RNA Interference by Cyanobacterial Feeding Demonstrates the SCSG1 Gene Is Essential for Ciliogenesis during Oral Apparatus Regeneration in Stentor

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Growth Conditions
2.2. RNAi Expression Vector Construction
2.3. Plasmid Construction
2.4. Synechocystis 6803 Transformation and Identification
2.5. Reverse Transcription-PCR Assay
2.6. RNAi by Feeding with Synechocystis 6803
2.7. Single-Cell RNA-Sequencing and Gene Expression Analysis
2.8. Quantitative Real-Time PCR Assay
2.9. Induction of OA Regeneration
2.10. Cell Shape Imaging and Analysis
3. Results and Discussion
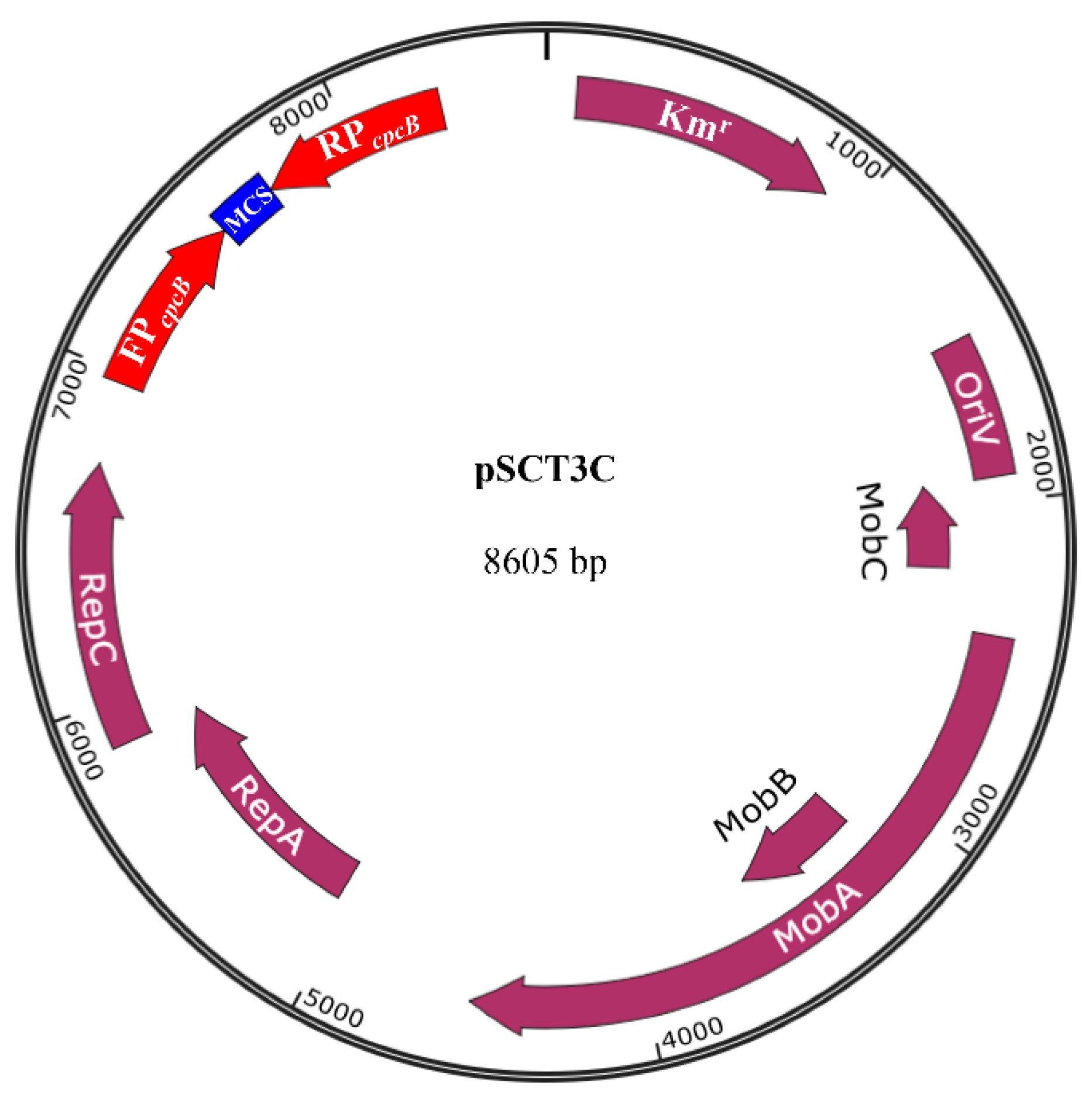
3.1. Structure of the pSCT3C RNAi Expression Vector
3.2. Synechocystis 6803 Transformation and Gene Expression Analysis
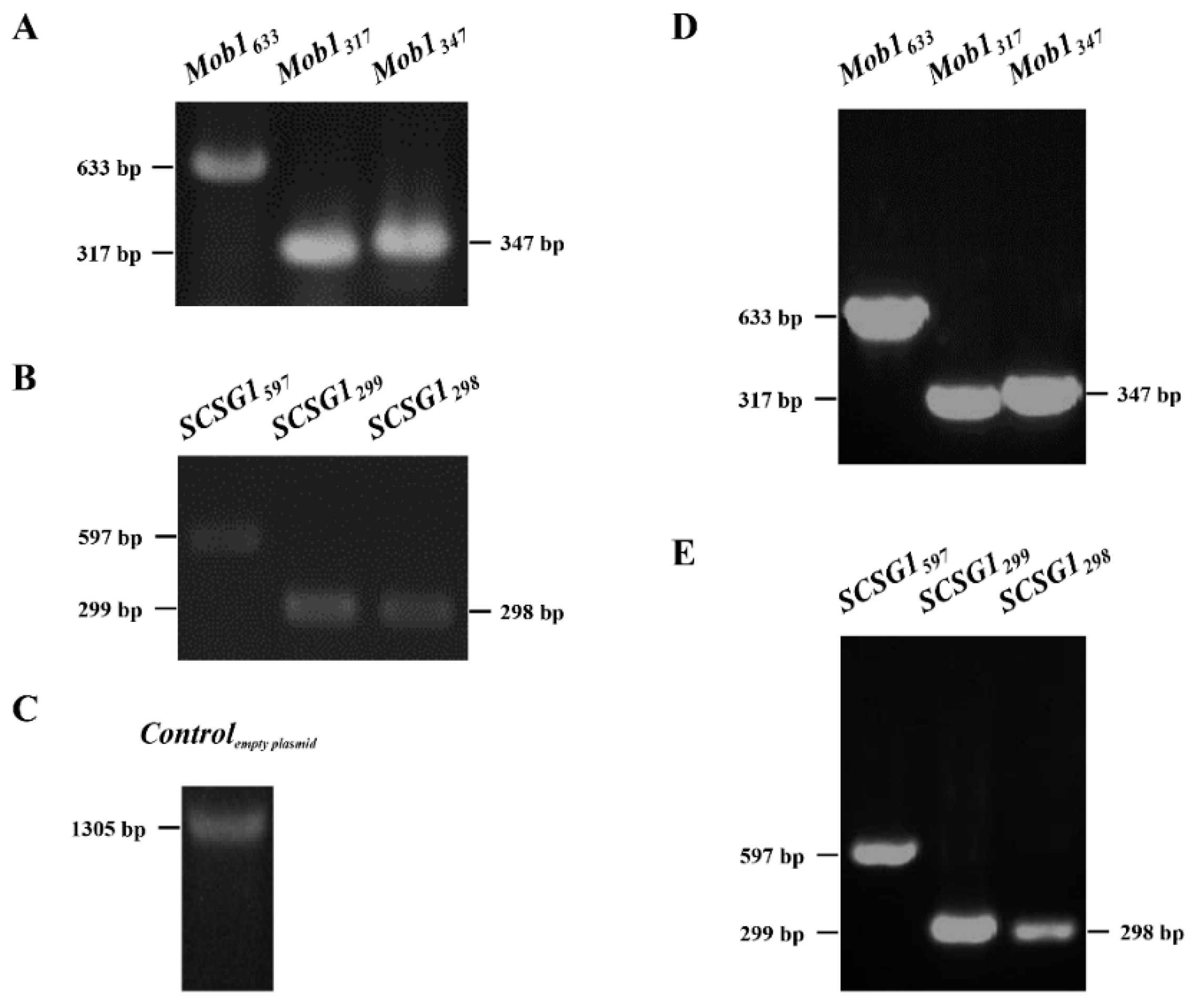
3.3. Successful Knockdown of Mob1 Supports the Applicability of RNAi by Synechocystis 6803 Feeding
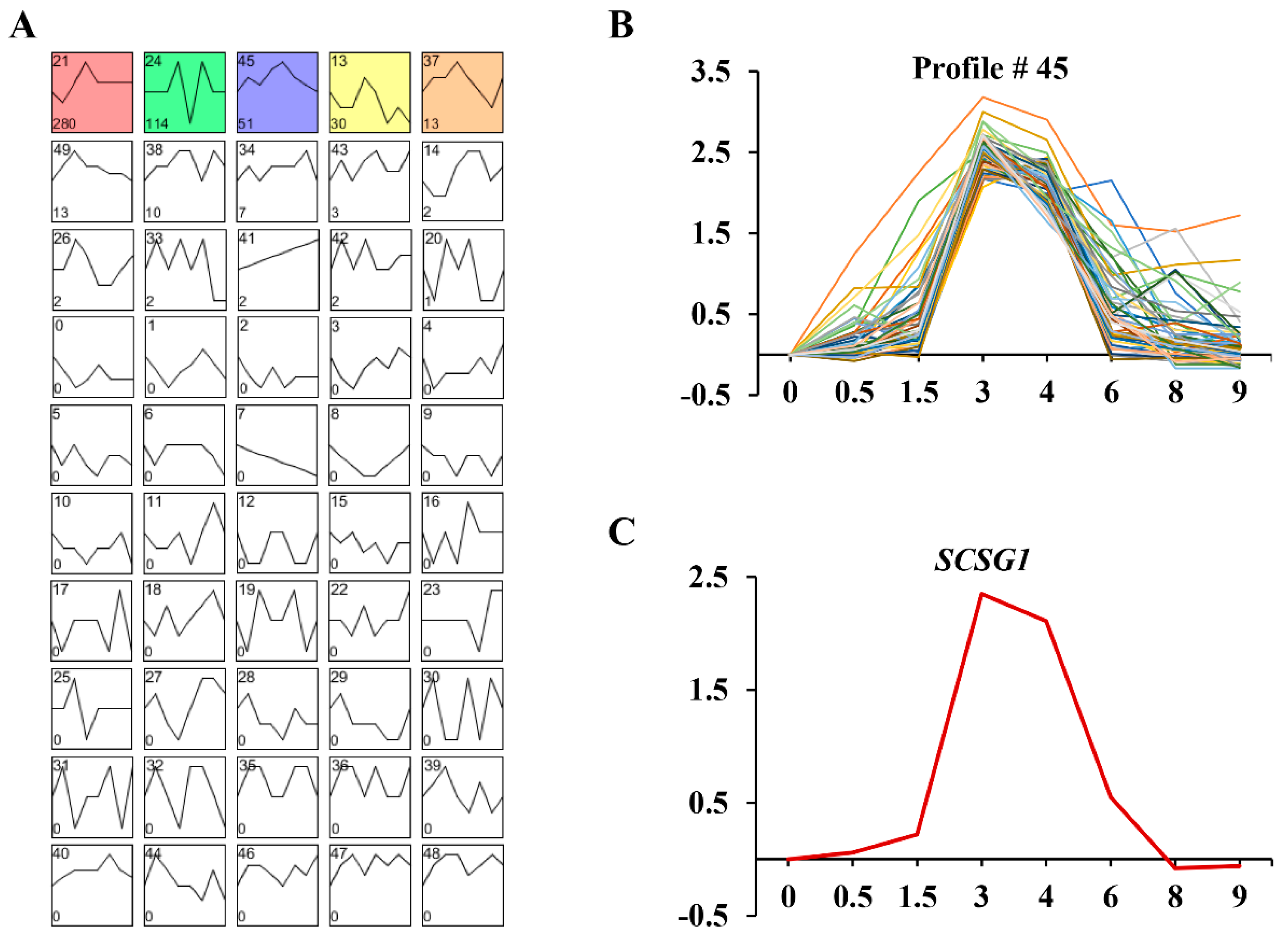
3.4. Gene Expression Profiles and Function Analysis of SCSG1 during OA Regeneration
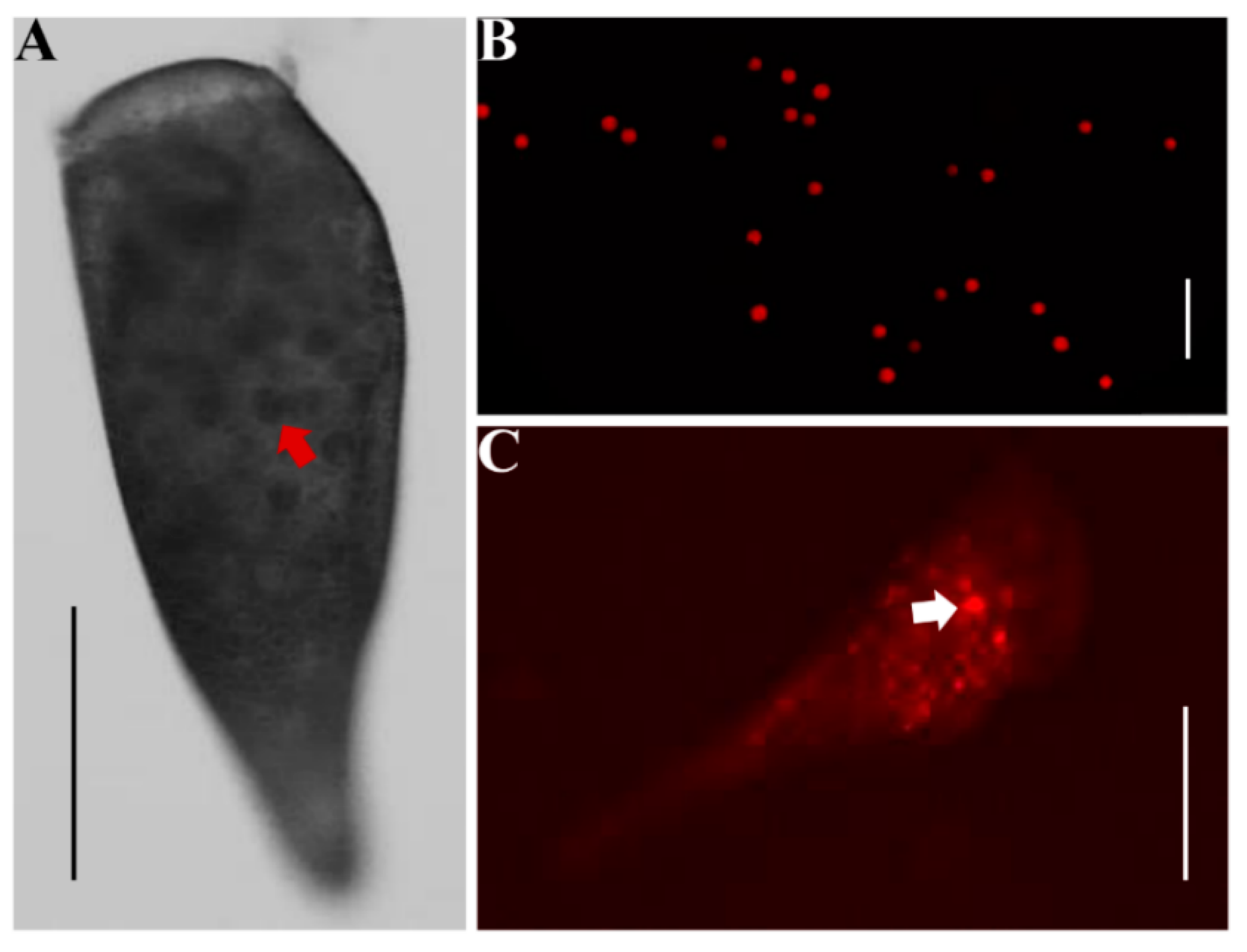
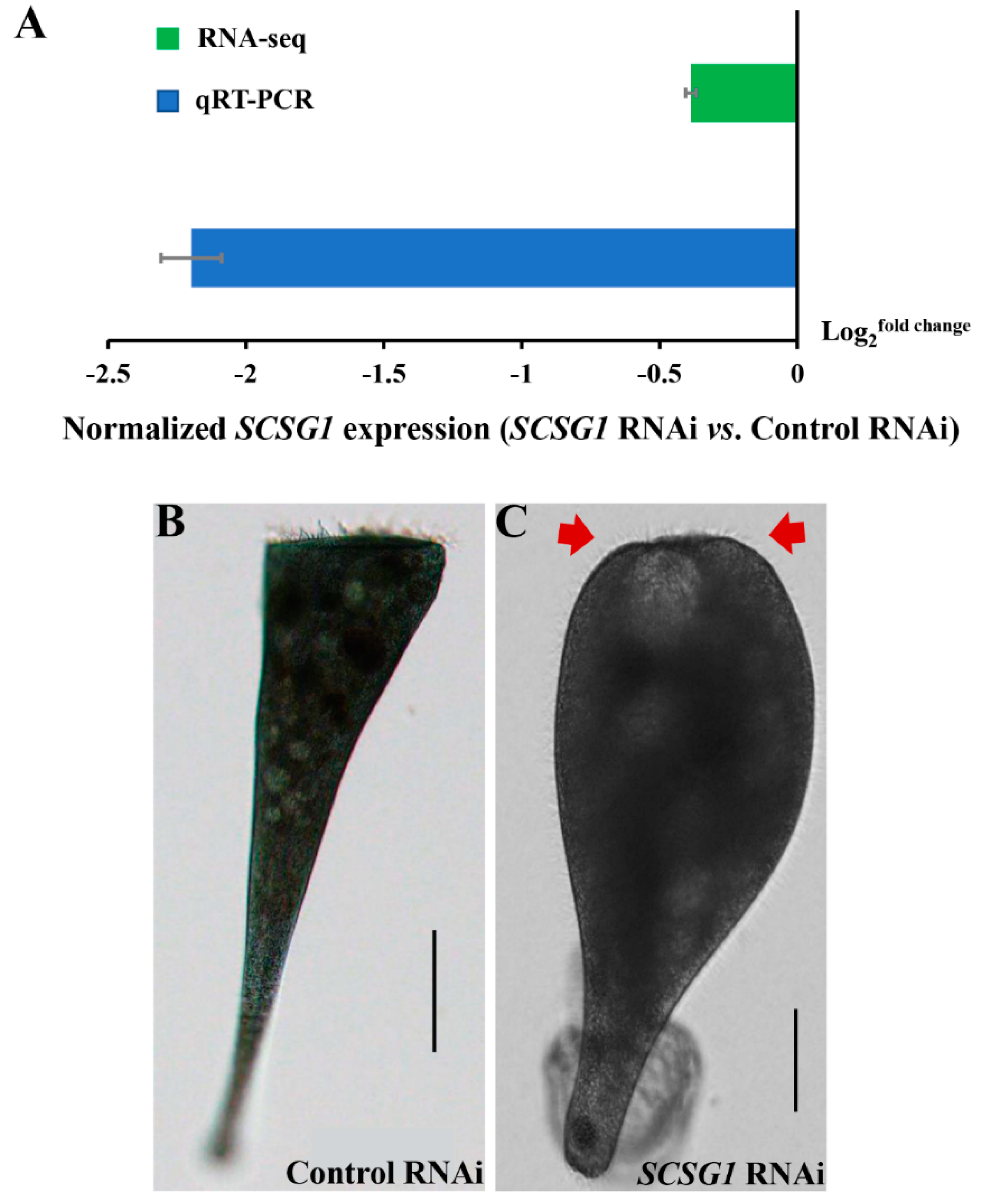
3.5. SCSG1 RNAi Illustrates Its Essential Function in OA Regeneration
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tartar, V. The Biology of Stentor; Pergammon Press: Oxford, UK, 1961. [Google Scholar]
- Neviackas, J.A.; Margulis, L. The effect of colchicine on regenerating membranellar cilia in Stentor coeruleus. J. Protozool. 1969, 16, 165–171. [Google Scholar] [CrossRef]
- Tartar, V. Reactions of Stentor coeruleus to certain substances added to the medium. Exp. Cell Res. 1957, 132, 317–332. [Google Scholar] [CrossRef]
- Sood, P.; McGillivary, R.; Marshall, W.F. The transcriptional program of regeneration in the giant single cell, Stentor coeruleus. Biorxiv Cell Biol. 2017, in press. [Google Scholar]
- Wei, W.; Jiang, C.Q.; Yang, W.T.; Miao, W.; Xiong, J. Proteomic identification and expression of oral apparatus constituents in cell regeneration of giant ciliate Stentor coeruleus (strain WHEL). Gene 2020, 743, 144624. [Google Scholar] [CrossRef] [PubMed]
- Galvani, A.; Sperling, L. RNA interference by feeding in Paramecium. Trends Genet. 2002, 18, 11–12. [Google Scholar] [CrossRef]
- Möllenbeck, M.; Postberg, J.; Paeschke, K.; Rossbach, M.; Jönsson, F.; Lipps, H.J. The telomerase-associated protein p43 is involved in anchoring telomerase in the nucleus. Cell Sci. 2003, 116, 1757–1761. [Google Scholar] [CrossRef] [Green Version]
- Nowacki, M.; Vijayan, V.; Zhou, Y.; Schotanus, K.; Doak, T.G.; Landweber, L.F. RNA-mediated epigenetic programming of a genome-rearrangement pathway. Nature 2008, 451, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Heyse, G.; Jönsson, F.; Chang, W.J.; Lipps, H.J. RNA-dependent control of gene amplification. Proc. Natl. Acad. Sci. USA 2010, 107, 22134–22139. [Google Scholar] [CrossRef] [Green Version]
- Kamath, R.S.; Martinez-Campos, M.; Zipperlen, P.; Fraser, A.G.; Ahringer, J. Effectiveness of specific RNA-mediated interference through ingested double-stranded RNA in Caenorhabditis elegans. Genome Biol. 2001, 2, 2. [Google Scholar]
- Slabodnick, M.M.; Ruby, J.G.; Dunn, J.G.; Feldman, J.L.; DeRisi, J.L.; Marshall, W.F. The kinase regulator Mob1 acts as a patterning protein for Stentor morphogenesis. PLoS Biol. 2014, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Konior, K. Tale of Two Proteins Sfi1p-Like and Centrin: Modeling of Contraction and Relaxation in V. convallaria. Ph.D. Thesis, University of Illinois at Chicago, Chicago, IL, USA, 2013. [Google Scholar]
- Yu, Y.; You, L.; Liu, D.; Hollinshead, W.; Zhang, F.Z. Development of Synechocystis sp. PCC 6803 as a phototrophic cell factory. Mar. Drugs 2013, 11, 2894–2916. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Sato, S.; Kotani, H.; Tanaka, A.; Asamizu, E.; Nakamura, Y.; Miyajima, N.; Hirosawa, M.; Sugiura, M.; Sasamoto, S.; et al. Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. II. sequence determination of the entire genome and assignment of potential protein-coding regions. DNA Res. 1996, 3, 109–136. [Google Scholar] [CrossRef] [Green Version]
- Marraccini, P.; Bulteau, S.; Cassier-Chauvat, C.; Mermet-Bouvier, P.; Chauvat, F. A conjugative plasmid vector for promoter analysis in several cyanobacteria of the genera Synechococcus and Synechocystis. Plant Mol. Biol. 1993, 23, 905–909. [Google Scholar] [CrossRef]
- Durfee, T.; Nelson, R.; Baldwin, S.; Plunkett, G.; Burland, V.; Mau, B.; Petrosino, J.F.; Qin, X.; Muzny, D.M.; Ayele, M.; et al. The complete genome sequence of Escherichia coli DH10B: Insights into the biology of a laboratory workhorse. Bacteriology 2008, 190, 2597–2606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varman, A.M.; Xiao, Y.; Pakrasi, H.B.; Tang, Y.J. Metabolic engineering of Synechocystis sp. strain PCC 6803 for isobutanol production. Appl. Environ. Microbiol. 2013, 79, 908–914. [Google Scholar] [CrossRef] [Green Version]
- Elhai, J.; Vepritskiy, A.; Muro-Pastor, A.M.; Flores, E.; Wolk, C.P. Reduction of conjugal transfer efficiency by three restriction activities of Anabaena sp. strain PCC 7120. Bacteriology 1997, 179, 1998–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidorn, T.; Camsund, D.; Huang, H.H.; Lindberg, P.; Oliveira, P.; Stensjö, K.; Lindblad, P. Synthetic biology in cyanobacteria: Engineering and analyzing novel functions. Methods Enzymol. 2011, 497, 539–579. [Google Scholar] [PubMed]
- Imashimizu, M.; Fujiwara, S.; Tanigawa, R.; Tanaka, K.; Hirokawa, T.; Nakajima, Y.; Higo, J.; Tsuzuki, M. Thymine at −5 is crucial for cpc promoter activity of Synechocystis sp. strain PCC 6714. Bacteriology 2003, 185, 6477–6480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeamton, W.; Dulsawat, S.; Laoteng, K.; Tanticharoen, M.; Cheevadhanarak, S. Phycocyanin promoter of Spirulina platensis controlling heterologous expression in cyanobacteria. J. Appl. Phycol. 2011, 23, 83–88. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, H.F.; Meng, H.K.; Zhu, Y.; Bao, G.H.; Zhang, Y.P.; Li, Y.; Ma, Y.H. Discovery of a super-strong promoter enables efficient production of heterologous proteins in cyanobacteria. Sci. Rep. 2014, 4, 4500. [Google Scholar] [CrossRef]
- Elhai, J.; Wolk, C.P. Conjugal transfer of DNA to cyanobacteria. Methods Enzymol. 1988, 167, 747–754. [Google Scholar] [PubMed]
- Scholz, P.; Haring, V.; Wittmann-Liebold, B.; Ashman, K.; Bagdasarian, M.; Scherzinger, E. Complete nucleotide sequence and gene organization of the broad-host-range plasmid RSF1010. Gene 1989, 75, 271–288. [Google Scholar] [CrossRef]
- Slabodnick, M.M.; Ruby, J.G.; Reiff, S.B.; Swart, E.C.; Gosai, S.; Prabakaran, S.; Witkowska, E.; Larue, G.E.; Fisher, S.; Freeman, R.M.; et al. The macronuclear genome of Stentor coeruleus reveals tiny introns in a giant cell. Curr. Biol. 2017, 27, 569–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, J.; Bar-Joseph, Z. STEM: A tool for the analysis of short time series gene expression data. BMC Bioinform. 2006, 7, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laligne, C.; Klotz, C.; Loubresse, N.G.; Lemullois, M.; Hori, M.; Laurent, F.X.; Papon, J.F.; Louis, B.; Cohen, J.; Koll, F. Bug22p, a conserved centrosomal/ciliary protein also present in higher plants, is required for an effective ciliary stroke in Paramecium. Eukaryot. Cell 2010, 9, 645–655. [Google Scholar] [CrossRef] [Green Version]
- Yanagisawa, H.A.; Mathis, G.; Oda, T.; Hirono, M.; Richey, E.A.; Ishikawa, H.; Marshall, W.F.; Kikkawa, M.; Qin, H. FAP20 is an inner junction protein of doublet microtubules essential for both the planar asymmetrical waveform and stability of flagella in Chlamydomonas. Mol. Biol. Cell 2014, 25, 1472–1483. [Google Scholar] [CrossRef]
- Maia, T.M.; Gogendeau, D.; Pennetier, C.; Janke, C.; Basto, R. Bug22 influences cilium morphology and the post-translational modification of ciliary microtubules. Biol. Open 2014, 3, 138–151. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Construct Name | Length (bp) | Target Region in CDS | Sample Size (Cell Number) | RNAi Efficiency |
|---|---|---|---|---|---|
| Mob1 CDS length: 675 bp | Mob1633 | 633 | 40–672 bp | 51 | 5.9% |
| Mob1317 | 317 | 40–356 bp | 34 | 5.9% | |
| Mob1347 | 347 | 326–672 bp | 40 | 0 | |
| SCSG1 CDS length: 597 bp | SCSG1597 | 597 | 1–597 bp | 43 | 0 |
| SCSG1299 | 299 | 1–299 bp | 58 | 5.2% | |
| SCSG1298 | 298 | 300–597 bp | 57 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, W.; Jiang, C.; Chai, X.; Zhang, J.; Zhang, C.-C.; Miao, W.; Xiong, J. RNA Interference by Cyanobacterial Feeding Demonstrates the SCSG1 Gene Is Essential for Ciliogenesis during Oral Apparatus Regeneration in Stentor. Microorganisms 2021, 9, 176. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010176
Wei W, Jiang C, Chai X, Zhang J, Zhang C-C, Miao W, Xiong J. RNA Interference by Cyanobacterial Feeding Demonstrates the SCSG1 Gene Is Essential for Ciliogenesis during Oral Apparatus Regeneration in Stentor. Microorganisms. 2021; 9(1):176. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010176
Chicago/Turabian StyleWei, Wei, Chuanqi Jiang, Xiaocui Chai, Juyuan Zhang, Cheng-Cai Zhang, Wei Miao, and Jie Xiong. 2021. "RNA Interference by Cyanobacterial Feeding Demonstrates the SCSG1 Gene Is Essential for Ciliogenesis during Oral Apparatus Regeneration in Stentor" Microorganisms 9, no. 1: 176. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010176