Screening, Identification and Efficacy Evaluation of Antagonistic Bacteria for Biocontrol of Soft Rot Disease Caused by Dickeya zeae

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Bacterial Strains
2.2. Screening of Antagonistic Bacteria
2.3. Identification of Antagonistic Bacteria
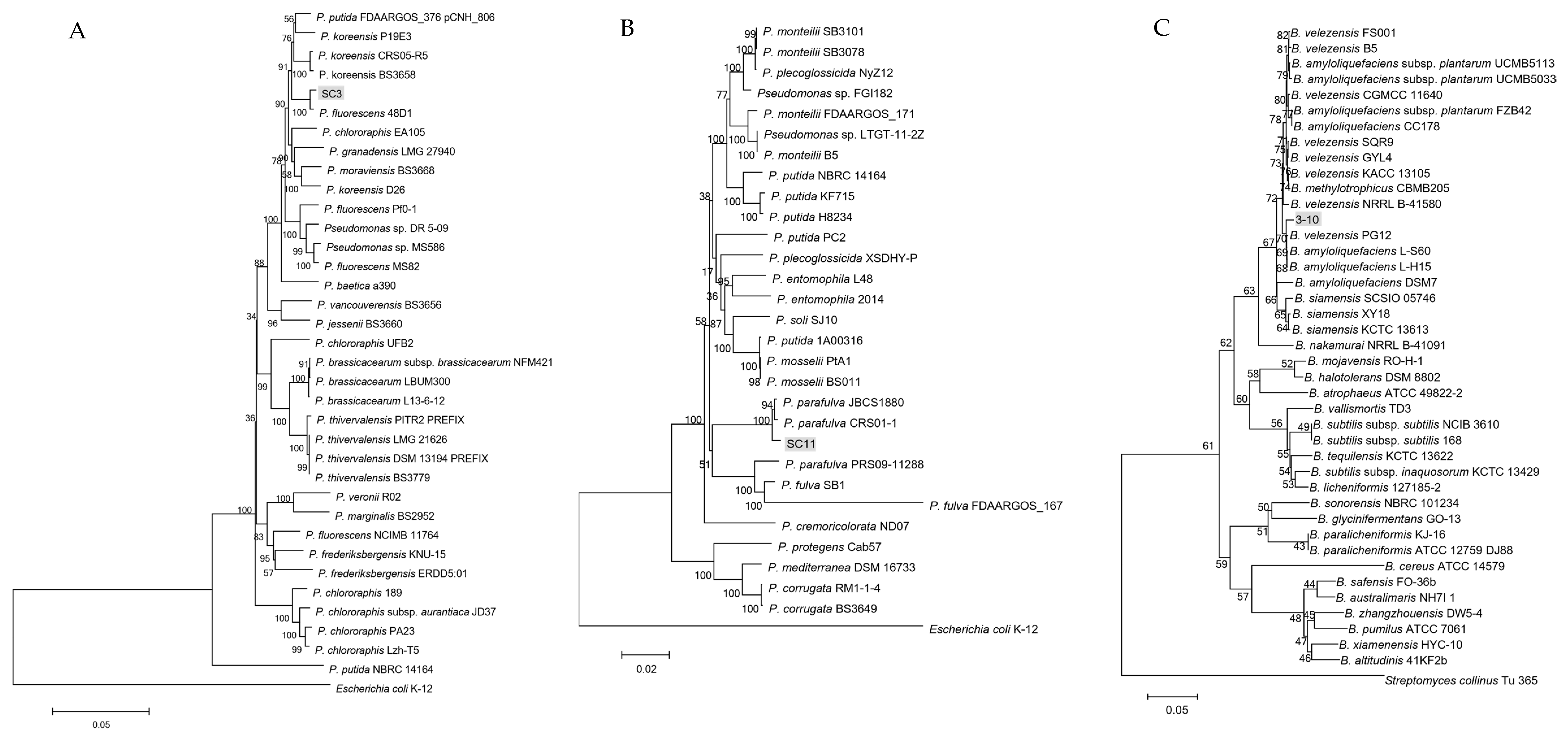
2.4. Phylogenetic Analysis of Antagonistic Bacteria
2.5. Determination of Antimicrobial Spectrum of Strains
2.6. Determination of Greenhouse Control Effect of Antagonistic Bacteria
2.7. Testing of Antibiosis Activities of the Antagonistic Bacteria in Liquid and Solid Media
3. Results
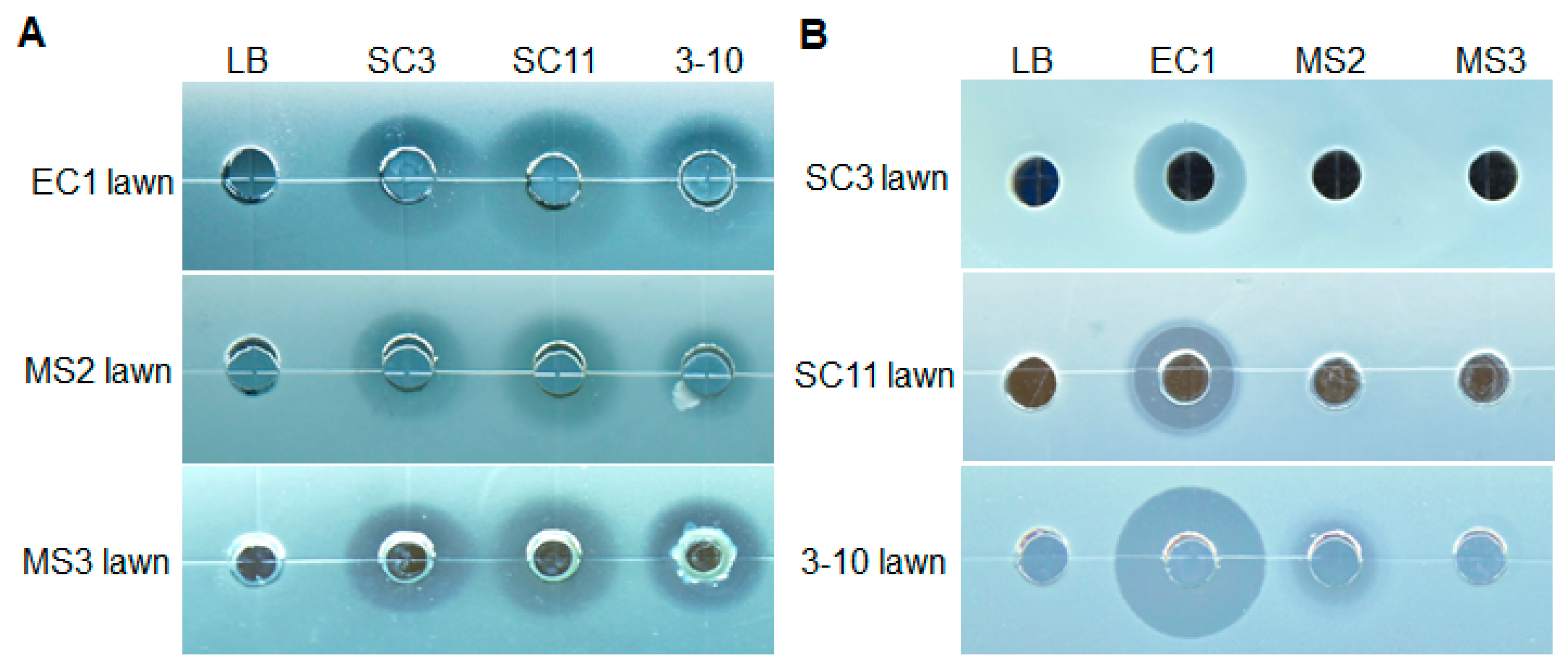
3.1. Isolation of Strains with Antagonistic Activities
3.2. Susceptibility of three Antagonistic Bacteria to Metabolites Secreted by D. zeae Strains
3.3. Identification of Antagonistic Strains SC3, SC11 and 3-10
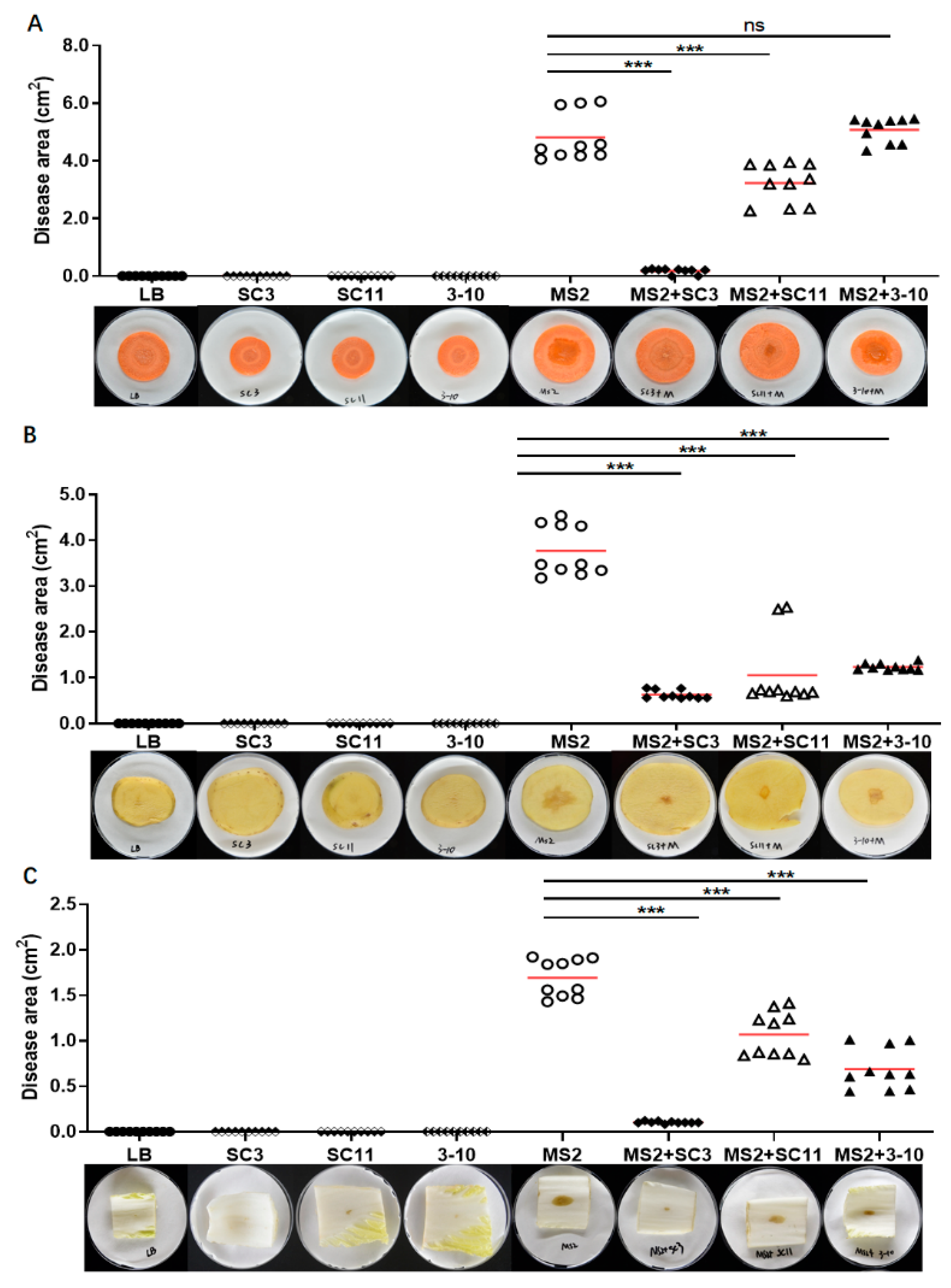
3.4. Antimicrobial Spectrum of Antagonistic Bacteria SC3, SC11 and 3-10
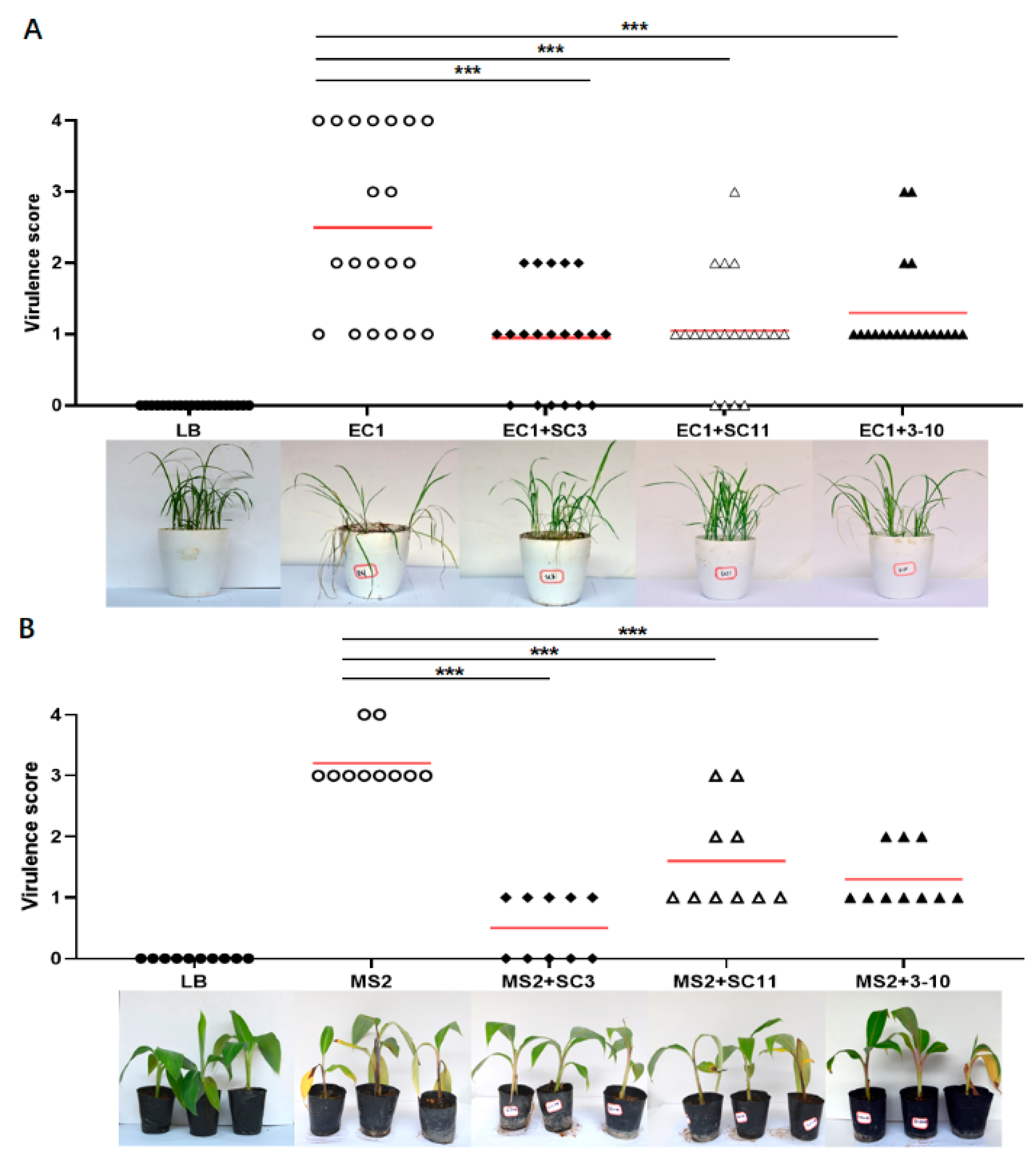
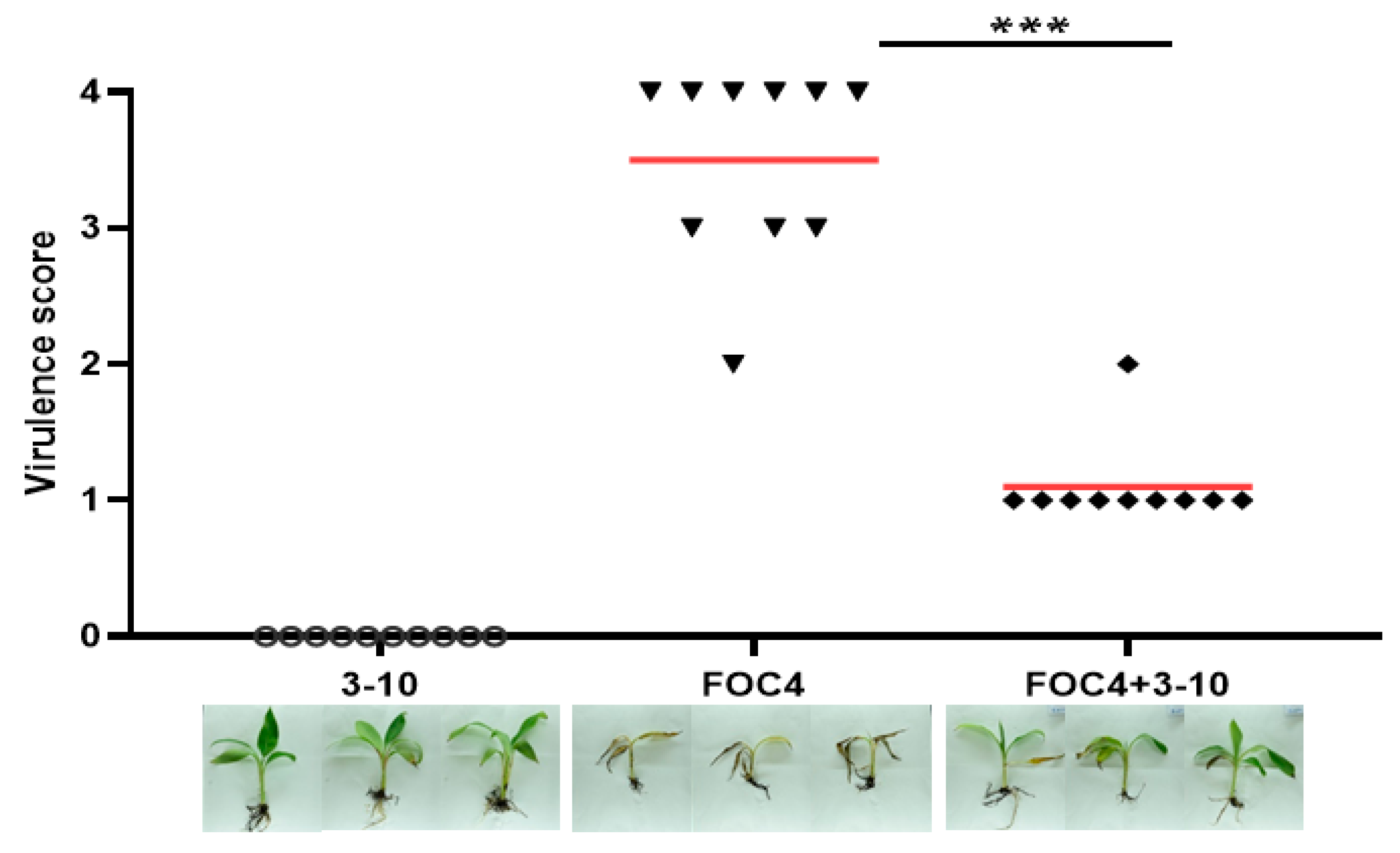
3.5. Greenhouse Control Effect of Antagonistic Bacteria SC3, SC11 and 3-10
3.6. Bacteriostatic Activities in Solid and Liquid Culture Conditions
3.7. The stability of Bacteriostatic Metabolites from the Three Antagonistic Bacteria
4. Discussion
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Samson, R.G.; Legendre, J.B.; Christen, R.; Saux, M.F.; Achouak, W.; Gardan, L. Transfer of Pectobacterium chrysanthemi (Burkholder et al. 1953) Brenner et al. 1973 and Brenneria paradisiaca to the genus Dickeya gen. nov. as Dickeya chrysanthemi comb. nov. and Dickeya paradisiaca comb. nov. and delineation of four novel species, Dickeya dadantii sp. nov., Dickeya dianthicola sp. nov., Dickeya dieffenbachiae sp. nov. and Dickeya zeae sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 1415–1427. [Google Scholar] [PubMed]
- Goto, M. Bacterial Foot Rot of Rice Caused by a Strain of Erwinia chrysanthemi. Phytopathology 1979, 69, 213–216. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.G.; Wang, Z.Z. Infection characteristics of Erwinia chrysanthemi pv. zeae on rice. J. South China Agric. Univ. 2004, 25, 55–57. [Google Scholar]
- Zhang, J.X.; Shen, H.F.; Pu, X.M.; Lin, B.R.; Hu, J. Identification of Dickeya zeae as a causal agent of bacterial soft rot in banana in China. Plant Dis. 2014, 98, 436–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Hunjan, M.S.; Kaur, H.; Dhillon, H.K.; Singh, P.P. Biochemical responses associated with resistance to bacterial stalk rot caused by Dickeya zeae in maize. J. Phytopathol. 2017, 165, 822–832. [Google Scholar] [CrossRef]
- Hu, M.; Li, J.; Chen, R.; Li, W.; Feng, L.; Shi, L.; Xue, Y.; Feng, X.; Zhang, L.; Zhou, J. Dickeya zeae strains isolated from rice, banana and clivia rot plants show great virulence differentials. BMC Microbiol. 2018, 18, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.; Zhang, H.; Xu, J.; Liu, Q.; Jiang, Z.; Zhang, L. The acyl-homoserine lactone-type quorum-sensing system modulates cell motility and virulence of Erwinia chrysanthemi pv. zeae. J. Bacteriol. 2008, 190, 1045–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nassar, A.; Bertheau, Y.; Dervin, C.; Narcy, J.P.; Lemattre, M. Ribotyping of Erwinia chrysanthemi strains in relation to their pathogenic and geographic distribution. Appl. Environ. Microbiol. 1994, 60, 3781–3789. [Google Scholar] [CrossRef] [Green Version]
- Sinha, S.K.; Prasad, M. Bacterial stalk rot of Maize, its symptoms and host-range. Zent. Bakteriol. Parasitenkd. Infekt. Hyg. 1977, 132, 81–88. [Google Scholar] [CrossRef]
- Zhao, L.W.; Xu, L.Y.; Chen, Y.B.; Zhang, X.M.; Hu, Y.F.; Jiang, T.M. Discussion on the causes of rice bacterial rot epidemic disease in Ningbo. China Rice 2017, 23, 100–102. (In Chinese) [Google Scholar]
- Ying, D.W. Occurrence characteristics and control measures of rice bacterial base rot in Yanping District. China Agric. Technol. Ext. 2017, 33, 57–58. (In Chinese) [Google Scholar]
- Wang, C.L.; Tai, D.L.; Mei, A.Z.; Chen, Q. Factors contributing to the occurrence of severe rice bacterial foot rot in 2016 in Dongtai City and relevant countermeasures. Barley Cereal Sci. 2017, 34, 48–50. (In Chinese) [Google Scholar]
- Liu, F.Y.; Wang, T.; Ding, H.H. Analysis of the causes of rice bacterial rot in Shayang County. Hubei Plant Prot. 2016, 1, 57–58. (In Chinese) [Google Scholar]
- Pan, Z.Q. Occurrence and control measures of rice foot rot in Sandu County. Agric. Technol. Serv. 2016, 33, 77. (In Chinese) [Google Scholar]
- Bertani, I.; Passos da Silva, D.; Abbruscato, P.; Piffanelli, P.; Venturi, V. Draft genome sequence of the plant pathogen Dickeya zeae DZ2Q, isolated from rice in Italy. Genome Announc. 2013, 1, e00905–e00913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, B.R.; Shen, H.F.; Pu, X.M.; Tian, X.S.; Zhao, W.J.; Zhu, S.F.; Dong, M.M. First report of a soft Rot of banana in mainland China caused by a Dickeya sp. (Pectobacterium chrysanthemi). Plant Dis. 2010, 94, 640. [Google Scholar] [CrossRef]
- Jafra, S.; Przysowa, J.; Gwizdek-Wisniewska, A.; van der Wolf, J.M. Potential of bulb-associated bacteria for biocontrol of hyacinth soft rot caused by Dickeya zeae. J. Appl. Microbiol. 2009, 106, 268–277. [Google Scholar] [CrossRef]
- Wu, J.P.; Song, Z.H.; Xiang, F.Y.; Zeng, X.G.; Gu, Y.C. Resistance mechanism of antagonistic bacterium in plant disease biocontrol. Hubei Agric. Sci. 2009, 48, 2286–2288. (In Chinese) [Google Scholar]
- Emmert, E.A.; Handelsman, J. Biocontrol of plant disease: A (gram-) positive perspective. Fems Microbiol. Lett. 1999, 171, 1–9. [Google Scholar] [CrossRef]
- Czajkowski, R.; Pérombelon, M.C.M.; van Veen, J.A.; van der Wolf, J.M. Control of blackleg and tuber soft rot of potato caused by Pectobacterium and Dickeya species: A review. Plant Pathol. 2011, 60, 999–1013. [Google Scholar] [CrossRef]
- Diallo, S.; Crepin, A.; Barbey, C.; Orange, N.; Burini, J.F.; Latour, X. Mechanisms and recent advances in biological control mediated through the potato rhizosphere. Fems Microbiol. Ecol. 2011, 75, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Hadizadeh, I.; Peivastegan, B.; Hannukkala, A.; van der Wolf, J.M.; Nissinen, R.; Pirhonen, M. Biological control of potato soft rot caused by Dickeya solani and the survival of bacterial antagonists under cold storage conditions. Plant Pathol. 2019, 68, 297–311. [Google Scholar] [CrossRef] [Green Version]
- Krzyzanowska, D.M.; Maciag, T.; Siwinska, J.; Krychowiak, M.; Jafra, S.; Czajkowski, R. Compatible mixture of bacterial antagonists developed to protect potato tubers from soft rot caused by Pectobacterium spp. and Dickeya spp. Plant Dis. 2019, 103, 1374–1382. [Google Scholar] [CrossRef] [PubMed]
- Raoul, D.E.Y.; Cigna, J.; Quetu-Laurent, A.; Caron, A.; Munier, E.; Beury-Cirou, A.; Helias, V.; Faure, D. Biocontrol of the potato blackleg and soft rot diseases caused by Dickeya dianthicola. Appl. Environ. Microbiol. 2016, 82, 268–278. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Liu, T.; Wei, M.; Zhu, Z.; Liu, W.; Zhang, Z. Macrolactin a is the key antibacterial substance of Bacillus amyloliquefaciens D2WM against the pathogen Dickeya chrysanthemi. Eur. J. Plant Pathol. 2019, 155, 393–404. [Google Scholar] [CrossRef]
- Wang, Y.H.; Tan, Z.Q.; Liu, H.; Zhang, R.Y. Screening and identification of antagonistic bacteria against bacterial soft rot of Guzmania denise. Guangdong Agric. Sci. 2012, 39, 63–64. (In Chinese) [Google Scholar]
- Cao, Y.; Pi, H.; Chandrangsu, P.; Li, Y.; Wang, Y.; Zhou, H.; Xiong, H.; Helmann, J.D.; Cai, Y. Antagonism of two plant-growth promoting Bacillus velezensis isolates against Ralstonia solanacearum and Fusarium oxysporum. Sci Rep-Uk 2018, 8, 4360. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhang, H.; Wu, J.; Liu, Q.; Xi, P.; Lee, J.; Liao, J.; Jiang, Z.; Zhang, L. A novel multidomain polyketide synthase is essential for zeamine production and the virulence of Dickeya zeae. Mol. Plant Microbe Interact. 2011, 24, 1156–1164. [Google Scholar] [CrossRef] [Green Version]
- Fanelli, V.; Cariddi, C.; Finetti-Sialer, M. Selective detection of Pseudomonas syringae pv. tomato using dot blot hybridization and real-time PCR. Plant Pathol. 2007, 56, 683–691. [Google Scholar]
- Toro, M.; Ramirez-Bahena, M.H.; Cuesta, M.J.; Velazquez, E.; Peix, A. Pseudomonas guariconensis sp. nov., isolated from rhizospheric soil. Int. J. Syst. Evol. Microbiol. 2013, 63, 4413–4420. [Google Scholar] [CrossRef]
- Ramos, E.; Ramirez-Bahena, M.H.; Valverde, A.; Velazquez, E.; Zuniga, D.; Velezmoro, C.; Peix, A. Pseudomonas punonensis sp. nov., isolated from straw. Int. J. Syst. Evol. Microbiol. 2013, 63, 1834–1839. [Google Scholar] [CrossRef] [PubMed]
- Tayeb, L.A.; Ageron, E.; Grimont, F.; Grimont, P.A.D. Molecular phylogeny of the genus Pseudomonas based on rpoB sequences and application for the identification of isolates. Res. Microbiol. 2005, 156, 763–773. [Google Scholar] [CrossRef]
- Mulet, M.; Bennasar, A.; Lalucat, J.; Garcia-Valdes, E. An rpoD-based PCR procedure for the identification of Pseudomonas species and for their detection in environmental samples. Mol. Cell Probes 2009, 23, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Bae, K.S. Phylogenetic analysis of Bacillus subtilis and related taxa based on partial gyrA gene sequences. Antonie Van Leeuwenhoek 2000, 78, 123–127. [Google Scholar] [CrossRef]
- Yamada, S.; Ohashi, E.; Agata, N.; Venkateswaran, K. Cloning and nucleotide sequence analysis of gyrB of Bacillus cereus, B. thuringiensis, B. mycoides, and B. anthracis and their application to the detection of B. cereus in rice. Appl. Environ. Microbiol. 1999, 65, 1483–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Nuoqiao, L.; Yumei, C.; Zhibin, L.; Lisheng, L.; Mingfa, L.; Yufan, C.; Yingxin, T.; Fei, H.; Shaohua, C. Biocontrol of sugarcane smut disease by interference of fungal sexual mating and hyphal growth using a bacterial isolate. Front. Microbiol. 2017, 8, 778. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Zhou, T.; Li, Q.; Xu, X.; Fan, X.; Zhang, L.; Chen, S. Cupriavidus sp. HN-2, a novel quorum quenching bacterial isolate, is a potent biocontrol agent against Xanthomonas campestris pv. campestris. Microorganisms 2019, 8, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Wu, J.; Yin, W.; Li, P.; Zhou, J.; Chen, S.; He, F.; Cai, J.; Zhang, L.H. Diffusible signal factor family signals provide a fitness advantage to Xanthomonas campestris pv. campestris in interspecies competition. Environ. Microbiol. 2016, 18, 1534–1545. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.N.; Liu, S.Y.; Chen, Y.F.; Liao, L.S. First report of Pantoea anthophila causing soft rot disease in Clausena lansium (Wampee) in China. Plant Dis. 2015, 99, 416. [Google Scholar] [CrossRef]
- Li, M.H.; Xie, X.L.; Lin, X.F.; Shi, J.X.; Ding, Z.J.; Ling, J.F.; Xi, P.G.; Zhou, J.N.; Leng, Y.; Zhong, S.; et al. Functional characterization of the gene FoOCH1 encoding a putative alpha-1,6-mannosyltransferase in Fusarium oxysporum f. sp. cubense. Fungal Genet. Biol. 2014, 65, 1–13. [Google Scholar] [CrossRef]
- Shu, C.W.; Zou, C.J.; Chen, J.L.; Tang, F.; Zhou, E.X. Genetic diversity and population structure of Rhizoctonia solani AG-1 IA, the causal agent of rice sheath blight, in South China. Can. J. Plant Pathol. 2014, 36, 179–186. [Google Scholar] [CrossRef]
- Zhang, S.; Liang, M.; Naqvi, N.I.; Lin, C.; Qian, W.; Zhang, L.H.; Deng, Y.Z. Phototrophy and starvation-based induction of autophagy upon removal of Gcn5-catalyzed acetylation of Atg7 in Magnaporthe oryzae. Autophagy 2017, 13, 1318–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, L.; Schaefer, A.L.; Hu, M.; Chen, R.; Greenberg, E.P.; Zhou, J. Virulence factor identification in the banana pathogen Dickeya zeae MS2. Appl. Environ. Microbiol. 2019, 85, e1611–e1619. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.G.; Zhong, W.Z.; Ming, Z.G.; Kang, O.Z.; Chen, Y.T.; Agr, S.C. Rice resistance to Erwinia chrysanthemi. J. South China Agric. Univ. 2003, 24, 89–90. (In Chinese) [Google Scholar]
- Liao, L.; Cheng, Y.; Liu, S.; Zhou, J.; An, S.; Lv, M.; Chen, Y.; Gu, Y.; Chen, S.; Zhang, L. Production of novel antibiotics Zeamines through optimizing Dickeya zeae fermentation conditions. PLoS ONE 2014, 9, e116047. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Cheng, Y.; Lv, M.; Liao, L.; Chen, Y.; Gu, Y.; Liu, S.; Jiang, Z.; Xiong, Y.; Zhang, L. The complete genome sequence of Dickeya zeae EC1 reveals substantial divergence from other Dickeya strains and species. BMC Genom. 2015, 16, 571. [Google Scholar] [CrossRef] [Green Version]
- Keel, C.; Weller, D.M.; Natsch, A.; Defago, G.; Cook, R.J.; Thomashow, L.S. Conservation of the 2,4-diacetylphloroglucinol biosynthesis locus among fluorescent Pseudomonas strains from diverse geographic locations. Appl. Environ. Microbiol. 1996, 62, 552–563. [Google Scholar] [CrossRef] [Green Version]
- Brodhagen, M.; Henkels, M.D.; Loper, J.E. Positive autoregulation and signaling properties of pyoluteorin, an antibiotic produced by the biological control organism Pseudomonas fluorescens Pf-5. Appl. Environ. Microbiol. 2004, 70, 1758–1766. [Google Scholar] [CrossRef] [Green Version]
- Pfender, W.F.; Kraus, J.; Loper, J.E. A genomic region from Pseudomonas fluorescens Pf-5 required for pyrrolnitrin production and inhibition of Pyrenophora tritici-repentis in wheat straw. Phytopathology 1993, 83, 1223–1228. [Google Scholar] [CrossRef]
- Timms-Wilson, T.M.; Ellis, R.J.; Renwick, A.; Rhodes, D.J.; Mavrodi, D.V.; Weller, D.M.; Thomashow, L.S.; Bailey, M.J. Chromosomal insertion of phenazine-1-carboxylic acid biosynthetic pathway enhances efficacy of damping-off disease control by Pseudomonas fluorescens. Mol. Plant Microbe Interact. 2000, 13, 1293–1300. [Google Scholar] [CrossRef]
- Voisard, C.; Keel, C.; Haas, D.; Dèfago, G. Cyanide production by Pseudomonas fluorescens helps suppress black root rot of tobacco under gnotobiotic conditions. Embo J. 1989, 8, 351–358. [Google Scholar] [CrossRef]
- Keswani, C.; Singh, H.B.; García-Estrada, C.; Caradus, J.; He, Y.W.; Mezaache-Aichour, S.; Glare, T.R.; Borriss, R.; Sansinenea, E. Antimicrobial secondary metabolites from agriculturally important bacteria as next-generation pesticides. Appl. Microbiol. Biotechnol. 2020, 104, 1013–1034. [Google Scholar] [CrossRef] [PubMed]
- Duffy, B.K.; Defago, G. Environmental factors modulating antibiotic and siderophore biosynthesis by Pseudomonas fluorescens biocontrol strains. Appl. Environ. Microbiol. 1999, 65, 2429–2438. [Google Scholar] [CrossRef] [Green Version]
- Parret, A.H.; Temmerman, K.; De Mot, R. Novel lectin-like bacteriocins of biocontrol strain Pseudomonas fluorescens Pf-5. Appl. Environ. Microbiol. 2005, 71, 5197–5207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Principe, A.; Fernandez, M.; Torasso, M.; Godino, A.; Fischer, S. Effectiveness of tailocins produced by Pseudomonas fluorescens SF4c in controlling the bacterial-spot disease in tomatoes caused by Xanthomonas vesicatoria. Microbiol. Res. 2018, 212, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Uchino, M.; Shida, O.; Uchimura, T.; Komagata, K. Recharacterization of Pseudomonas fulva Iizuka and Komagata 1963, and proposals of Pseudomonas parafulva sp. nov. and Pseudomonas cremoricolorata sp. nov. J. Gen. Appl. Microbiol. 2001, 47, 247–261. [Google Scholar] [CrossRef] [Green Version]
- Guan-lin, X.; Algam, S.; Swings, J.; Mew, M.T. Diversity of Gram-negative bacteria antagonistic against major pathogens of rice from rice seed in the tropic environment. J. Zhejiang Univ. Sci. 2003, 4, 463–468. [Google Scholar]
- Mafia, R.G.; Alfenas, A.C.; Maffia, L.A.; Ferreira, E.M.; Mafia, G.M.V. Plant growth promoting rhizobacteria as agents in the biocontrol of eucalyptus mini-cutting rot. Trop. Plant Pathol. 2009, 34, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Zhang, Y.; Yu, N.; Bi, Z.; Zhu, A.; Zhan, X.; Wu, W.; Yu, P.; Chen, D.; Cheng, S.; et al. Genome sequence of Pseudomonas parafulva CRS01-1, an antagonistic bacterium isolated from rice field. J. Biotechnol. 2015, 206, 89–90. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, P.; Ye, G.; Lin, H.; Ren, D.; Guo, L.; Zhu, B.; Wang, Z. Complete genome sequence of Pseudomonas Parafulva PRS09-11288, a biocontrol strain produces the antibiotic phenazine-1-carboxylic Acid. Curr. Microbiol. 2019, 76, 1087–1091. [Google Scholar] [CrossRef]
- Jaaffar, A.K.M.; Parejko, J.A.; Paulitz, T.C.; Weller, D.M.; Thomashow, L.S. Sensitivity of Rhizoctonia isolates to phenazine-1-carboxylic acid and biological control by phenazine-producing Pseudomonas spp. Phytopathology 2017, 107, 692–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z. Research progress on antimicrobial mechanism and genetic engineering of Bacillus for plant diseases biocontrol. Acta Phytopathol. Sin. 2003, 33, 97–103. [Google Scholar]
- Tao, Y.M.; Pan, H.J.; Huang, J.; Xi, X.; Li, P.; Lu, Z.J. Research and application of a novel bio-control microbial factor Bacillus velezensis. China Plant Prot. 2019, 39, 26–33. (In Chinese) [Google Scholar]
- Ting, X.; Tianhui, Z.; Shujiang, L. β-1,3-1,4-glucanase gene from Bacillus velezensis ZJ20 exerts antifungal effect on plant pathogenic fungi. World J. Microb. Biot. 2016, 32, 26. [Google Scholar]
- Ye, M.; Tang, X.; Yang, R.; Zhang, H.; Li, F.; Tao, F.; Li, F.; Wang, Z. Characteristics and application of a novel species of Bacillus: Bacillus velezensis. ACS Chem. Biol. 2018, 13, 500–505. [Google Scholar] [CrossRef]
- Liu, Y.; Teng, K.; Wang, T.; Dong, E.; Zhang, M.; Tao, Y.; Zhong, J. Antimicrobial Bacillus velezensis HC6: Production of three kinds of lipopeptides and biocontrol potential in maize. J. Appl. Microbiol. 2020, 128, 242–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, B.; Wang, C.; Ding, X.; Zhu, B.; Song, X.; Borriss, R. AmyloWiki: An integrated database for Bacillus velezensis FZB42, the model strain for plant growth-promoting Bacilli. Database 2019, 2019, baz071. [Google Scholar] [PubMed]
- Pengfei, J.; Haonan, W.; Zheng, T.; Zhe, X.; Yaseen, D.G.; Li, Q.X.; Weiguo, M.; Wenbo, L. Antifungal mechanism of bacillomycin D from Bacillus velezensis HN-2 against Colletotrichum gloeosporioides Penz. Pestic. Biochem. Phys. 2020, 163, 102–107. [Google Scholar]
- Wu, L.M. Research about biocontrol activity of Bacillus amyloliquefaciens FZB42 and cyclodipeptides as a novel elicitor. Ph. D. Thesis, Nanjing Agricultural University, Nanjing, China, May 2016. (In Chinese). [Google Scholar]
- Gratia, J.P.; Crenier, L. Differential activity of bacteriocins and cefotaxime against Serratia marcescens clinical isolate SMG40 and its pigmented variant. Zent. FÜR Bakteriol. Int. J. Med Microbiol. 1992, 276, 340–346. [Google Scholar] [CrossRef]
- Laukova, A.; Marekova, M. Antimicrobial spectrum of bacteriocin-like substances produced by rumen staphylococci. Folia Microbiol. 1993, 38, 74–76. [Google Scholar] [CrossRef]
- Keswani, C.; Prakash, O.; Bharti, N.; Vílchez, J.I.; Sansinenea, E.; Lally, R.D.; Borriss, R.; Singh, S.P.; Gupta, V.K.; Fraceto, L.F.; et al. Re-addressing the biosafety issues of plant growth promoting rhizobacteria. Sci. Total Environ. 2019, 690, 841–852. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Host | Source or Reference | Antimicrobial Distance (mm) | Inhibition Rate a of 3–10 (%) | ||
|---|---|---|---|---|---|---|
| SC3 | SC11 | 3-10 | ||||
| Dickeya zeae EC1 | Rice | [28] | 16.64 ± 0.59 | 7.94 ± 0.49 | 9.86 ± 0.50 | / |
| D. zeae MS2 | Banana | [6] | 15.20 ± 0.23 | 8.89 ± 0.29 | 5.72 ± 0.24 | / |
| D. zeae MS3 | Banana | [6] | 16.48 ± 0.19 | 9.07 ± 0.13 | 6.59 ± 0.18 | / |
| D. fangzhongdai HK1 | Orchid | Lab storage | 12.59 ± 0.52 | 7.87 ± 0.34 | 3.76 ± 0.23 | / |
| D. dadantii 3937 | Potato | Lab storage | 3.66 ± 0.08 | 6.64 ± 0.21 | 4.24 ± 0.18 | / |
| Ralstonia solanacearum EP1 | Eggplant | Lab storage | 3.58 ± 0.14 | 7.44 ± 0.25 | 10.26 ± 0.27 | / |
| Xanthomonas campetris pv. campetris Xcc1 | Crucifer | [38] | 4.54 ± 0.17 | 0.86 ± 0.09 | 11.34 ± 0.83 | / |
| Pantoea anthophila CL1 | Wampee | [39] | 0 | 3.29 ± 0.19 | 4.23 ± 0.11 | / |
| Pa. ananatis PP1 | Peach | Lab storage | 0 | 1.12 ± 0.09 | 4.58 ± 0.18 | / |
| Pa. ananatis SC7 | Rice | Lab storage | 0 | 0 | 5.28 ± 0.10 | / |
| Colletotrichum capsici | Capsicum | Lab storage | 0 | 0 | 9.34 ± 0.20 | 48.50 ± 1.04 |
| C. gloeosporioides | Mango | Lab storage | 0 | 0 | 8.45 ± 0.23 | 42.10 ± 1.15 |
| Fusarium oxysporum f. sp. cubensis FOC4 | Banana | [40] | 0 | 0 | 7.30 ± 0.20 | 35.06 ± 0.99 |
| Rhizoctonia solani AG-1 1A | Rice | [41] | 0 | 0 | 3.64 ± 0.11 | 8.68 ± 0.28 |
| Magnaporthe oryzae B157 | Rice | [42] | 0 | 0 | 12.27 ± 0.23 | 80.18 ± 1.49 |
| Sporisorium scitamineum | Sugarcane | [36] | 0 | 0 | 17.04 ± 0.55 | 34.54 ± 1.11 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Hu, M.; Xue, Y.; Chen, X.; Lu, G.; Zhang, L.; Zhou, J. Screening, Identification and Efficacy Evaluation of Antagonistic Bacteria for Biocontrol of Soft Rot Disease Caused by Dickeya zeae. Microorganisms 2020, 8, 697. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8050697
Li J, Hu M, Xue Y, Chen X, Lu G, Zhang L, Zhou J. Screening, Identification and Efficacy Evaluation of Antagonistic Bacteria for Biocontrol of Soft Rot Disease Caused by Dickeya zeae. Microorganisms. 2020; 8(5):697. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8050697
Chicago/Turabian StyleLi, Jieling, Ming Hu, Yang Xue, Xia Chen, Guangtao Lu, Lianhui Zhang, and Jianuan Zhou. 2020. "Screening, Identification and Efficacy Evaluation of Antagonistic Bacteria for Biocontrol of Soft Rot Disease Caused by Dickeya zeae" Microorganisms 8, no. 5: 697. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8050697