Behavior and Mechanism of Cesium Biosorption from Aqueous Solution by Living Synechococcus PCC7002

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cyanobacteria and Growth Conditions
2.2. Cs Solution
2.3. Effect of Cs(I) on Synechococcus PCC7002 Growth
2.4. EPS Extraction and Chemical Analysis
2.5. Adsorption Experiments
2.6. Surface Analysis
2.7. Transmission Electron Microscopy (TEM), Scanning Electron Microscopy (SEM), and Energy-Dispersive X-ray Spectroscopy (EDX) Analyses
2.8. Statistical Analysis
3. Results
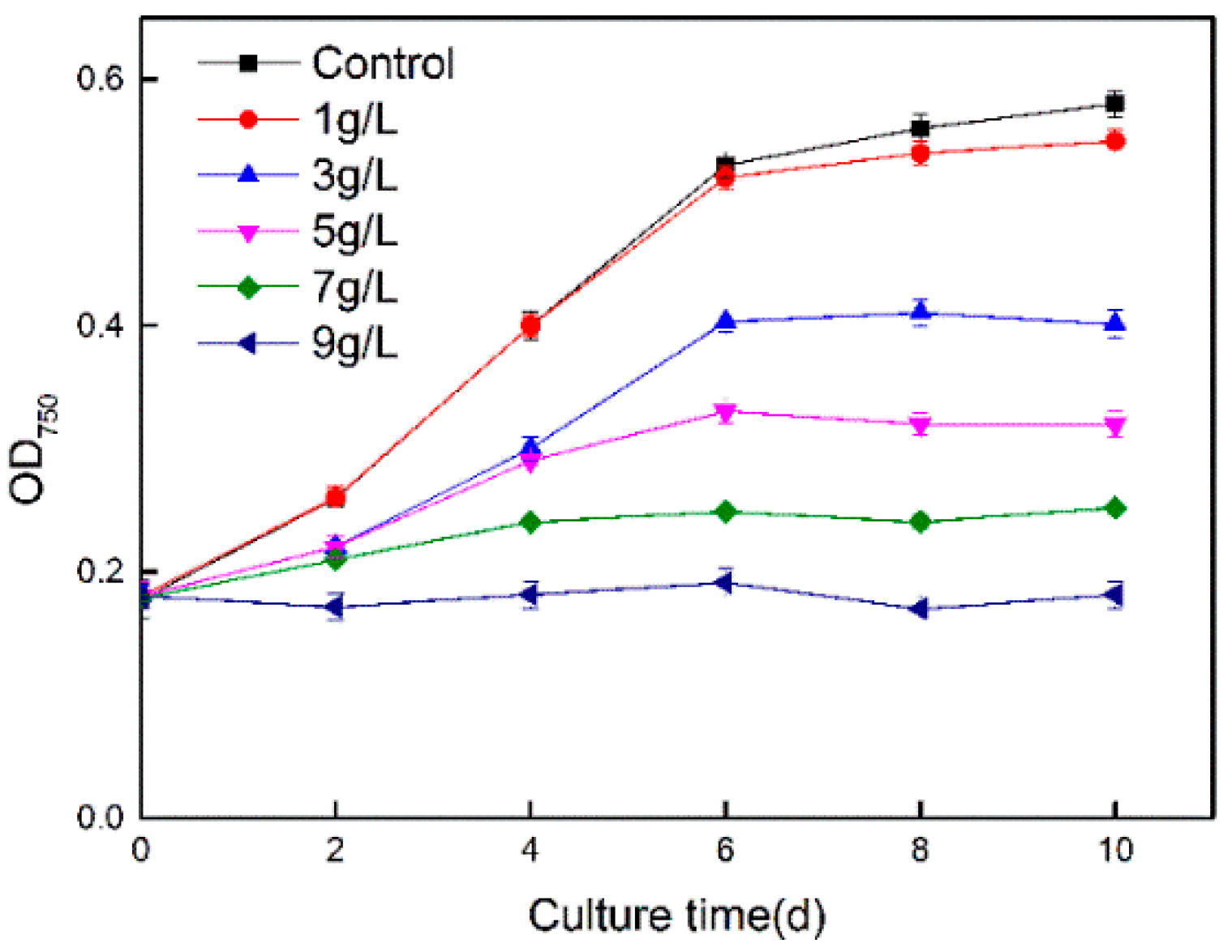
3.1. Impacts of Cs(I) on Cyanobacterial Growth
3.2. Effects of Cs(I) on EPS Production and Composition
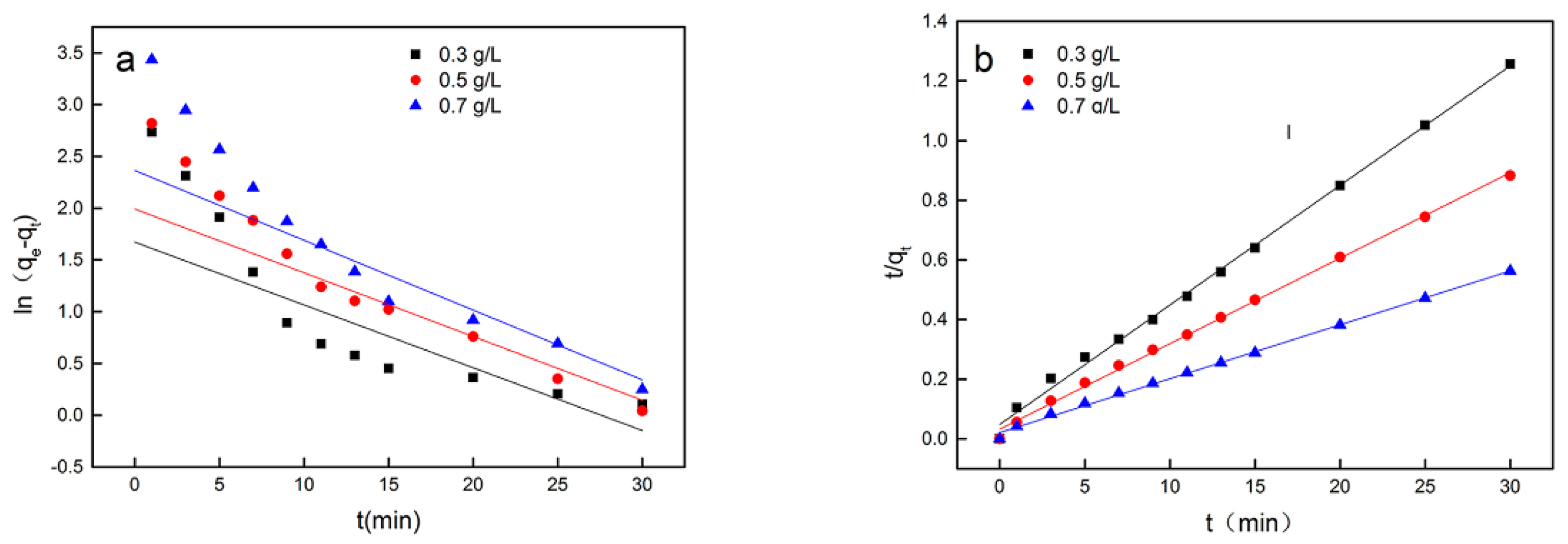
3.3. Adsorption Kinetics of Cs(I) on Synechococcus PCC7002
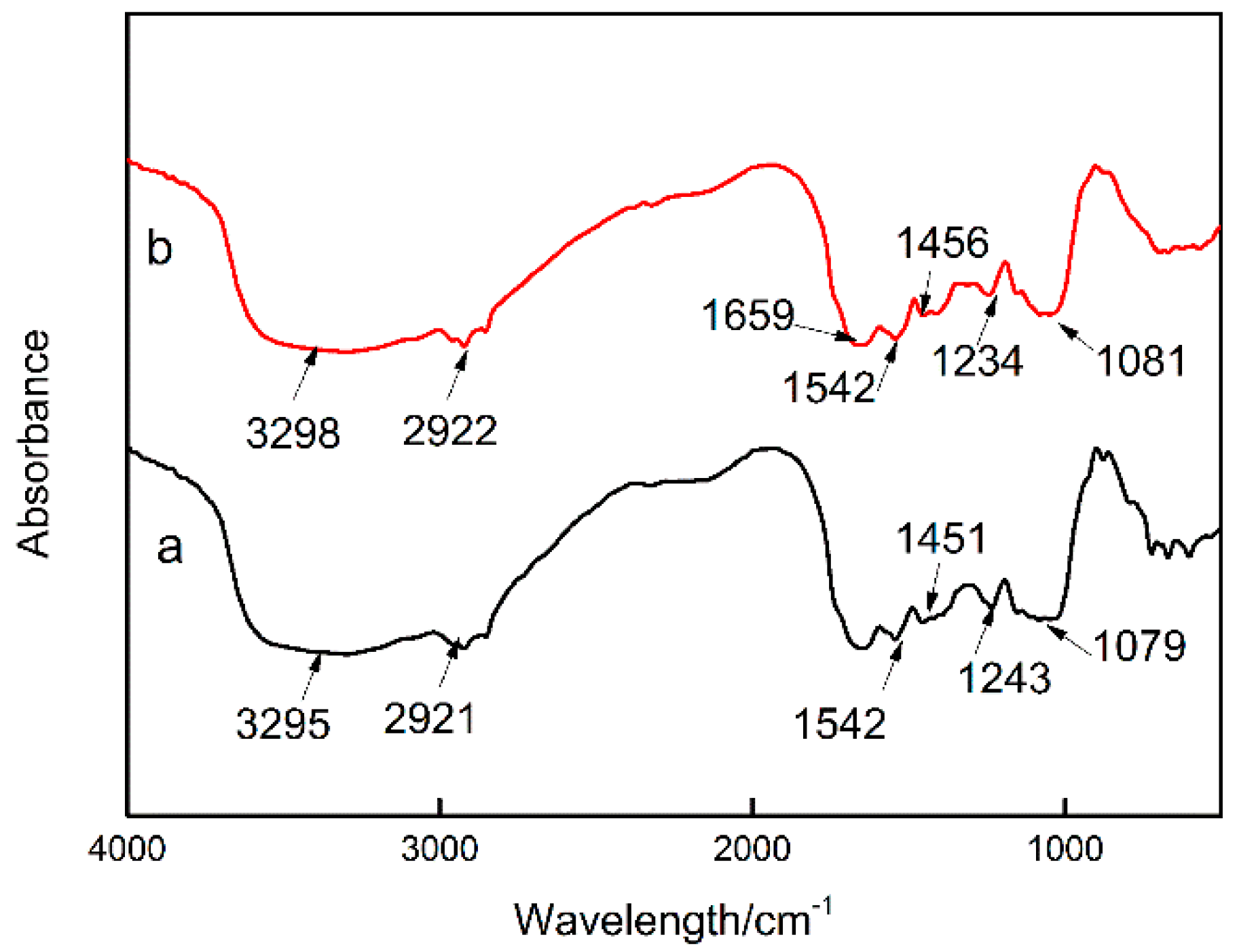
3.4. Effects of Cs(I) on the Chemical Functionalities of Synechococcus PCC7002
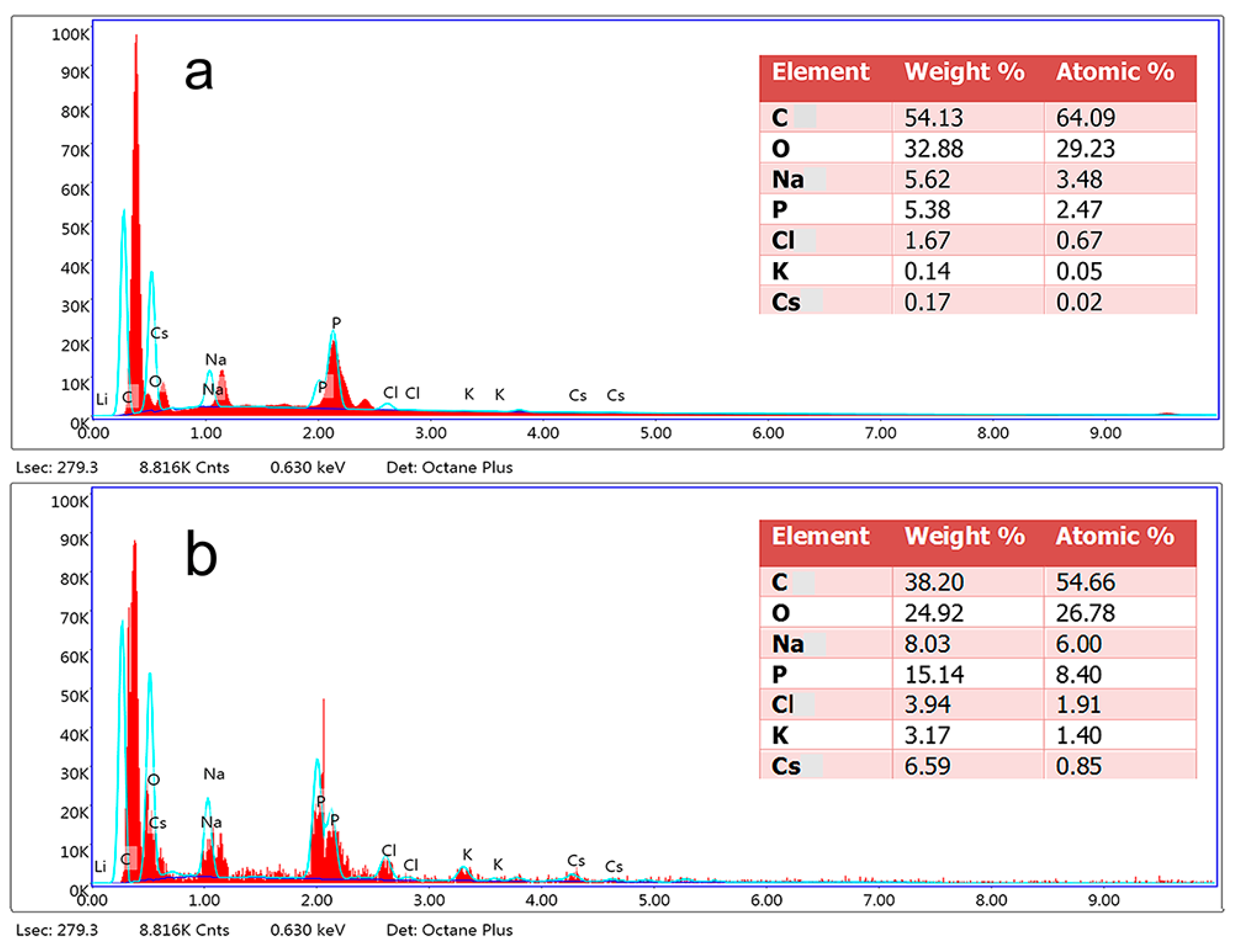
3.5. Effects of Cs(I) on the Surface Morphology and Composition of Synechococcus PCC7002
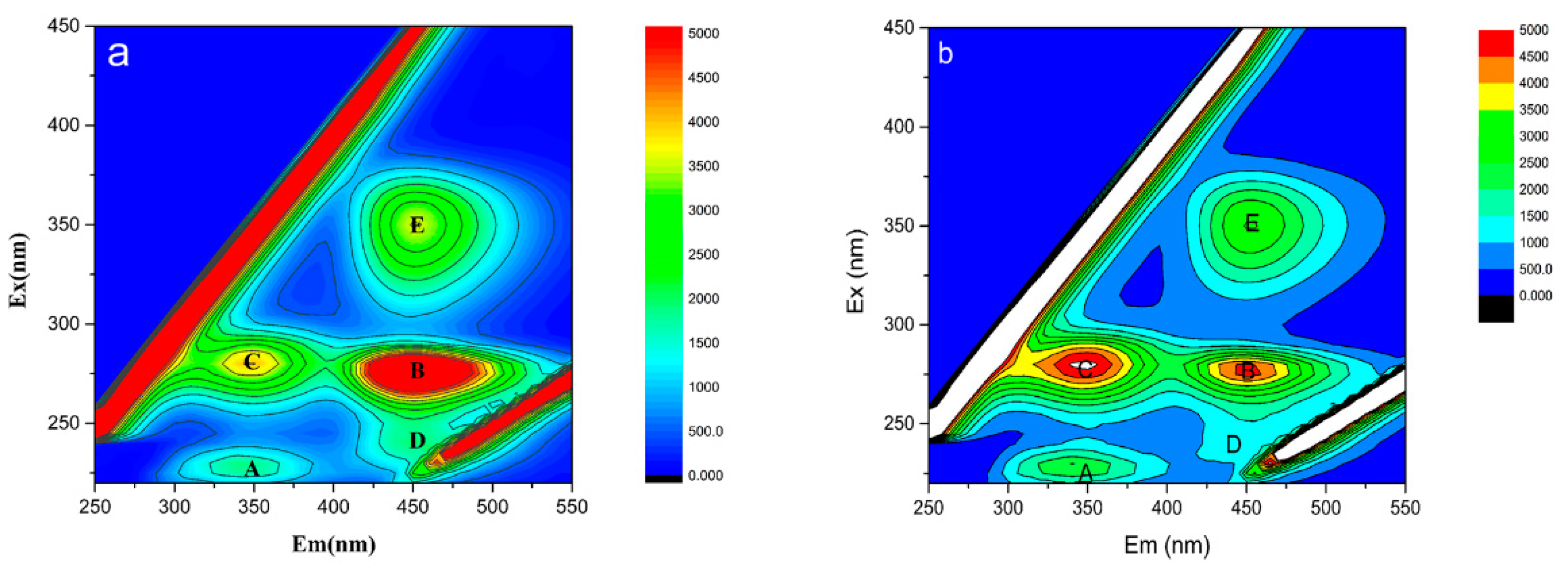
3.6. Effects of Cs(I) on EPS
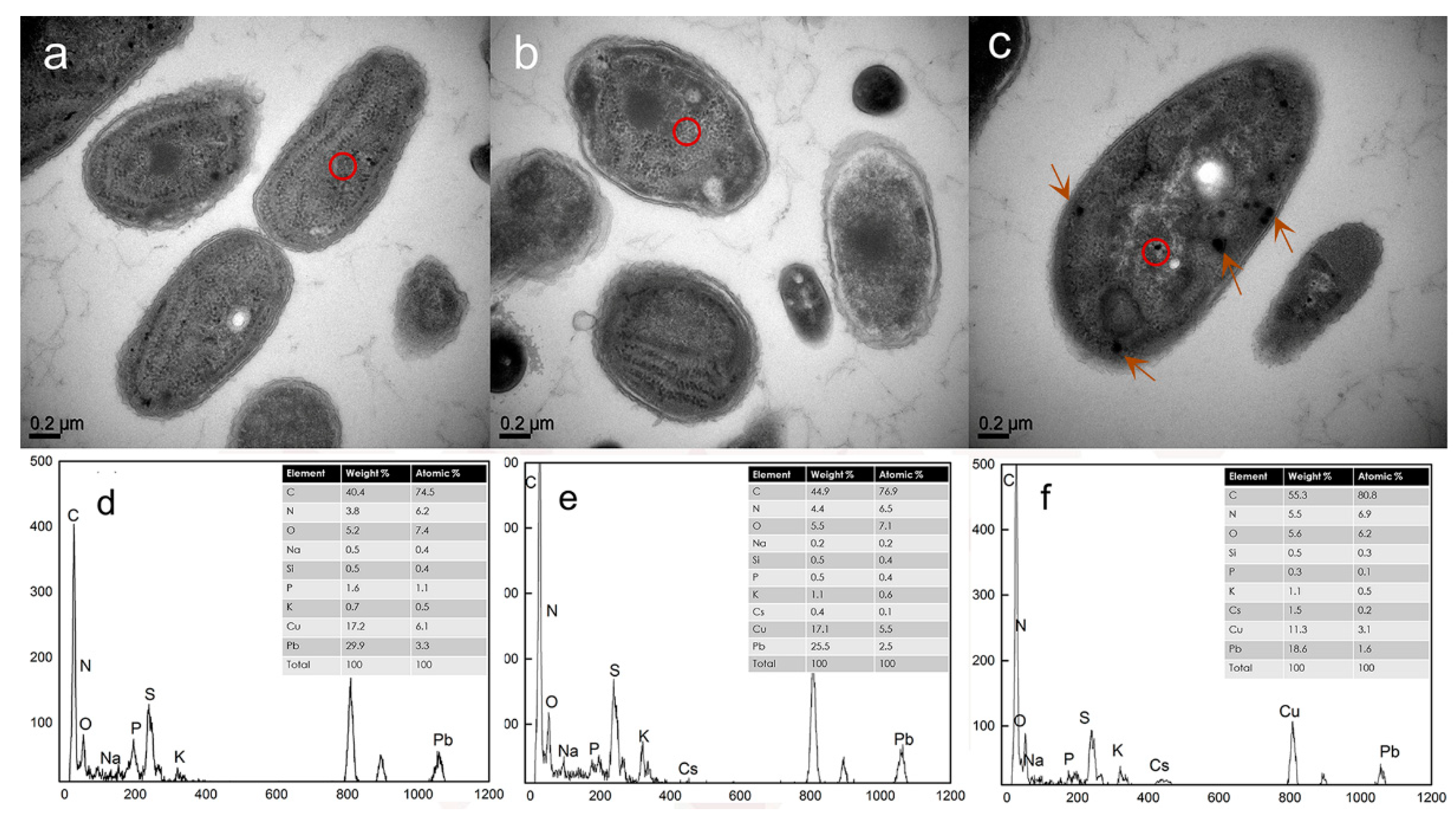
3.7. TEM/EDX Analysis of the Localization of Cs(I) in Synechococcus PCC7002 During Adsorption
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Inoue, K.; Gurung, M.; Adhikari, B.B.; Alam, S.; Kawakita, H.; Ohto, K.; Kurata, M.; Atsumi, K. Adsorptive removal of cesium using bio fuel extraction microalgal waste. J. Hazard. Mater. 2014, 271, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Jalali-Rad, R.; Ghafourian, H.; Asef, Y.; Dalir, S.T.; Sahafipour, M.H.; Gharanjik, B.M. Biosorption of cesium by native and chemically modified biomass of marine algae: Introduce the new biosorbents for biotechnology applications. J. Hazard. Mater. 2004, 116, 125–134. [Google Scholar] [CrossRef]
- Ghorbanzadeh Mashkani, S.; Tajer Mohammad Ghazvini, P. Biotechnological potential of Azolla filiculoides for biosorption of Cs and Sr: Application of micro-PIXE for measurement of biosorption. Bioresour. Technol. 2009, 100, 1915–1921. [Google Scholar] [CrossRef] [PubMed]
- Tomioka, N.; Tanaka, K.; Uchiyama, H.; Yagi, O.; Kokufuta, E. Recovery of 137Cs by a bioaccumulation system using Rhodococcus erythropolis CS98. J. Ferment. Bioeng. 1998, 85, 604–608. [Google Scholar] [CrossRef]
- Karthikeyan, S.; Balasubramanian, R.; Iyer, C.S.P. Evaluation of the marine algae Ulva fasciata and Sargassum sp. for the biosorption of Cu(II) from aqueous solutions. Bioresour. Technol. 2007, 98, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, G.-R.; Lee, D.-J.; Kawamoto, T.; Tanaka, H.; Chen, M.-L.; Luo, Y.-K. Adsorption removal of cesium from drinking waters: A mini review on use of biosorbents and other adsorbents. Bioresour. Technol. 2014, 160, 142–149. [Google Scholar] [CrossRef]
- Mandal, A.; Lahiri, S.; Chemistry, N. Separation of 134Cs and 133Ba radionuclides by calcium alginate beads. J. Radioanal. Nucl. Chem. 2011, 290, 115. [Google Scholar] [CrossRef]
- Drake, L.R.; Lin, S.; Rayson, G.D.; Jackson, P. Chemical modification and metal binding studies of Datura innoxia. Environ. Sci. Technol. 1995, 30, 110–114. [Google Scholar] [CrossRef]
- Schiewer, S. Modelling complexation and electrostatic attraction in heavy metal biosorption by Sargassum biomass. J. Appl. Phycol. 1999, 11, 79–87. [Google Scholar] [CrossRef]
- Holmes, J.D.; Richardson, D.J.; Saed, S.; Evans-Gowing, R.; Russell, D.A.; Sodeau, J.R.J.M. Cadmium-specific formation of metal sulfide ‘Q-particles’ by Klebsiella pneumoniae. Microbiology 1997, 143, 2521–2530. [Google Scholar] [CrossRef] [Green Version]
- Bruins, M.R.; Kapil, S.; Oehme, F.W.J.E.; Safety, E. Microbial resistance to metals in the environment. Ecotoxicol. Environ. Saf. 2000, 45, 198–207. [Google Scholar] [CrossRef]
- Volesky, B.; May, H.; Holan, Z.J.B. Cadmium biosorption by Saccharomyces cerevisiae. Biotechnol. Bioeng. 1993, 41, 826–829. [Google Scholar] [CrossRef] [PubMed]
- Ai, C.; Yan, Z.; Chai, H.; Gu, T.; Wang, J.; Chai, L.; Qiu, G.; Zeng, W.J.J. Increased chalcopyrite bioleaching capabilities of extremely thermoacidophilic Metallosphaera sedula inocula by mixotrophic propagation. J. Ind. Microbiol. Biotechnol. 2019, 46, 1113–1127. [Google Scholar] [CrossRef] [PubMed]
- Avery, S. Caesium accumulation by microorganisms: Uptake mechanisms, cation competition, compartmentalization and toxicity. J. Ind. Microbiol. 1995, 14, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y.; Lee, S.-H.; Lee, J.E.; Lee, S. Biosorption of radioactive cesium from contaminated water by microalgae Haematococcus pluvialis and Chlorella vulgaris. J. Environ. Manag. 2019, 233, 83–88. [Google Scholar] [CrossRef]
- Li, W.-W.; Yu, H.-Q. Insight into the roles of microbial extracellular polymer substances in metal biosorption. Bioresour. Technol. 2014, 160, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Mota, R.; Pereira, S.B.; Meazzini, M.; Fernandes, R.; Santos, A.; Evans, C.A.; De Philippis, R.; Wright, P.C.; Tamagnini, P. Effects of heavy metals on Cyanothece sp. CCY 0110 growth, extracellular polymeric substances (EPS) production, ultrastructure and protein profiles. J. Proteom. 2015, 120, 75–94. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Mishra, A.K. Regulation of calcium ion and its effect on growth and developmental behavior in wild type and ntcA mutant of Anabaena sp. PCC 7120 under varied levels of CaCl2. Microbiology 2014, 83, 235–246. [Google Scholar] [CrossRef]
- Pereira, S.B.; Mota, R.; Vieira, C.P.; Vieira, J.; Tamagnini, P. Phylumwide analysis of genes/proteins related to the last stEPS of assembly and export of EPS (EPS) in cyanobacteria. Sci. Rep.-UK 2015, 5, 14835. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Yang, Y.; Wang, P.; Wang, C.; Miao, L.; Wang, X.; Lv, B.; You, G.; Liu, Z. Effects of CeO2, CuO and ZnO nanoparticles on physiolog-ical features of Microcystisaeruginosa and the productionandcom-position of extracellular polymeric substances. Environ. Sci. Poll. Res. Int. 2016, 24, 110. [Google Scholar] [CrossRef]
- Willis, L.M.; Whitfield, C. Structure, biosynthesis, and function of bacterial capsular polysaccharides synthesized by ABC transporter-dependent pathways. Carbohydr. Res. 2013, 378, 3544. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Alessi, D.; Owttrim, G.; Petrash, D.; Mloszewska, A.; Lalonde, S.; Martinez, R.; Zhou, Q.; Konhauser, K. Cell surface reactivity of Synechococcus sp. PCC 7002: Implications for metal sorption from seawater. Geochim. Cosmochim. Acta 2015, 169, 30–44. [Google Scholar] [CrossRef]
- Ludwig, M.; Bryant, D. Transcription Profiling of the Model Cyanobacterium Synechococcus sp. Strain PCC 7002 by Next-Gen (SOLiD™) Sequencing of cDNA. Front. Microbiol. 2011, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castenholz, R.W. Culturing methods for cyanobacteria. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1988; Volume 167, pp. 68–93. [Google Scholar] [CrossRef]
- Ball, M.M.; Carrero, P.; Castro, D.; Yarzábal, L.A. Mercury resistance in bacterial strains isolated from tailing ponds in a gold mining area near El Callao (Bolívar State, Venezuela). Curr. Microbiol. 2007, 54, 149–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.; Li, Z.; Wang, J.; Liu, A.; Li, Z.; Yu, R.; Wu, X.; Liu, Y.; Li, J.; Zeng, W.J.E.S.; et al. Characterization of extracellular polysaccharide/protein contents during the adsorption of Cd (II) by Synechocystis sp. PCC6803. Environ. Sci. Pollut. Res. 2018, 25, 20713–20722. [Google Scholar] [CrossRef]
- Yu, R.; Liu, Z.; Yu, Z.; Wu, X.; Shen, L.; Liu, Y.; Zeng, W. Relationship among the secretion of extracellular polymeric substances, heat resistance, and bioleaching ability of Metallosphaera sedula. Int. J. Miner. Metall. Mater. 2019, 26, 1504–1511. [Google Scholar] [CrossRef]
- Yang, L.; Chen, J.P. Biosorption of hexavalent chromium onto raw and chemically modified Sargassum sp. Bioresour. Technol. 2008, 99, 297–307. [Google Scholar] [CrossRef]
- Xia, M.; Bao, P.; Liu, A.; Shen, L.; Yu, R.; Liu, Y.; Li, J.; Wu, X.; Huang, C.; Chen, M.; et al. Application of the kinetic and isotherm models for better understanding of the mechanism of biomineralization process induced by Purpureocillium lilacinum Y3. Coll. Surf. B 2019, 181, 207–214. [Google Scholar] [CrossRef]
- Xu, H.; Tan, L.; Dong, H.; He, J.; Liu, X.; Qiu, G. Competitive biosorption behavior of pt(iv) and pd(ii) by providencia vermicola. RSC Adv. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Westerhoff, P.; Leenheer, J.A.; Booksh, K. Fluorescence Excitation−Emission Matrix Regional Integration to Quantify Spectra for Dissolved Organic Matter. Environ. Sci. Technol. 2003, 37, 5701–5710. [Google Scholar] [CrossRef]
- Blasi, B.; Peca, L.; Vass, I.; Kós, P. Characterization of stress responses of heavy metal and metalloid inducible promoters in Synechocystis PCC6803. J. Microbiol. Biotechnol. 2012, 22, 166–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latifi, A.; Ruiz, M.; Zhang, C.-C. Oxidative stress in cyanobacteria. FEMS Microbiol. Rev. 2009, 33, 258–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aquino, S.F.; Stuckey, D.C. Soluble microbial products formation in anaerobic chemostats in the presence of toxic compounds. Water Res. 2004, 38, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Ueshima, M.; Ginn, B.R.; Haack, E.A.; Szymanowski, J.E.S.; Fein, J.B. Cd adsorption onto Pseudomonas putida in the presence and absence of extracellular polymeric substances. Geochim. Cosmochim. Acta 2008, 72, 5885–5895. [Google Scholar] [CrossRef]
- Mikes, J.; Siglova, M.; Cejkova, A.; Masak, J.; Jirku, V. The influence of heavy metals on the production of extracellular polymer substances in the processes of heavy metal ions elimination. Water Sci. Technol. 2005, 52, 151–156. [Google Scholar] [CrossRef]
- Ding, P.; Song, W.; Yang, Z.; Jian, J.J.B.; Engineering, B. Influence of Zn (II) stress-induction on component variation and sorption performance of extracellular polymeric substances (EPS) from Bacillus vallismortis. Bioprocess Biosyst. Eng. 2018, 41, 781–791. [Google Scholar] [CrossRef]
- Yamamura, S.; Amachi, S. Microbiology of inorganic arsenic: From metabolism to bioremediation. J. Biosci. Bioeng. 2014, 118, 1–9. [Google Scholar] [CrossRef]
- Azizian, S.; Science, I. Kinetic models of sorption: A theoretical analysis. J. Coll. Interface Sci. 2004, 276, 47–52. [Google Scholar] [CrossRef]
- Chen, C.; Wang, J. Removal of Pb2+, Ag+, Cs+ and Sr2+ from aqueous solution by brewery’s waste biomass. J. Hazard. Mater. 2008, 151, 65–70. [Google Scholar] [CrossRef]
- Gupta, V.K.; Rastogi, A. Biosorption of lead(II) from aqueous solutions by non-living algal biomass Oedogonium sp. and Nostoc sp.—A comparative study. Coll. Surf. B 2008, 64, 170–178. [Google Scholar] [CrossRef]
- Wang, X.-S.; Huang, J.; Hu, H.-Q.; Wang, J.; Qin, Y. Determination of kinetic and equilibrium parameters of the batch adsorption of Ni (II) from aqueous solutions by Na-mordenite. J. Hazard. Mater. 2007, 142, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Mashitah, M.; Azila, Y.Y.; Bhatia, S. Biosorption of cadmium (II) ions by immobilized cells of Pycnoporus sanguineus from aqueous solution. Bioresour. Technol. 2008, 99, 4742–4748. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; McEwen, G.D.; Wu, Y.; Miller, C.D.; Zhou, A.J.A.; Chemistry, B. Characterization and analysis of mycobacteria and Gram-negative bacteria and co-culture mixtures by Raman microspectroscopy, FTIR, and atomic force microscopy. Anal. Bioanal. Chem. 2013, 405, 1577–1591. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Weakley, A.T.; Aston, D.E.; Rasco, B.A.; Wang, S.; Konkel, M.E. Examination of nanoparticle inactivation of C ampylobacter jejuni biofilms using infrared and R aman spectroscopies. J. Appl. Microbiol. 2012, 113, 952–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suo, Z.; Yang, X.; Avci, R.; Kellerman, L.; Pascual, D.W.; Fries, M.; Steele, A.J.L. HEPES-stabilized encapsulation of Salmonella typhimurium. Langmuir 2007, 23, 1365–1374. [Google Scholar] [CrossRef]
- Merroun, M.L.; Ben Chekroun, K.; Arias, J.M.; González-Muñoz, M.T. Lanthanum fixation by Myxococcus xanthus: Cellular location and extracellular polysaccharide observation. Chemosphere 2003, 52, 113–120. [Google Scholar] [CrossRef]
- Zhang, D.; Pan, X.; Mostofa, K.M.G.; Chen, X.; Mu, G.; Wu, F.; Liu, J.; Song, W.; Yang, J.; Liu, Y.; et al. Complexation between Hg(II) and biofilm extracellular polymeric substances: An application of fluorescence spectroscopy. J. Hazard. Mater. 2010, 175, 359–365. [Google Scholar] [CrossRef]
- Chen, B.; Li, F.; Liu, N.; Ge, F.; Xiao, H.; Yang, Y. Role of extracellular polymeric substances from Chlorella vulgaris in the removal of ammonium and orthophosphate under the stress of cadmium. Bioresour. Technol. 2015, 190, 299–306. [Google Scholar] [CrossRef]
- Chen, J.; Gu, B.; LeBoeuf, E.J.; Pan, H.; Dai, S.J.C. Spectroscopic characterization of the structural and functional properties of natural organic matter fractions. Chemosphere 2002, 48, 59–68. [Google Scholar] [CrossRef]
- Sivaperumal, P.; Kamala, K.; Rajaram, R.J.E.S.; Research, P. Adsorption of cesium ion by marine actinobacterium Nocardiopsis sp. 13H and their extracellular polymeric substances (EPS) role in bioremediation. Environ. Sci. Pollut. Res. 2018, 25, 4254–4267. [Google Scholar] [CrossRef]
- Nagpal, U.M.; Bankar, A.V.; Pawar, N.J.; Kapadnis, B.P.; Zinjarde, S.S.J.W.; Pollution, S. Equilibrium and kinetic studies on biosorption of heavy metals by leaf powder of paper mulberry (Broussonetia papyrifera). Water Air Soil Pollut. 2011, 215, 177–188. [Google Scholar] [CrossRef]
- Ang, X.; Sethu, V.; Andresen, J.; Sivakumar, M.J.C.T.; Policy, E. Copper (II) ion removal from aqueous solutions using biosorption technology: Thermodynamic and SEM–EDX studies. Clean Technol. Environ. Policy 2013, 15, 401–407. [Google Scholar] [CrossRef]
- Wang, J.; Chen, C.J.B.A. Biosorption of heavy metals by Saccharomyces cerevisiae: A review. Biotechnol. Adv. 2006, 24, 427–451. [Google Scholar] [CrossRef] [PubMed]
- Fourest, E.; Volesky, B.J.E.S. Contribution of sulfonate groups and alginate to heavy metal biosorption by the dry biomass of Sargassum fluitans. Environ. Sci. Technol. 1995, 30, 277–282. [Google Scholar] [CrossRef]
- Lan, T.; Feng, Y.; Liao, J.; Li, X.; Ding, C.; Zhang, D.; Yang, J.; Zeng, J.; Yang, Y.; Tang, J.; et al. Biosorption behavior and mechanism of cesium-137 on Rhodosporidium fluviale strain UA2 isolated from cesium solution. J. Environ. Radioact. 2014, 134, 6–13. [Google Scholar] [CrossRef]
- Stancu, M.M. Physiological cellular responses and adaptations of Rhodococcus erythropolis IBBPo1 to toxic organic solvents. J. Environ. Sci. 2014, 26, 2065–2075. [Google Scholar] [CrossRef]
- Hosseinkhani, B.; Hennebel, T.; Van Nevel, S.; Verschuere, S.; Yakimov, M.M.; Cappello, S.; Blaghen, M.; Boon, N. Biogenic Nanopalladium Based Remediation of Chlorinated Hydrocarbons in Marine Environments. Environ. Sci. Technol. 2014, 48, 550–557. [Google Scholar] [CrossRef]
- Maes, S.; Props, R.; Fitts, J.P.; De Smet, R.; Vanhaecke, F.; Boon, N.; Hennebel, T. Biological recovery of platinum complexes from diluted aqueous streams by axenic cultures. PLoS ONE 2017, 12, e0169093. [Google Scholar] [CrossRef]
- Avery, S. Microbial interactions with caesium—Implications for biotechnology. J. Chem. Technol. Biotechnol. Int. Res. Process Environ. Clean Technol. 1995, 62, 3–16. [Google Scholar] [CrossRef]
- Volesky, B.; May-Phillips, H.J.A.M. Biosorption of heavy metals by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 1995, 42, 797–806. [Google Scholar] [CrossRef]
- Suzuki, Y.; Banfield, J. Resistance to, and accumulation of, uranium by bacteria from a uranium-contaminated site. Geomicrobiol. J. 2004, 21, 113–121. [Google Scholar] [CrossRef]
- Strandberg, G.W.; Shumate, S.E.; Parrott, J. Microbial cells as biosorbents for heavy metals: Accumulation of uranium by Saccharomyces cerevisiae and Pseudomonas aeruginosa. Appl. Environ. Microbiol. 1981, 41, 237–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avery, S.; Codd, G.; Gadd, G. Interactions of cyanobacteria and microalgae with caesium. Impact Heavy Metals Environ. 1992, 54, 133–182. [Google Scholar]
- Turner, J.S.; Robinson, N.J. Cyanobacterial metallothioneins biochemistry and molecular-genetics. J. Ind. Microbiol. 1995, 14, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, S.; Tripathi, R.M.; Hegde, A.G. Biosorption of heavy metals and radionuclide from aqueous solutions by pre-treated arca shell biomass. J. Hazard. Mater. 2008, 150, 376–386. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomass | Initial Concentration | First-Order Model | Second-Order Model | ||||
|---|---|---|---|---|---|---|---|
| (mg L−1) | qe | K1 | R2 | qe | K2 | R2 | |
| Synechococcus | 300 | 8.49 | 0.085 | 0.84 | 25.4 | 0.025 | 0.99 |
| PCC7002 | 500 | 12.75 | 0.091 | 0.84 | 35.46 | 0.019 | 0.99 |
| 700 | 20.59 | 0.102 | 0.78 | 56.27 | 0.011 | 0.99 | |
| Biomass | T/℃ | Langmuir Constant | Freundlich Constant | ||||
|---|---|---|---|---|---|---|---|
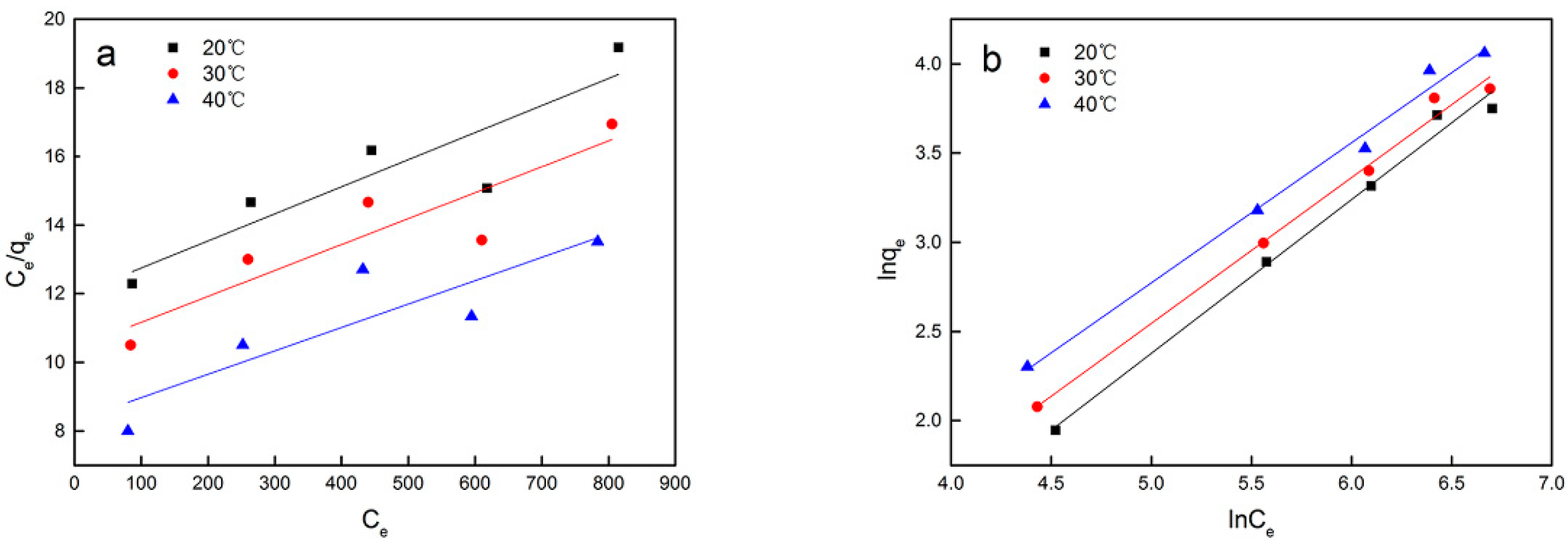
| B (L mg−1) | Qm (mg g−1) | R2 | n | KF (mg g−1) | R2 | ||
| Synechococcus | 20 | 0.00066 | 126.74 | 0.84 | 1.16 | 0.145 | 0.99 |
| PCC7002 | 30 | 0.00072 | 134.05 | 0.84 | 1.22 | 0.214 | 0.99 |
| 40 | 0.00082 | 146.84 | 0.78 | 1.27 | 0.223 | 0.99 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, R.; Chai, H.; Yu, Z.; Wu, X.; Liu, Y.; Shen, L.; Li, J.; Ye, J.; Liu, D.; Ma, T.; et al. Behavior and Mechanism of Cesium Biosorption from Aqueous Solution by Living Synechococcus PCC7002. Microorganisms 2020, 8, 491. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040491
Yu R, Chai H, Yu Z, Wu X, Liu Y, Shen L, Li J, Ye J, Liu D, Ma T, et al. Behavior and Mechanism of Cesium Biosorption from Aqueous Solution by Living Synechococcus PCC7002. Microorganisms. 2020; 8(4):491. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040491
Chicago/Turabian StyleYu, Runlan, Hongsheng Chai, Zhaojing Yu, Xueling Wu, Yuandong Liu, Li Shen, Jiaokun Li, Jun Ye, Danchan Liu, Tao Ma, and et al. 2020. "Behavior and Mechanism of Cesium Biosorption from Aqueous Solution by Living Synechococcus PCC7002" Microorganisms 8, no. 4: 491. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040491