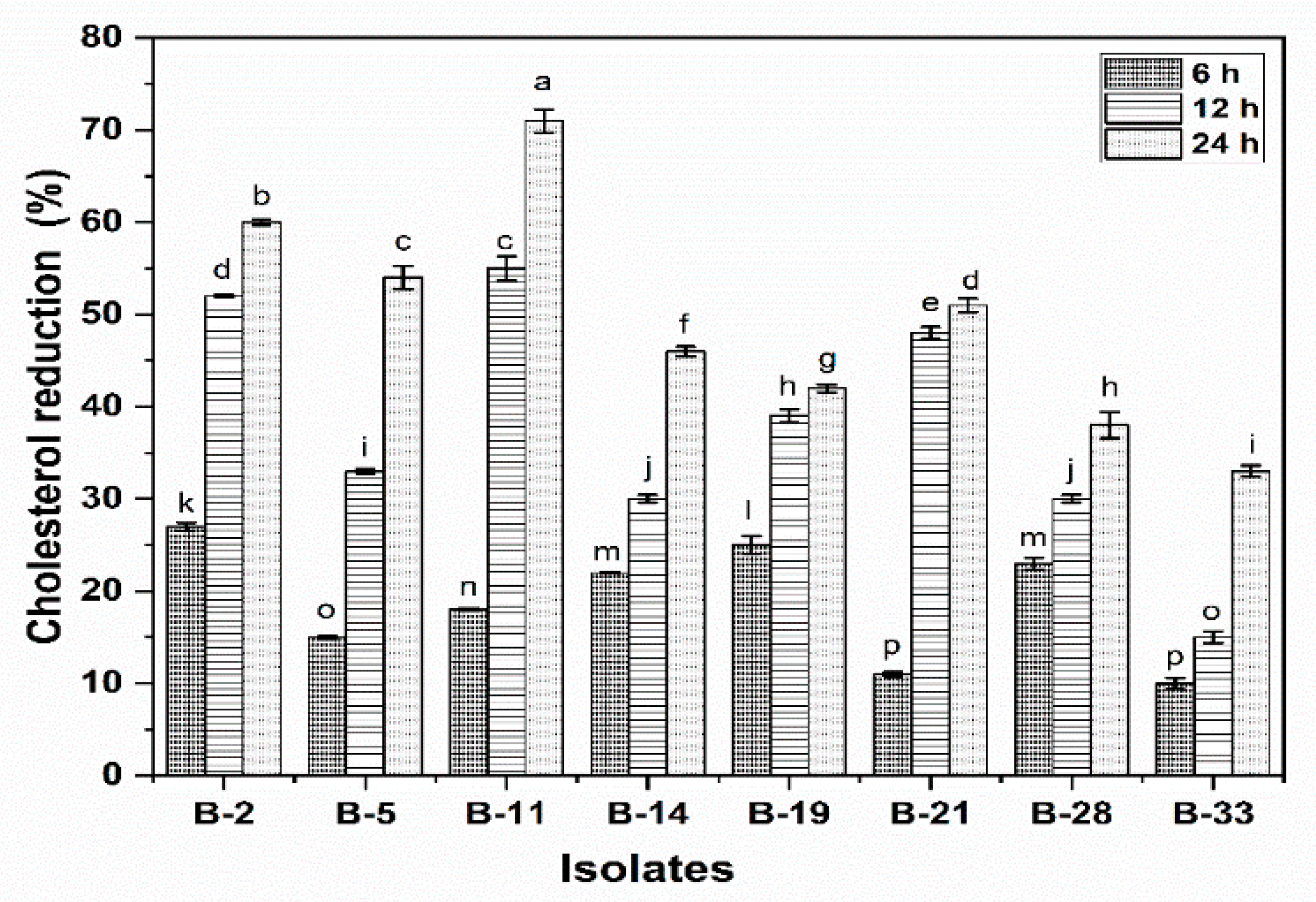
2.4.9. Cholesterol Reduction Assay
2.5. Safety Evaluation of Isolates
2.5.1. Antibiotic Susceptibility
2.5.2. Hemolytic Activity
2.5.3. Cytotoxicity
2.6. Molecular Identification of Isolate (B-11)
2.7. Attachment of B. Longum B-11 to Rats Intestine; In Vivo Studies
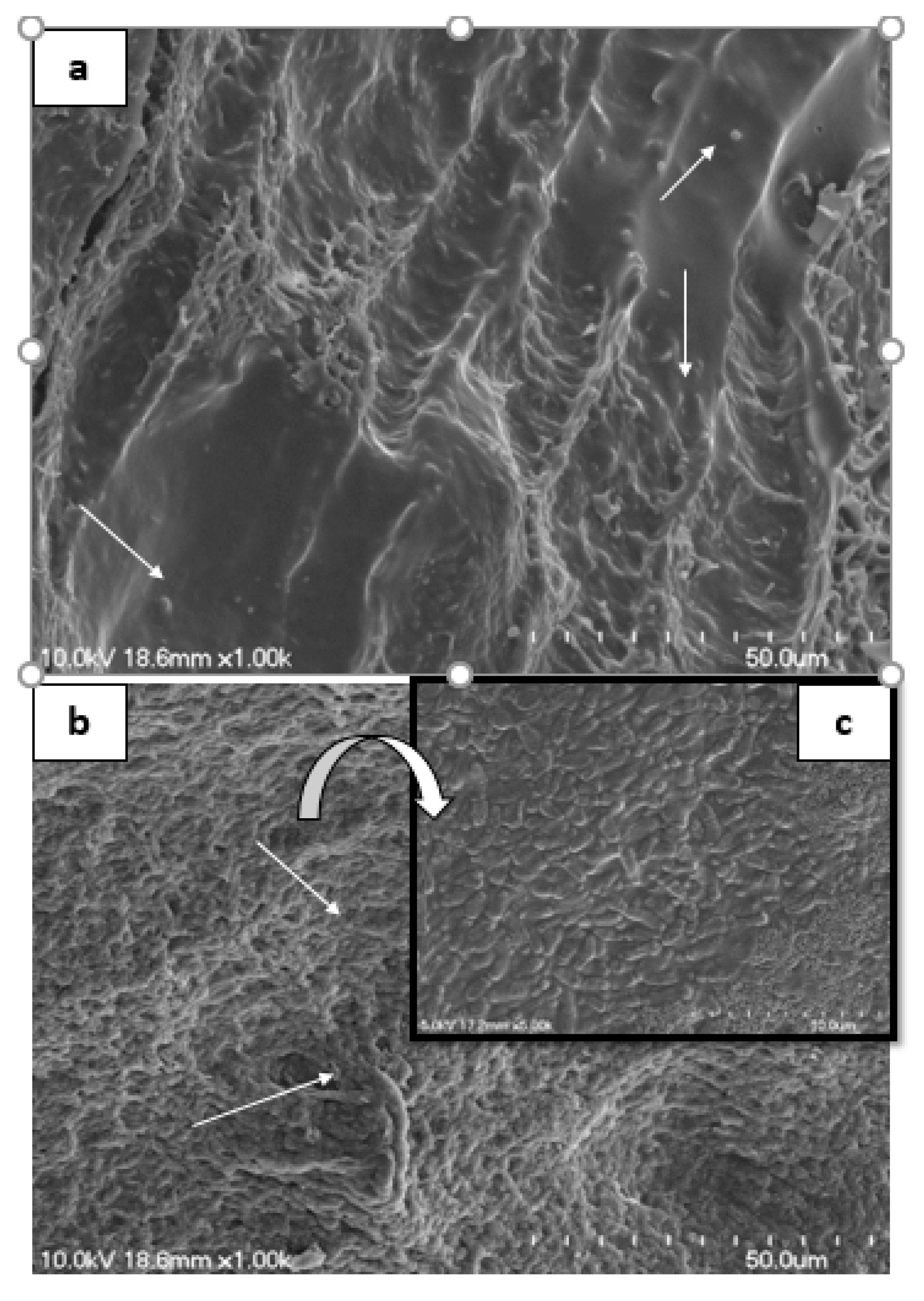
2.8. Scanning Electron Microscopy of Rats Intestine
2.9. Statistical Analysis
2.10. Ethical Approval
3. Results
3.1. Isolation and Identification of Presumptive Bifidobacterium Isolates
3.2. In Vitro Screening of Probiotic Potential of Isolates
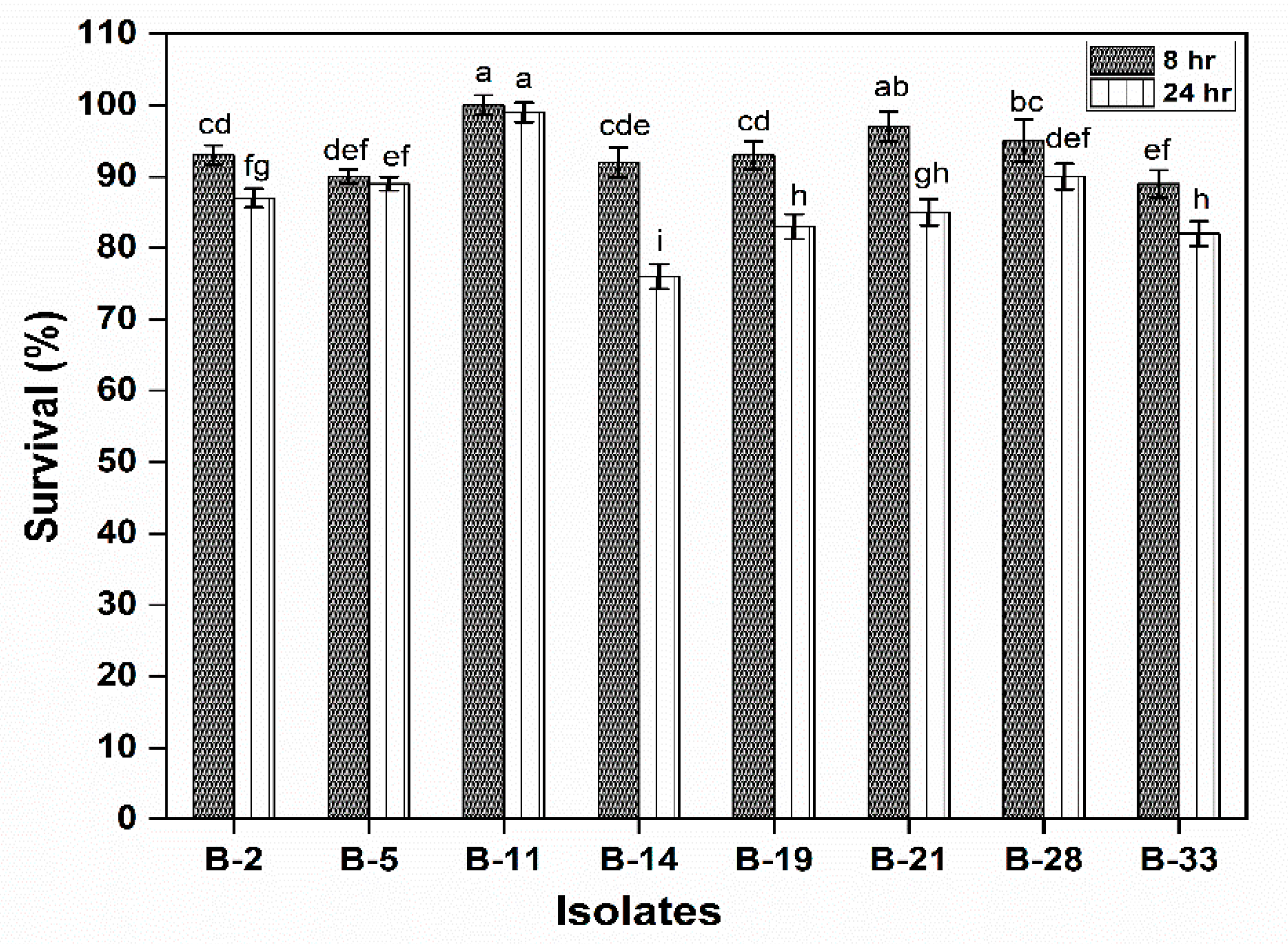
3.2.1. Survival under Gastrointestinal Conditions and Phenol Tolerance
3.2.2. Production of Exopolysaccharides (EPS)
3.2.3. Auto-Aggregation Assay
3.2.4. Cell Surface Hydrophobicity
3.2.5. DPPH Free Radical Scavenging Activity
3.2.6. Resistance to Hydrogen Peroxide
3.2.7. Depletion of Sodium Nitrite
3.2.8. Antibacterial Activity
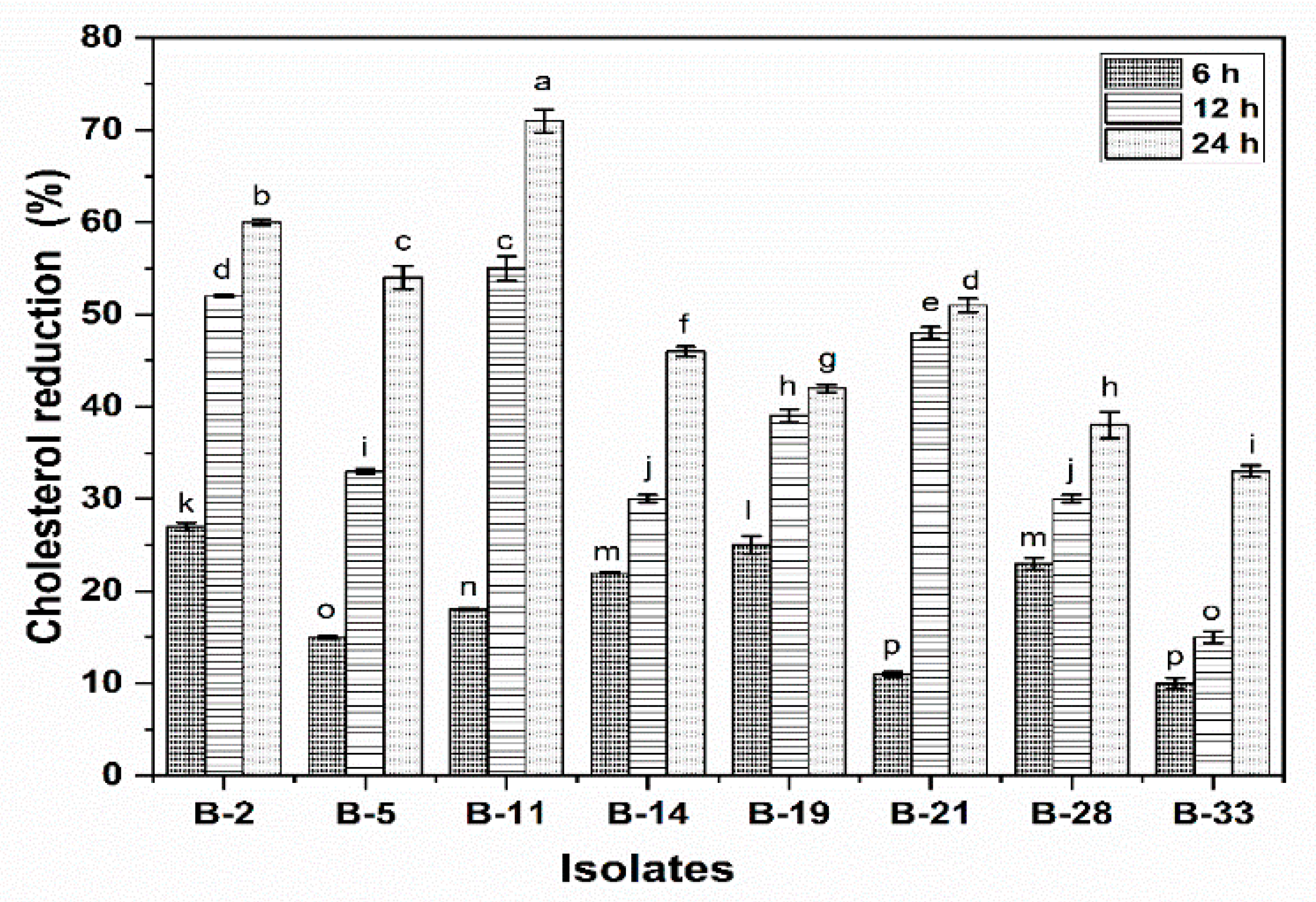
3.2.9. Cholesterol Reduction Assay
3.3. Safety Evaluation of Isolates
3.3.1. Antibiotics Susceptibility
3.3.2. Hemolytic Activity
3.3.3. Cytotoxicity
3.4. Molecular Identification of Isolate (B-11)
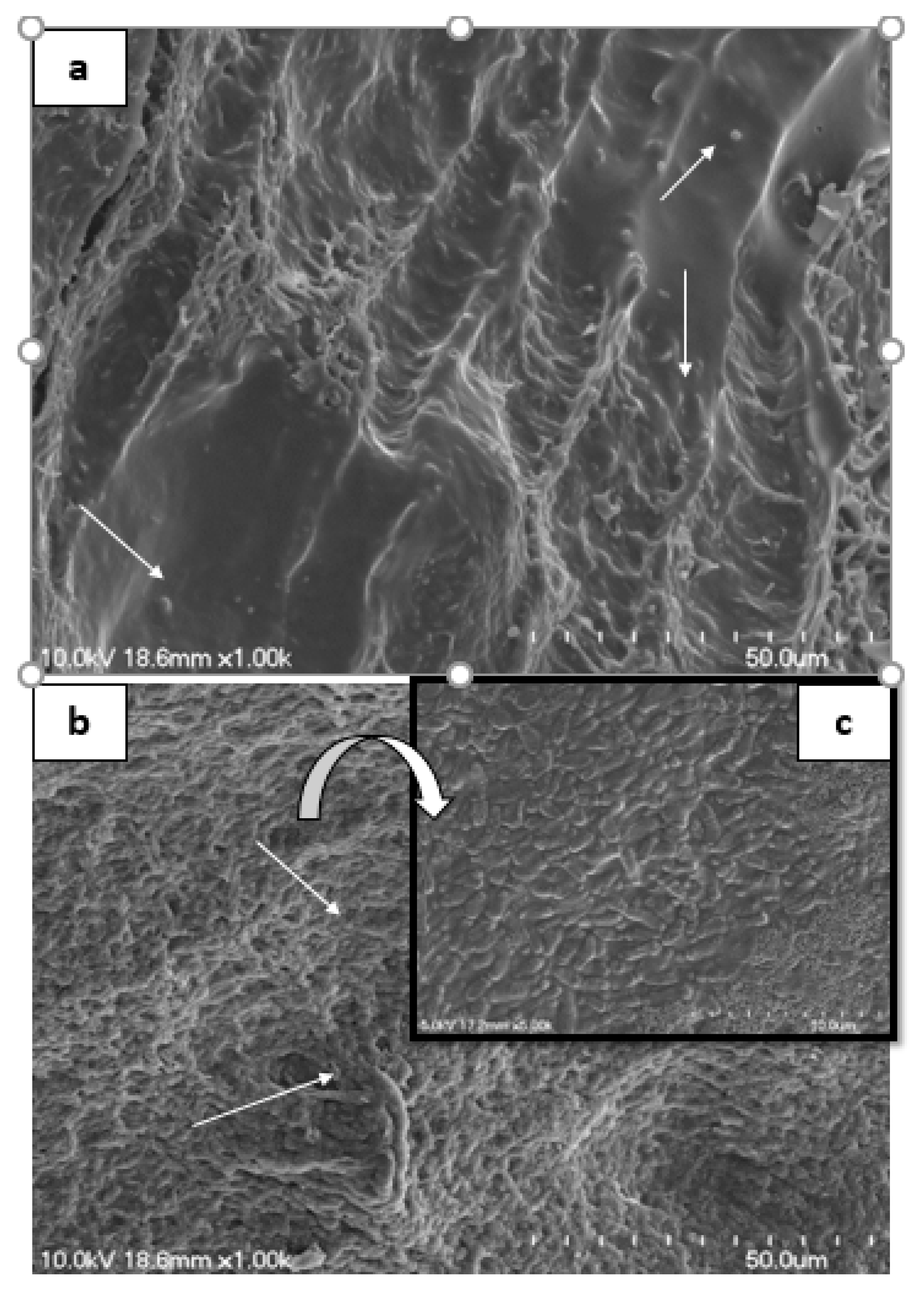
3.5. Scanning Electron Microscopy of Rat Intestine
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- García-Ruiz, A.; de Llano, D.G.; Esteban-Fernández, A.; Requena, T.; Bartolomé, B.; Moreno-Arribas, M.V. Assessment of probiotic properties in lactic acid bacteria isolated from wine. Food Microbiol. 2014, 44, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, L.C.S.; Casarotti, S.N.; Todorov, S.D.; Penna, A.L.B. Probiotic potential and safety of enterococci strains. Annals Microbiol. 2019, 69, 241–252. [Google Scholar] [CrossRef]
- Moreno, I.; Marasca, E.T.G.; de Sá, P.B.Z.R.; de Souza Moitinho, J.; Marquezini, M.G.; Alves, M.R.C.; Bromberg, R. Evaluation of probiotic potential of bacteriocinogenic lactic acid bacteria strains isolated from meat products. Probiotics Antimicrob. Proteins 2018, 10, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Lugli, G.A.; Duranti, S.; Turroni, F.; Bottacini, F.; Mangifesta, M.; Sanchez, B.; Viappiani, A.; Mancabelli, L.; Taminiau, B. Genomic encyclopedia of type strains of the genus Bifidobacterium. Appl. Environ. Microbiol. 2014, 80, 6290–6302. [Google Scholar] [CrossRef] [Green Version]
- Verso, L.L.; Lessard, M.; Talbot, G.; Fernandez, B.; Fliss, I. Isolation and selection of potential probiotic bacteria from the pig gastrointestinal tract. Probiotics Antimicrob. Proteins 2018, 10, 299–312. [Google Scholar] [CrossRef]
- Curto, A.L.; Pitino, I.; Mandalari, G.; Dainty, J.R.; Faulks, R.M.; Wickham, M.S.J. Survival of probiotic lactobacilli in the upper gastrointestinal tract using an in vitro gastric model of digestion. Food Microbiol. 2011, 28, 1359–1366. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, X.; Li, S.; Li, C.; Li, D.; Yang, Z. Evaluation of probiotic properties of Lactobacillus plantarum strains isolated from Chinese sauerkraut. World J. Microbiol. Biotechnol. 2013, 29, 489–498. [Google Scholar] [CrossRef]
- Riaz, T.; Iqbal, M.W.; Saeed, M.; Yasmin, I.; Hassanin, H.A.; Mahmood, S.; Rehman, A. In vitro survival of Bifidobacterium bifidum microencapsulated in zein-coated alginate hydrogel microbeads. J. Microencap. 2019, 36, 192–203. [Google Scholar] [CrossRef]
- Ayhan, D.K.; Temiz, A.; Sana, F.A.; Gümüşderelioğlu, M. Surface properties and exopolysaccharide production of surface-associated microorganisms isolated from a dairy plant. Ann. Microbiol. 2019, 69, 895–907. [Google Scholar] [CrossRef]
- Ren, D.; Li, C.; Qin, Y.; Yin, R.; Du, S.; Ye, F.; Liu, C.; Liu, H.; Wang, M.; Li, Y.; et al. In vitro evaluation of the probiotic and functional potential of Lactobacillus strains isolated from fermented food and human intestine. Anaerobe 2014, 30, 1–10. [Google Scholar] [CrossRef]
- Yateem, A.; Balba, M.; Al-Surrayai, T.; Al-Mutairi, B.; Al-Daher, R. Isolation of lactic acid bacteria with probiotic potential from camel milk. Inter. J. Dairy Sci. 2008, 3, 194–199. [Google Scholar] [CrossRef]
- Ashmaig, A.; Hasan, A.; El Gaali, E. Identification of lactic acid bacteria isolated from traditional Sudanese fermented camels milk (Gariss). Afri. J. Microbiol. Res. 2009, 3, 451–457. [Google Scholar]
- Amina, Z.; Noureddine, S.; Venkatesan, A.; Perumal, V.; Hichem, B.; Asma, Z.; Yamina, M.; Miloud, H.; Mebrouk, K. Characterization and potential probiotic attributes of Lactobacillus plantarum DU10 isolated from Algerian raw camel milk. Biotechnology 2014, 13, 282–288. [Google Scholar]
- Carballo, J. Potential technological interest of indigenous lactic acid bacteria from Algerian camel milk. Ital. J. Food Sci. 2016, 28, 598–611. [Google Scholar]
- Biratu, K.; Seifu, E. Chemical composition and microbiological quality of Dhanaan: Traditional fermented camel milk produced in eastern Ethiopia. Inter. Food Res. J. 2016, 23, 2223–2228. [Google Scholar]
- Fguiri, I.; Ziadi, M.; Atigui, M.; Ayeb, N.; Arroum, S.; Assadi, M.; Khorchani, T. Isolation and characterisation of lactic acid bacteria strains from raw camel milk for potential use in the production of fermented Tunisian dairy products. Inter. J. Dairy Technol. 2016, 69, 103–113. [Google Scholar] [CrossRef]
- Mahmoudi, I.; Moussa, O.B.; Khaldi, T.E.M.; Kebouchi, M.; Soligot, C.; Le Roux, Y.; Hassouna, M. Functional in vitro screening of Lactobacillus strains isolated from Tunisian camel raw milk toward their selection as probiotic. Small Rumin. Res. 2016, 137, 91–98. [Google Scholar] [CrossRef]
- Soleymanzadeh, N.; Mirdamadi, S.; Kianirad, M. Antioxidant activity of camel and bovine milk fermented by lactic acid bacteria isolated from traditional fermented camel milk (Chal). Dairy Sci. Technol. 2016, 96, 443–457. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhou, J.; Xia, X.; Zhao, Y.; Shao, W. Probiotic potential of Lactobacillus paracasei FM-LP-4 isolated from Xinjiang camel milk yoghurt. Inter. Dairy J. 2016, 62, 28–34. [Google Scholar] [CrossRef]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyash, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. LWT-Food Sci. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Kuo, J.; Yang, Y.-T.; Lu, M.-C.; Wong, T.-Y.; Sung, P.-J.; Huang, Y.-S. Antimicrobial activity and diversity of bacteria associated with Taiwanese marine sponge Theonella swinhoei. Ann. Microbiol. 2019, 69, 253–265. [Google Scholar] [CrossRef]
- Gao, Y.; Li, D. Screening of lactic acid bacteria with cholesterol-lowering and triglyceride-lowering activity in vitro and evaluation of probiotic function. Ann. Microbiol. 2018, 68, 537–545. [Google Scholar] [CrossRef]
- Sybesma, W.; Kort, R.; Lee, Y.-K. Locally sourced probiotics, the next opportunity for developing countries? Trends Biotechnol. 2015, 33, 197–200. [Google Scholar] [CrossRef]
- Yadav, R.; Puniya, A.K.; Shukla, P. Probiotic properties of Lactobacillus plantarum RYPR1 from an indigenous fermented beverage Raabadi. Front. Microbiol. 2016, 7, 1683. [Google Scholar] [CrossRef] [Green Version]
- Yasmin, I.; Saeed, M.; Pasha, I.; Zia, M.A. Development of Whey Protein Concentrate-Pectin-Alginate Based Delivery System to Improve Survival of B. longum BL-05 in Simulated Gastrointestinal Conditions. Probiotics Antimicrob. Proteins 2019, 11, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.F.; Tseng, K.C.; Chiang, S.S.; Lee, B.H.; Hsu, W.H.; Pan, T.M. Immunomodulatory and antioxidant potential of Lactobacillus exopolysaccharides. J. Sci. Food Agric. 2011, 91, 2284–2291. [Google Scholar] [CrossRef]
- Nikolic, M.; López, P.; Strahinic, I.; Suárez, A.; Kojic, M.; Fernández-García, M.; Topisirovic, L.; Golic, N.; Ruas-Madiedo, P. Characterisation of the exopolysaccharide (EPS)-producing Lactobacillus paraplantarum BGCG11 and its non-EPS producing derivative strains as potential probiotics. Inter. J. Food Microbiol. 2012, 158, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Jeong, H.; Lee, H.; Ahn, J. Assessment of cell surface properties and adhesion potential of selected probiotic strains. Lett. App. Microbiol. 2009, 49, 434–442. [Google Scholar] [CrossRef]
- Kotzamanidis, C.; Kourelis, A.; Litopoulou-Tzanetaki, E.; Tzanetakis, N.; Yiangou, M. Evaluation of adhesion capacity, cell surface traits and immunomodulatory activity of presumptive probiotic Lactobacillus strains. Inter. J. Food Microbiol. 2010, 140, 154–163. [Google Scholar] [CrossRef]
- Chen, F.A.; Wu, A.B.; Shieh, P.; Kuo, D.H.; Hsieh, C.Y. Evaluation of the antioxidant activity of Ruellia tuberosa. Food Chem. 2006, 94, 14–18. [Google Scholar] [CrossRef]
- Oberg, T.; Steele, J.; Ingham, S.; Smeianov, V.; Briczinski, E.; Abdalla, A.; Broadbent, J.R. Intrinsic and inducible resistance to hydrogen peroxide in Bifidobacterium species. J. Indus. Microbiol. Biotechnol. 2011, 38, 1947–1953. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Y.; Liu, F.J.; Li, L.H.; Yang, X.Q.; Deng, J.C.; Chen, S.J. Isolation and identification of nitrite-degrading lactic acid bacteria from salted fish. Adv. Mater. Res. 2012, 393, 828–834. [Google Scholar] [CrossRef]
- Yan, P.M.; Xue, W.T.; Tan, S.S.; Zhang, H.; Chang, X.H. Effect of inoculating lactic acid bacteria starter cultures on the nitrite concentration of fermenting Chinese paocai. Food Control 2008, 19, 50–55. [Google Scholar] [CrossRef]
- Rudel, L.L.; Morris, M. Determination of cholesterol using o-phthalaldehyde. J. Lipid Res. 1973, 14, 364–366. [Google Scholar] [PubMed]
- Liong, M.; Shah, N. Bile salt deconjugation ability, bile salt hydrolase activity and cholesterol co-precipitation ability of lactobacilli strains. Int. Dairy J. 2005, 15, 391–398. [Google Scholar] [CrossRef]
- Vijayakumar, M.; Ilavenil, S.; Kim, D.H.; Arasu, M.V.; Priya, K.; Choi, K.C. In-vitro assessment of the probiotic potential of Lactobacillus plantarum KCC-24 isolated from Italian rye-grass (Lolium multiflorum) forage. Anaerobe 2015, 32, 90–97. [Google Scholar] [CrossRef]
- Mangia, N.P.; Saliba, L.; Deiana, P. Functional and safety characterization of autochthonous Lactobacillus paracasei FS103 isolated from sheep cheese and its survival in sheep and cow fermented milks during cold storage. Ann. Microbiol. 2019, 69, 161–170. [Google Scholar] [CrossRef]
- Mohanty, D.; Panda, S.; Kumar, S.; Ray, P. In vitro evaluation of adherence and anti-infective property of probiotic Lactobacillus plantarum DM 69 against Salmonella enterica. Microbial. Pathog. 2019, 126, 212–217. [Google Scholar] [CrossRef]
- Martinez, I.; Stegen, J.C.; Maldonado-Gomez, M.X.; Eren, A.M.; Siba, P.M.; Greenhill, A.R.; Walter, J. The gut microbiota of rural papua new guineans: Composition, diversity patterns, and ecological processes. Cell Rep. 2015, 11, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, J.; Dubey, R.; Sengar, G.; Dheeman, S. Evaluation of probiotic potential and safety assessment of Lactobacillus pentosus MMP4 isolated from mare’s lactation. Probiotics Antimicrob. Proteins 2019, 11, 403–412. [Google Scholar] [CrossRef]
- Khan, W.A.; Butt, M.S.; Pasha, I.; Jamil, A. Microencapsulation of vitamin D in protein matrices: In vitro release and storage stability. J. Food Meas. Charact. 2020, 1–11. [Google Scholar] [CrossRef]
- Izquierdo, E.; Medina, M.; Ennahar, S.; Marchioni, E.; Sanz, Y. Resistance to simulated gastrointestinal conditions and adhesion to mucus as probiotic criteria for Bifidobacterium longum strains. Curr. Microbiol. 2008, 56, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Shigwedha, N.; Mwandemele, O.D. Use of Dacid-, Dbile-, zacid-, and zbile-Values in Evaluating Bifidobacteria with Regard to Stomach pH and Bile Salt Sensitivity. J. Food Sci. 2010, 75. [Google Scholar] [CrossRef]
- Casarotti, S.N.; Penna, A.L.B. Acidification profile, probiotic in vitro gastrointestinal tolerance and viability in fermented milk with fruit flours. Inter. Dairy J. 2015, 41, 1–6. [Google Scholar] [CrossRef]
- Larsen, N.; Cahú, T.B.; Saad, S.M.I.; Blennow, A.; Jespersen, L. The effect of pectins on survival of probiotic Lactobacillus spp. in gastrointestinal juices is related to their structure and physical properties. Food Microbiol. 2018, 74, 11–20. [Google Scholar] [CrossRef]
- Sanchez, B.; Champomier-Verges, M.C.; Collado Mdel, C.; Anglade, P.; Baraige, F.; Sanz, Y.; de los Reyes-Gavilan, C.G.; Margolles, A.; Zagorec, M. Low-pH adaptation and the acid tolerance response of Bifidobacterium longum biotype longum. Appl. Environ. Microbiol. 2007, 73, 6450–6459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haros, M.; Carlsson, N.G.; Almgren, A.; Larsson-Alminger, M.; Sandberg, A.S.; Andlid, T. Phytate degradation by human gut isolated Bifidobacterium pseudocatenulatum ATCC27919 and its probiotic potential. Inter. J. Food Microbiol. 2009, 135, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Haghshenas, B.; Abdullah, N.; Nami, Y.; Radiah, D.; Rosli, R.; Khosroushahi, A.Y. Different effects of two newly-isolated probiotic Lactobacillus plantarum 15HN and Lactococcus lactis subsp. Lactis 44Lac strains from traditional dairy products on cancer cell lines. Anaerobe 2014, 30, 51–59. [Google Scholar] [CrossRef]
- Bengoa, A.A.; Llamas, M.G.; Iraporda, C.; Dueñas, M.T.; Abraham, A.G.; Garrote, G.L. Impact of growth temperature on exopolysaccharide production and probiotic properties of Lactobacillus paracasei strains isolated from kefir grains. Food Microbiol. 2018, 69, 212–218. [Google Scholar] [CrossRef]
- Prasanna, P.; Grandison, A.S.; Charalampopoulos, D. Screening human intestinal Bifidobacterium strains for growth, acidification, EPS production and viscosity potential in low-fat milk. Inter. Dairy J. 2012, 23, 36–44. [Google Scholar] [CrossRef]
- Alp, G.; Aslim, B. Relationship between the resistance to bile salts and low pH with exopolysaccharide (EPS) production of Bifidobacterium spp. isolated from infants feces and breast milk. Anaerobe 2010, 16, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Kim, W.-S.; Kumura, H.; Shimazaki, K.-i. Autoaggregation and surface hydrophobicity of bifidobacteria. World J. Microbiol. Biotechnol. 2008, 24, 1593–1598. [Google Scholar] [CrossRef]
- Russell, D.A.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Metabolic activities and probiotic potential of bifidobacteria. Inter. J. Food Microbiol. 2011, 149, 88–105. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Shang, N.; Li, P. In vitro and in vivo antioxidant activity of Bifidobacterium animalis 01 isolated from centenarians. Cur. Microbiol. 2011, 62, 1097–1103. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Yu, R.-C.; Chou, C.-C. Antioxidative activities of soymilk fermented with lactic acid bacteria and bifidobacteria. Food Microbiol. 2006, 23, 128–135. [Google Scholar] [CrossRef]
- Soubra, L.; Sarkis, D.; Hilan, C.; Verger, P. Dietary exposure of children and teenagers to benzoates, sulphites, butylhydroxyanisol (BHA) and butylhydroxytoluen (BHT) in Beirut (Lebanon). Regul. Toxicol. Pharm. 2007, 47, 68–77. [Google Scholar] [CrossRef]
- Collado, M.; Gonzalez, A.; Gonzalez, R.; Hernandez, M.; Ferrus, M.; Sanz, Y. Antimicrobial peptides are among the antagonistic metabolites produced by Bifidobacterium against Helicobacter pylori. Inter. J. Anti. Agents 2005, 25, 385–391. [Google Scholar] [CrossRef]
- El-Gawad, I.A.A.; El-Sayed, E.; Hafez, S.; El-Zeini, H.; Saleh, F. The hypocholesterolaemic effect of milk yoghurt and soy-yoghurt containing bifidobacteria in rats fed on a cholesterol-enriched diet. Int. Dairy J. 2005, 15, 37–44. [Google Scholar] [CrossRef]
- Xiao, J.; Kondo, S.; Takahashi, N.; Miyaji, K.; Oshida, K.; Hiramatsu, A.; Iwatsuki, K.; Kokubo, S.; Hosono, A. Effects of milk products fermented by Bifidobacterium longum on blood lipids in rats and healthy adult male volunteers. J. Dairy Sci. 2003, 86, 2452–2461. [Google Scholar] [CrossRef]
- Zhou, J.; Pillidge, C.; Gopal, P.; Gill, H. Antibiotic susceptibility profiles of new probiotic Lactobacillus and Bifidobacterium strains. Inter. J. Food Microbiol. 2005, 98, 211–217. [Google Scholar] [CrossRef]
- Temmerman, R.; Pot, B.; Huys, G.; Swings, J. Identification and antibiotic susceptibility of bacterial isolates from probiotic products. Int. J. Food Microbiol. 2003, 81, 1–10. [Google Scholar] [CrossRef]
- Oh, Y.J.; Jung, D.S. Evaluation of probiotic properties of Lactobacillus and Pediococcus strains isolated from Omegisool, a traditionally fermented millet alcoholic beverage in Korea. LWT-Food Sci. Technol. 2015, 63, 437–444. [Google Scholar] [CrossRef]
- Wang, J.; Yang, K.; Liu, M.; Zhang, J.; Wei, X.; Fan, M. Screening for potential probiotic from spontaneously fermented non-dairy foods based on in vitro probiotic and safety properties. Ann. Microbiol. 2018, 68, 803–813. [Google Scholar] [CrossRef]
- Bujnakova, D.; Strakova, E. Safety, probiotic and technological properties of Lactobacilli isolated from unpasteurised ovine and caprine cheeses. Ann. Microbiol. 2017, 67, 813–826. [Google Scholar] [CrossRef]
- Nami, Y.; Haghshenas, B.; Bakhshayesh, R.V.; Jalaly, H.M.; Lotfi, H.; Eslami, S.; Hejazi, M.A. Novel autochthonous lactobacilli with probiotic aptitudes as a main starter culture for probiotic fermented milk. LWT-Food Sci. Technol. 2018, 98, 85–93. [Google Scholar] [CrossRef]
- Tarrah, A.; da Silva Duarte, V.; de Castilhos, J.; Pakroo, S.; Junior, W.J.F.L.; Luchese, R.H.; Guerra, A.F.; Rossi, R.C.; Ziegler, D.R.; Corich, V. Probiotic potential and biofilm inhibitory activity of Lactobacillus casei group strains isolated from infant feces. J. Funct. Foods. 2019, 54, 489–497. [Google Scholar] [CrossRef]
- Chiu, Y.-H.; Lu, Y.-C.; Ou, C.-C.; Lin, S.-L.; Tsai, C.-C.; Huang, C.-T.; Lin, M.-Y. Lactobacillus plantarum MYL26 induces endotoxin tolerance phenotype in Caco-2 cells. BMC Microbiol. 2013, 13, 190. [Google Scholar] [CrossRef] [Green Version]
- Awaisheh, S.; Obeidat, M.; Al-Tamimi, H.; Assaf, A.; EL-Qudah, J.; Rahahleh, R. In vitro cytotoxic activity of probiotic bacterial cell extracts against Caco-2 and HRT-18 colorectal cancer cells. Milk Sci. Inter.-Milchwiss. 2016, 69, 33–37. [Google Scholar]
- Er, S.; KOPARAL, A.T.; Kivanc, M. Cytotoxic effects of various lactic acid bacteria on Caco-2 cells. Turk. J. Biol. 2015, 39, 23–30. [Google Scholar] [CrossRef]
- Bhat, M.I.; Singh, V.K.; Sharma, D.; Kapila, S.; Kapila, R. Adherence capability and safety assessment of an indigenous probiotic strain Lactobacillus rhamnosus MTCC-5897. Microbial Pathog. 2019, 130, 120–130. [Google Scholar] [CrossRef]
- Joghataei, M.; Shahidi, F.; Pouladfar, G.; Mortazavi, S.A.; Ghaderi, A. Probiotic potential comparison of Lactobacillus strains isolated from Iranian traditional food products and human feces with standard probiotic strains. J. Sci. Food Agric. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.-K.; Hong, J.-Y.; Yi, S.-H.; Hong, S.-P.; Lee, J.-E.; Paik, H.-D. Bioactive compounds of probiotic Saccharomyces cerevisiae strains isolated from cucumber jangajji. J. Funct. Foods 2019, 58, 324–329. [Google Scholar] [CrossRef]
- Ozkan, A.D.; Han, D.; Umu, O.C.; Angun, P.; Senturk, B.; Yasa, O.; Tekinay, T. Screening and selection of novel animal probiotics isolated from bovine chyme. Ann. Microbiol. 2013, 63, 1291–1300. [Google Scholar] [CrossRef]
- Le Leu, R.K.; Hu, Y.; Brown, I.L.; Woodman, R.J.; Young, G.P. Synbiotic intervention of Bifidobacterium lactis and resistant starch protects against colorectal cancer development in rats. Carcinogenesis 2009, 31, 246–251. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tests | Isolates | |||||||
|---|---|---|---|---|---|---|---|---|
| B-2 | B-5 | B-11 | B-14 | B-19 | B-21 | B-28 | B-33 | |
| Morphology | V shaped rod | rod | Y- shaped rod | V-shaped rod | rod | rod | V-shaped rod | rod |
| Gram Staining | + | + | + | + | + | + | + | + |
| Catalase test | - | - | - | - | - | - | - | - |
| Growth at different temperature | ||||||||
| 10 °C | - | + | + | + | - | + | - | + |
| 37 °C | + | + | + | + | + | + | + | + |
| 45 °C | + | - | + | + | + | + | - | + |
| Growth at different NaCl concentration | ||||||||
| 3% | + | + | + | + | + | + | + | + |
| 5% | + | - | + | + | - | + | - | + |
| 7% | + | - | - | - | - | + | - | - |
| Growth at different pH | ||||||||
| 4.4 | + | + | + | + | + | + | + | + |
| 9.6 | - | - | - | - | - | + | - | - |
| Motility test | - | - | - | - | - | - | - | - |
| Endospore test | - | - | - | - | - | - | - | - |
| CO2 from glucose | - | - | - | - | - | - | - | - |
| Carbohydrate fermentation | ||||||||
| Glucose | + | + | + | + | + | + | + | + |
| Maltose | - | + | + | - | + | + | + | + |
| Lactose | + | + | + | + | + | - | - | + |
| Galactose | + | + | - | - | - | + | + | + |
| Raffinose | + | + | + | + | + | + | + | + |
| Sorbitol | - | + | - | - | + | + | + | + |
| Sucrose | + | - | + | + | - | - | - | + |
| Xylose | + | + | + | - | - | + | + | + |
| Fructose | + | + | + | - | + | + | + | + |
| Isolates | Phenol Tolerance (log CFU/mL) | EPS Production (mg/L) | Cell Surface Hydrophobicity (%) | DPPH Free Radical Scavenging Activity (%) | Depletion of Sodium Nitrite (%) | |
|---|---|---|---|---|---|---|
| 0 h | 24 h | |||||
| B-2 | 8.65 ± 0.14 | 8.32 ± 0.14 | 91 ± 2.41 | 56.89 ± 1.59 | 80.29 ± 3.12 | 65 ±0.08 |
| B-5 | 8.32 ± 0.43 | 8.21 ± 0.23 | 112 ± 2.53 | 74.21 ± 3.12 | 84.44 ± 2.93 | 76 ± 0.14 |
| B-11 | 8.54 ± 0.56 | 8.45 ± 0.41 | 123 ± 3.92 | 78.89 ± 2.43 | 87.72 ± 3.01 | 84 ± 0.13 |
| B-14 | 8.92 ± 0.80 | 8.12 ± 0.17 | 80 ± 2.02 | 45.21 ± 0.91 | 41.38 ± 0.42 | 54 ± 0.19 |
| B-19 | 8.46 ± 0.21 | 7.94 ± 0.21 | 56 ± 1.34 | 67.93 ± 1.25 | 70.23 ± 1.40 | 74 ± 0.13 |
| B-21 | 8.53 ± 0.11 | 8.39 ± 0.19 | 102 ± 2.10 | 49.34 ± 2.51 | 79.74 ± 2.52 | 50 ± 0.09 |
| B-28 | 7.91 ± 0.14 | 7.59 ± 0.10 | 61 ± 3.21 | 71.99 ± 2.13 | 81.19 ± 2.16 | 79 ± 0.31 |
| B-33 | 8.12 ± 0.10 | 7.98 ± 0.34 | 75 ± 2.37 | 60.77 ± 1.76 | 80.72 ± 2.34 | 69 ± 0.56 |
| Zone of Inhibition | |||
|---|---|---|---|
| Isolates | Staphylococcus aureus (mm) | Salmonella typhimurium (mm) | Escherichia coli (mm) |
| B-2 | 8 ± 0.51 | 8 ± 0.11 | 7 ± 0.26 |
| B-5 | 10 ±0.42 | 7 ± 0.09 | 10 ± 0.21 |
| B-11 | 11 ± 0.35 | 12 ±0.12 | 14 ± 0.13 |
| B-14 | 8 ± 0.14 | 10 ± 0.21 | 11 ± 0.12 |
| B-19 | 8 ± 0.21 | 9 ± 0.08 | 0 ± 0.27 |
| B-21 | 7 ± 0.28 | 10 ± 0.12 | 6 ± 0.06 |
| B-28 | 9 ± 0.17 | 0 ± 0.10 | 8 ± 0.15 |
| B-33 | 6 ± 0.06 | 6 ± 0.04 | 9 ± 0.19 |
| Antibiotics | Disc Potency (µg) | B-2 | B-5 | B-11 | B-14 | B-19 | B-21 | B-28 | B-33 |
|---|---|---|---|---|---|---|---|---|---|
| Ampicillin | 10 | S | S | S | S | S | S | S | S |
| Cephalothin | 15 | S | S | S | S | S | S | S | S |
| Chloramphenicol | 50 | S | S | S | S | S | S | S | S |
| Cloxacillin | 20 | S | S | S | S | S | S | S | S |
| Erythromycin | 10 | S | S | S | S | S | S | S | S |
| Fusidic acid | 10 | R | R | R | R | R | R | R | R |
| Gentamicin | 10 | R | R | R | R | R | R | R | R |
| Kanamycin | 30 | R | R | R | R | R | R | R | R |
| Nalidixic acid | 20 | R | R | R | R | R | R | R | R |
| Neomycin | 20 | R | R | R | S | R | R | R | R |
| Novobiocin | 30 | S | S | S | S | S | S | S | S |
| Penicillin | 10 | S | S | S | S | S | S | S | S |
| Polymyxin B | 20 | R | R | R | R | R | R | R | R |
| Rifampicin | 20 | R | R | R | R | R | S | R | R |
| Spectinomycin | 10 | S | S | S | S | S | S | S | S |
| Streptomycin | 25 | R | R | R | R | R | S | R | R |
| Tetracycline | 30 | S | S | S | S | S | S | S | S |
| Vancomycin | 30 | S | S | S | S | S | S | S | S |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasmin, I.; Saeed, M.; Khan, W.A.; Khaliq, A.; Chughtai, M.F.J.; Iqbal, R.; Tehseen, S.; Naz, S.; Liaqat, A.; Mehmood, T.; et al. In Vitro Probiotic Potential and Safety Evaluation (Hemolytic, Cytotoxic Activity) of Bifidobacterium Strains Isolated from Raw Camel Milk. Microorganisms 2020, 8, 354. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030354
Yasmin I, Saeed M, Khan WA, Khaliq A, Chughtai MFJ, Iqbal R, Tehseen S, Naz S, Liaqat A, Mehmood T, et al. In Vitro Probiotic Potential and Safety Evaluation (Hemolytic, Cytotoxic Activity) of Bifidobacterium Strains Isolated from Raw Camel Milk. Microorganisms. 2020; 8(3):354. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030354
Chicago/Turabian StyleYasmin, Iqra, Muhammad Saeed, Wahab Ali Khan, Adnan Khaliq, Muhammad Farhan Jahangir Chughtai, Rabia Iqbal, Saima Tehseen, Saima Naz, Atif Liaqat, Tariq Mehmood, and et al. 2020. "In Vitro Probiotic Potential and Safety Evaluation (Hemolytic, Cytotoxic Activity) of Bifidobacterium Strains Isolated from Raw Camel Milk" Microorganisms 8, no. 3: 354. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030354