
Airport Malaria in Non-Endemic Areas: New Insights into Mosquito Vectors, Case Management and Major Challenges



1
Malaria Research Laboratory, Yaoundé Research Institute (IRY), Organization for the Coordination of Endemic Diseases’ Control in Central Africa (OCEAC), Yaoundé P.O. Box 288, Cameroon
2
Department of Biological Sciences, Faculty of Medicine and Pharmaceutical Sciences, University of Douala, Douala P.O. Box 2701, Cameroon
3
Department of Insect Biotechnology in Plant Protection, Institute for Insect Biotechnology, Faculty 09—Agricultural Sciences, Nutritional Sciences and Environmental Management, Justus-Liebig-University Gießen, Winchester Str. 2, 35394 Giessen, Germany
*
Author to whom correspondence should be addressed.
Microorganisms 2021, 9(10), 2160; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102160
Submission received: 3 September 2021
/
Revised: 11 October 2021
/
Accepted: 13 October 2021
/
Published: 16 October 2021
(This article belongs to the Special Issue Malaria in Non-Endemic Areas: Diagnosis, Therapy and Epidemiology)
Abstract
:Despite the implementation of preventive measures in airports and aircrafts, the risk of importing Plasmodium spp. infected mosquitoes is still present in malaria-free countries. Evidence suggests that mosquitoes have found a new alliance with the globalization of trade and climate change, leading to an upsurge of malaria parasite transmission around airports. The resulting locally acquired form of malaria is called Airport malaria. However, piecemeal information is available, regarding its epidemiological and entomological patterns, as well as the challenges in the diagnosis, treatment, and prevention. Understanding these issues is a critical step towards a better implementation of control strategies. To cross reference this information, we conducted a systematic review on 135 research articles published between 1969 (when the first cases of malaria in airports were reported) and 2020 (i.e., 51 years later). It appears that the risk of malaria transmission by local mosquito vectors in so called malaria-free countries is not zero; this risk is more likely to be fostered by infected vectors coming from endemic countries by air or by sea. Furthermore, there is ample evidence that airport malaria is increasing in these countries. From 2010 to 2020, the number of cases in Europe was 7.4 times higher than that recorded during the 2000–2009 decade. This increase may be associated with climate change, increased international trade, the decline of aircraft disinsection, as well as delays in case diagnosis and treatment. More critically, current interventions are weakened by biological and operational challenges, such as drug resistance in malaria parasites and vector resistance to insecticides, and logistic constraints. Therefore, there is a need to strengthen malaria prevention and treatment for people at risk of airport malaria, and implement a rigorous routine entomological and epidemiological surveillance in and around airports.
1. Introduction
Malaria is a parasitic disease transmitted by the Anopheles mosquito species, to which 3.5 billion people are exposed worldwide [1]. In 2019, 229 million new cases were recorded and the disease is endemic in 87 countries [1]. Although it has been eliminated in the United States, Canada, and Europe, more and more cases are reported in these regions, mostly imported cases [2]. Overall, it is estimated that over 30,000 annual cases of imported malaria occur among the travelers from endemic countries [3]. The majority of these travelers arrive from sub-Saharan Africa (97.5%), mainly the native Africans residing in Europe (82.2%) [4,5]. Indeed, malaria is a real emergency in these areas because it can rapidly become fatal if not diagnosed and treated promptly.
The epidemiological patterns of imported malaria are closely related to the travels of Anopheles mosquito vectors and humans infected with Plasmodium spp. parasites [6]. When local environmental conditions at the destinations are favorable for the development of Anopheles species (i.e., 27 ± 2 °C temperature and 70 ± 5% relative humidity), those which are efficient malaria vectors and capable of breeding near airports can cause local transmission of Plasmodium spp. parasites after blood feeding on infected travelers [5]. In addition, malaria parasite transmission in the vicinity of airports can result from infected Anopheles mosquitoes brought by aircraft, that can survive long haul flights and adapt for enough time in the new environment after arrival [7]. The resulting autochthonous (locally-acquired) malaria, in which cases are principally clustered around international airports, is known as “airport malaria” [8]. The patients had neither recently traveled to endemic areas nor recently received blood transfusions [9]. This form of malaria is increasingly being reported in Europe and North American countries that are considered malaria-free [9,10].
During the early 1930s, the World Health Organization (WHO) was already thinking about how to disinsectize airplanes in order to prevent airport malaria. Currently, the method recommended by WHO is disinsection by spraying a pyrethroid insecticide in the cabins and holds of aircraft flying from endemic areas [11,12]. However, mosquitoes in passenger luggage and cargo can escape this insecticide and survive during the flight, more importantly if they have developed resistance to pyrethroid insecticides [13]. Despite the implementation of prevention measures, the risk of importing infected mosquito vectors has increased with the intensification of air transport, as well as the size and complexity of today’s aircraft, with a multitude of niches that escape insecticide treatment. However, little research has been conducted to better understand the transmission of malaria at airports and around. Furthermore, piecemeal information is available, regarding the evolution of epidemiological patterns, as well as the challenges in its diagnosis, treatment, and prevention. Understanding these issues is a critical step towards a better implementation of control strategies. The purpose of this study is to draw the attention of researchers and policy makers to the concerns about airport malaria in non-endemic areas.
In this paper, we provide new insights into the factors that might lead to the re-emergence of airport malaria and potential actions to be taken in order to strengthen the strategies for preventing the reintroduction of malaria in malaria-free regions.
2. Data Retrieval
We conducted a systematic review of the literature published from 1969 (when the first cases of airport malaria were reported) to 2020 (51 years later), in four main databases, including Hinari Health, PubMed, Google and Google Scholar. Priority was granted to documents from the “Journal Article” category and other papers highlighting at least one of the following terms: “airport”, “malaria”, “prevention”, “Plasmodium spp.”, “Anopheles” “mosquitoes”, “non-endemic areas” in their titles, abstracts, or keywords.
Advanced searches conducted in databases were based on search equations, which represent associations of at least two of the selected terms. The research equations were formulated in English and French.
For the selected articles, full texts were obtained and examined in-depth to explore how airport malaria in non-endemic areas was documented. From each of the included articles in this review, information and data on the following topics related to airport malaria were extracted when available:
- -
- Vector bionomics, incrimination, and importation from endemic areas;
- -
- Airport malaria epidemiological profiles;
- -
- Facilities and tools for diagnosis of Plasmodium spp. infections;
- -
- Treatment and management of malaria cases;
- -
- Airport malaria mass prevention strategies.
Epidemiological data were entered into a Microsoft Excel spreadsheet for easy access and data analysis. We also collected information on the study locations and grouped them into regions according to the United Nations designation of areas and regions [14]. Furthermore, we paid particular attention to the challenges encountered in the implementation of prevention and control strategies.
3. Results
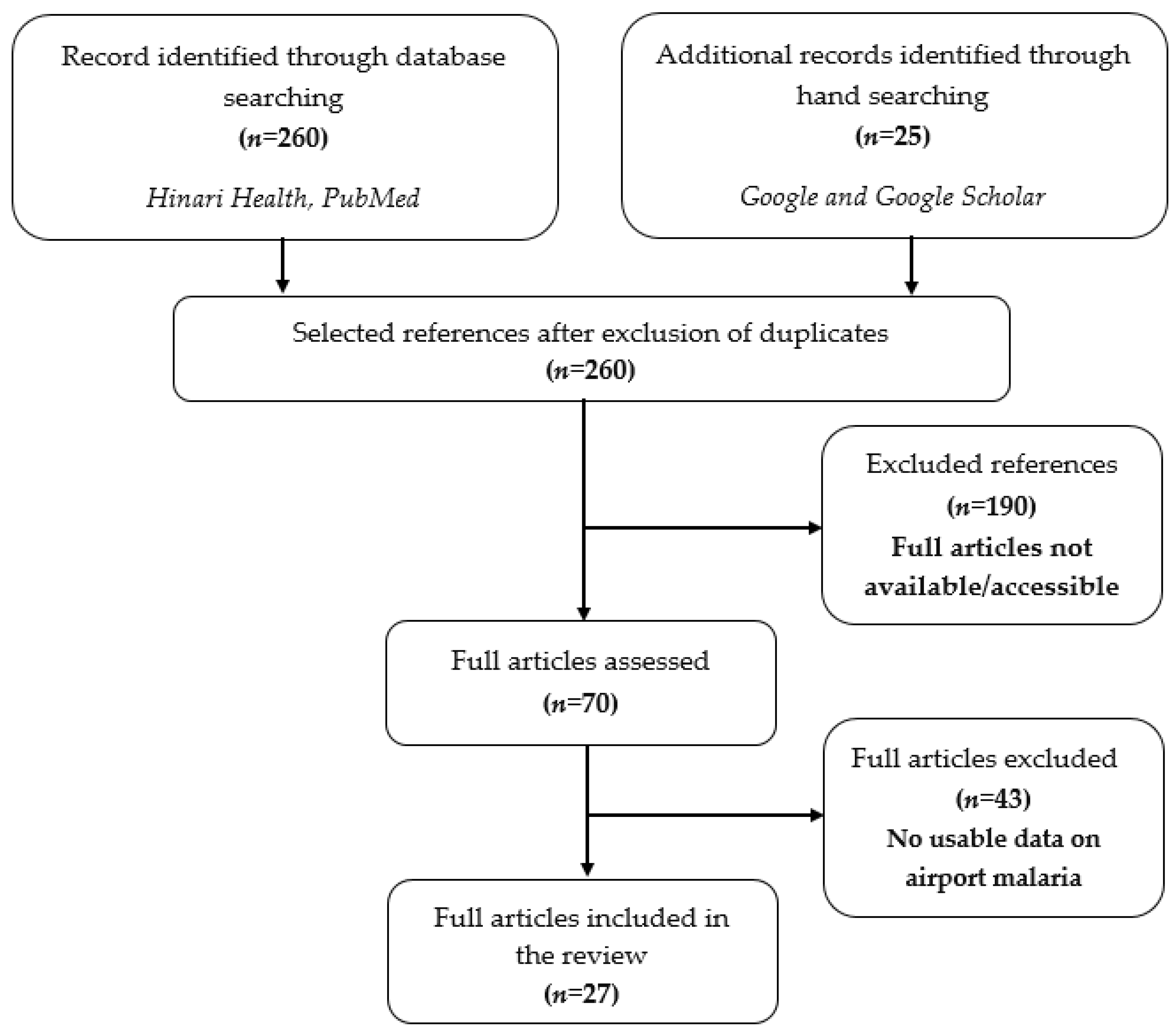
The search resulted in 285 published articles from which more than 235 articles were excluded because they were not related to airport malaria or did not report data from non-endemic areas, as shown in Figure 1.
The 27 papers that were included in this work were those in which malaria cases in patients were associated with Anopheles transportation by aircraft or ships.
Of the 27 published papers, 52% (14/27) were specific to Europe, 37% (14/27) were multiregional, and 11% (3/27) for other countries (Americas, Tunisia, Turkmenistan). A total of 33 studies were recorded in the 27 selected articles. Among these studies, 33.3% (11/33) were about vector bionomics, incrimination, and importation from endemic areas, 21.2% (7/33) about malaria epidemiological profiles, 15.2% (5/33) about facilities and tools for diagnosis of Plasmodium spp. infections, 9.1% (3/33) about treatment and management of cases, and 21.2% (7/33) about mass prevention strategies, as shown in Table 1.
3.1. Mosquito Vectors
3.1.1. Vector Bionomics and Incrimination
In Europe and the Mediterranean region, malaria elimination was achieved by the middle 20th century [41,42]. Anopheles sacharovi, An. atroparvus, An. artemievi, An. beklemishevi, An. daciae, An. labranchiae, An. maculipennis, An. martinus, An. melanoon, An. messeae, and An. persiensis were the main vectors of Plasmodium spp. parasites due to their anthropophilic behavior [17]. These 11 species are members of the An. maculipennis complex, and they are still widely distributed throughout the European Region [18]. Four of these 11 species, An. sacharovi, An. atroparvus, An. labranchiae and An. messeae are considered dominant vectors of malaria [19].
Anopheles sacharovi
Anopheles sacharovi has the most southerly distribution owing to its high adaptive capacity at adult and larval stages. It can breed in different types of water bodies, such as swamps, marshes, riverbanks, streams, ponds, and ditches. It is the most efficient mosquito vector of Plasmodium spp. parasites in southern Europe and the Middle East [18,19]. Currently, this species is involved in the transmission of P. vivax malaria in Iran [20,21], Iraq [43], and Turkey [44]. It is also responsible for the reemergence of malaria in Georgia, Armenia, and Azerbaijan [18]. Female mosquitoes have opportunistic blood feeding behavior and can feed on any available host, including man, cow, sheep, chicken, horse, and donkey [18]. Anopheles sacharovi has been found resistant to DDT [21] and dieldrin [45]. Such ecological and behavioral plasticity, together with emerging insecticide resistance, has prevented the elimination of An. sacharovi in Israel, Greece, and Turkey [18,44].
Anopheles atroparvus
Anopheles atroparvus is present in Eastern and Central Europe, as well as in the United Kingdom [22,46]. Its absence has been reported in Greece, Turkey [47], and southern Italy [22]. Although described as a species with a preference for brackish larval habitats [23], this species has been found in a number of freshwater habitats, including canals, ditches, riverbanks, pools in riverbeds, and rice paddies [19]. Anopheles atroparvus has been described as an endophilic species, mostly endophagous and zoophilic [19]. Its hosts, in order of preference, are the rabbit, horse, cow, pig, and sheep [48,49,50,51]. Because of its association with human settlements, An. atroparvus also exhibits anthropophilic behavior [19]. Historically, An. atroparvus was involved in the transmission of local strains of P. vivax [52] and P. falciparum in Spain [22]. Studies have shown that this vector species is refractory to Asian and African P. falciparum [19,22]. De Zulueta et al. [53] and Cambournac [54] though that the refractory character of An. atroparvus with African and eastern P. falciparum strains is a “fact undeniable”. However, Sousa [55], under laboratory conditions, succeeded in infecting An. atroparvus with tropical strains of P. falciparum. Thus, An. atroparvus can be infected with tropical strains of P. falciparum, although its transmission would unlikely occur under natural conditions.
Anopheles labranchiae
Anopheles labranchiae is widely present in central Italy despite the eradication campaign launched in 1946 [56]. In general, its larvae are found in stagnant or low flowing water [57] and rice fields [58,59]. Females can aggressively attack human hosts and are de-scribed as “persistent” in their attempt to enter rooms during the night [59]. Although de-scribed as zoophilic, this species is opportunistic in its choice of host; it can alternatively bite humans or animals [58,60,61]. As with An. atroparvus, some studies have shown that An. labranchiae is refractory to tropical strains of P. falciparum [53]. However, Toty et al. [62] reported historical evidence, as well as the results of a contemporary study conducted by the Centre de Production et d’Infection d’Anophèles (CEPIA) in Paris where 14% (13/99) of specimens of An. labranchiae were experimentally infected with the laboratory-grown African strain of P. falciparum. This study also detected sporozoites in the salivary glands of three specimens, indicating that An. labranchiae is not only susceptible but also potentially capable of transmitting at least some strains of African P. falciparum.
Anopheles messeae
Anopheles messeae is the most widespread species of the An. maculipennis complex [23]. Its distribution extends from Europe to Asia passing through Russia [63]. Di Luca et al. [59] identified a number of genetic polymorphisms within An. messeae and defined five distinct haplotypes associated with different geographical areas. However, they could not confirm whether this polymorphism was associated with behavioral variability at different places. In surveys conducted in Slovenia and Croatia [64,65], the authors found adult An. messeae larvae in swamps, stagnant freshwater marshes, alluvial plains, and at sites near large lakes. Takken et al. [66] also inferred the presence of An. messeae in more brackish habitats. In urban areas, females have been found inside human dwellings [67]. Considering its high degree of zoophily and outdoor biting behavior, some authors thought it is very unlikely that this mosquito species is involved in local malaria transmission [66,68]. In contrast, Fyodorova et al. [67] found that 40% of female An. messeae collected in urban areas contained human blood, with the remaining 60% having fed on cats (40%) and chickens (20%). Becker et al. [69] stated that “blood meals are taken from humans only when the density of An. messeae is very high and there is a shortage of livestock”. Some evidence suggests that An. messeae may be refractory (or essentially refractory) to tropical strains of P. falciparum [70]. Indeed, the status of An. messeae as a malaria vector has been questioned, especially since the discovery of a new species in 2004, named An. daciae, widely distributed in Romania, southwest England, and parts of southern and central Finland [19,71]. Anopheles daciae can only be distinguished from An. messeae using egg morphology or by sequencing the internal transcribed spacer 2 (ITS2) of ribosomal DNA and the cytochrome oxidase 1 (COI) of mitochondrial DNA [19]. It is therefore possible that An. daciae, and not An. messeae is involved in malaria transmission.
3.1.2. Suspected Vectors
In addition to proven malaria vectors, there are also other vectors of the Hyrcanus group that may play distinct roles in the transmission of malaria pathogens, in particular P. vivax [72]. The Hyrcanus group includes approximately 30 closely-related species of mosquitoes distributed in the Palearctic and Eastern regions [73]. Among these 30 species, An. sinensis and An. lesteri are the main vectors of malaria in China [74]. Anopheles hyrcanus s.s. is a potential malaria vector in southern France and central Europe [48]. Anopheles kleini and An. pullus are the main vectors of malaria in the Republic of Korea [75], while An. sinensis, An. nigerrimus and An. peditaeniatus are potential malaria vectors in Thailand [76,77]. Within this group, there is some confusion as to the taxonomic status of An. hyrcanus s.s. and An. pseudopictus [15,16]. They were previously considered synonymous species. However, recent studies and identification keys refer to their taxa either as distinct species or as synonymous species. According to Miaoulis et al. [16], there is no strong evidence for treating these taxa as different species, so they concluded that An. hyrcanus s.s. and An. pseudopictus should be reclassified to their former synonym status.
3.1.3. Vector Importation
In 1930, shortly after the intensification of steamship traffic between the African and South American continents, An. arabiensis, a major malaria vector in Africa, was observed in the state of Natal in Brazil [24]. In 1994, 2000–5000 Anopheles mosquitoes were estimated to be imported in France within a week, i.e., 8–20 Anopheles specimens per flight [9]. In 2013, the presence of an Asian anopheline species (An. stephensi) was discovered in urban areas to Djibouti, and in subsequent years in Ethiopia [78]. More recently, in 2017 the mosquito species An. bancroftii was reported to be introduced in New Caledonia in an area nearby the airport of Noumea. New Caledonia is a French island in the southern Pacific Ocean which had been free of Anopheles mosquito species vectors of human Plasmodium spp., making this island free of malaria transmission [79,80]. The investigations suggest that the importation of this mosquito species was made by aircraft, although the route of this introduction was not clearly identified [81].
Since 1969, locally transmitted malaria cases (acquired on the European continent) have been associated with infected anophelines from endemic areas. Germany, the Netherlands, Spain, France, Italy, Greece, and Turkey have reported large numbers of cases [13]. Studies indicate that during the summer months (May-September), it is likely that a tropical mosquito such as An. gambiae can successfully establish itself in temperate regions of Europe or North America [7,25].
3.2. Airport Malaria Epidemiological Profiles
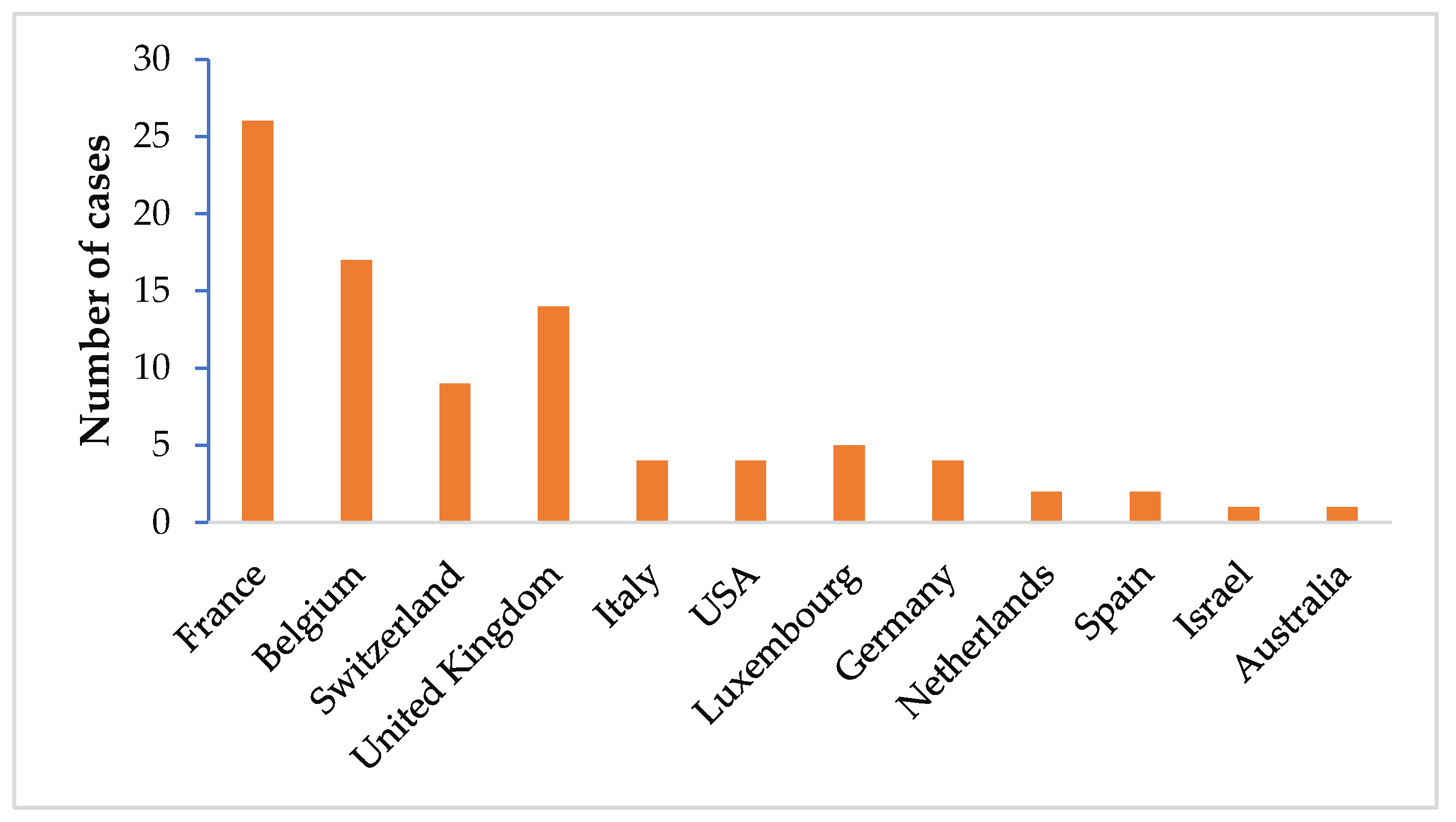
The global epidemiological data reported in 2006 showed that countries mostly affected by airport malaria are in Europe, particularly France (29.2%), Belgium (19.1%) and Switzerland (10.1%), Luxembourg (5.6%), Italy and Germany (4.5%), followed by the UK (15.7%), USA (4.5%), Israel and Australia (1.1%) (Figure 2) [6]. Some cases have also been reported in Tunisia [82]. In the meantime, this epidemiological situation may be changed, with the intensification of international trade.
In France in 2008, a man and his wife living in Paris and who never travelled to a malaria endemic area, were hospitalized at the University Hospital Central (CHU) of Nice for uncomplicated P. falciparum malaria. These people were living in Saint Mard (a town located 7 km from Roissy Charles de Gaulle Airport) [33]. In 2013, four cases of airport malaria due to P. falciparum were notified for the first time in Tunisia; all these patients were neighbors living within 2 km around the Tunis International Airport [82]. In Germany in 2019, two employees of Frankfurt International Airport, working in aircraft maintenance were hospitalized with severe P. falciparum malaria at the University Hospital Frankfurt [31]. The airplanes under maintenance were returning from three P. falciparum malaria endemic countries in sub-Saharan Africa. Similar cases of airport malaria due to P. falciparum have been reported in other countries. Furthermore, there has been an increasing number of human P. knowlesi malaria cases imported from Southeast Asia to other areas across the world [83]. Plasmodium knowlesi is a malaria parasite species commonly found in primates, which is now spreading in the human population in Asia. Indeed, numerous studies have shown an increase in the incidence of malaria due to this species among people returning from countries such as Indonesia, Singapore, the Philippines, Vietnam, Thailand, and Malaysia, where malaria has considerably increased during the last decade [84,85]. Over time, environmental changes, particularly those associated with extensive deforestation and global warming, may have resulted in an increase of the densities of zoophilic mosquitoes and their ability to feed on human hosts, thereby fostering the transmission of P. knowlesi to humans [83]. The rapid onset P. knowlesi parasite species in humans is therefore considered a leading factor for the upsurge of airport malaria.
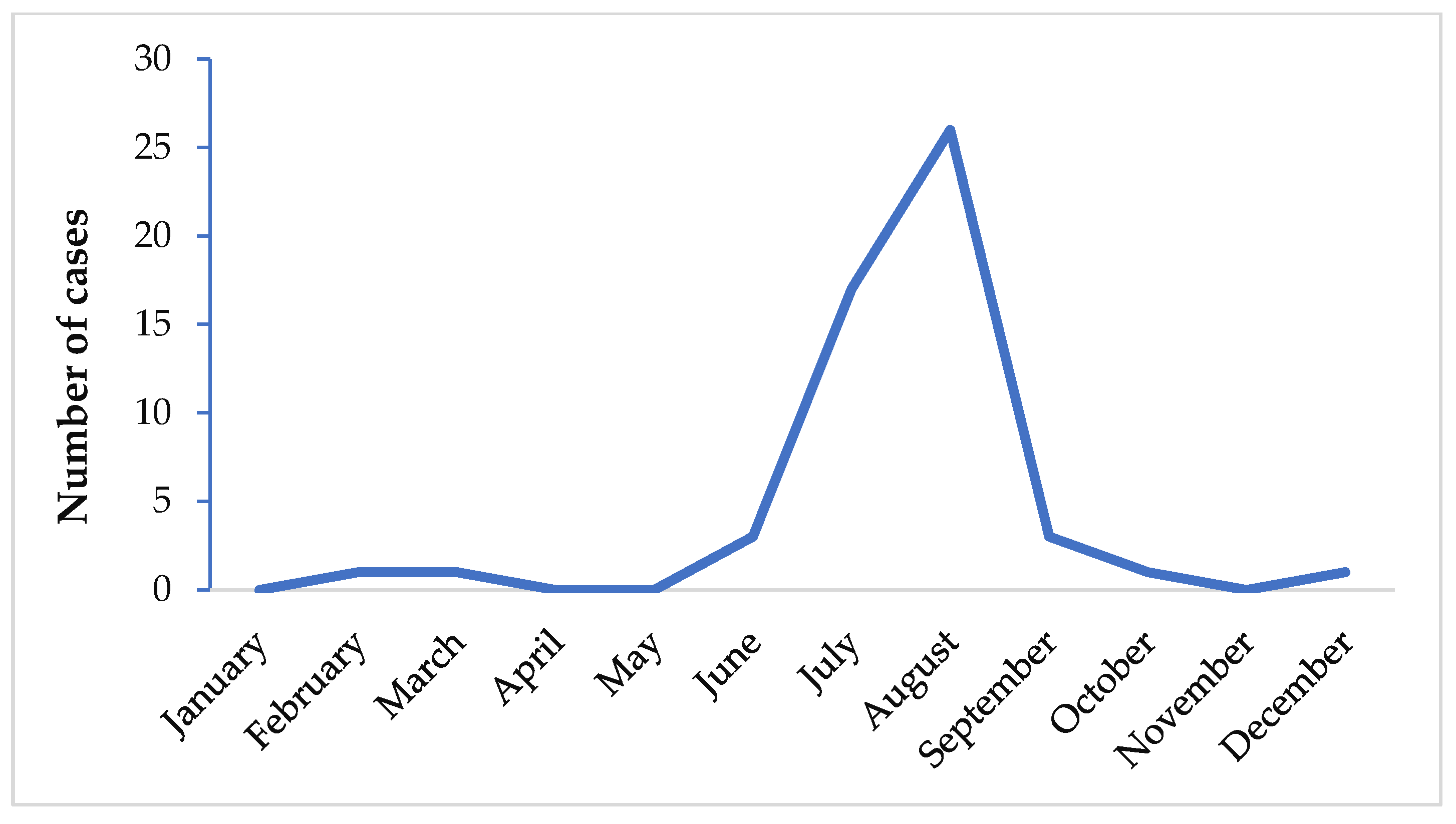
It has been shown that the greatest number of airport malaria cases occurs in July and August (Figure 3) [6,25]. At that period of the year, the climate is similar to that of malaria endemic countries and therefore suitable for Anopheles spp. survival at their arrival, with subsequent development and transmission of Plasmodium spp. in non-endemic areas.
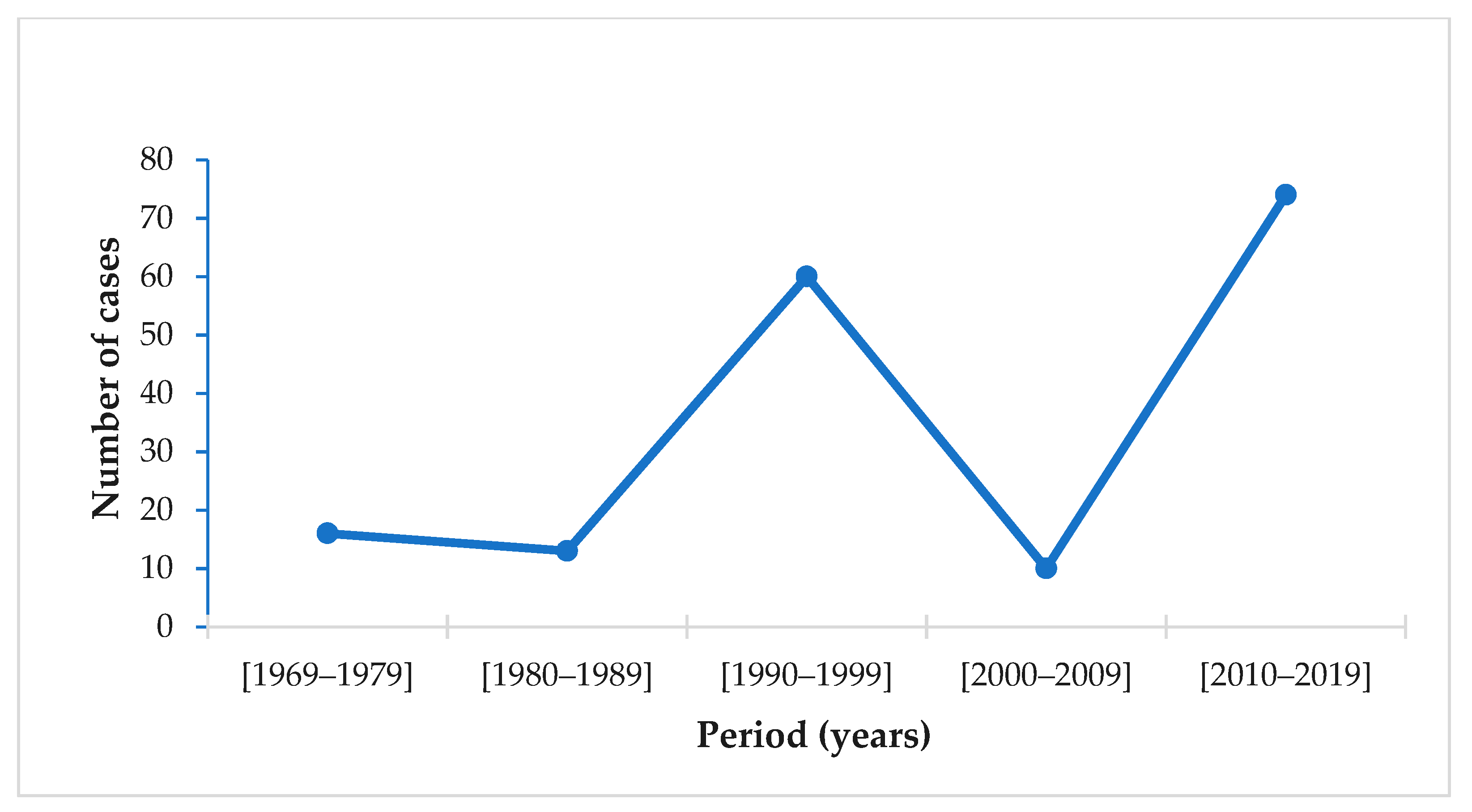
The evolution of the number of cases in Europe from 1969 to 2020 reveals an increase in the number of cases during the last decade (Figure 4). Among the 29 European Union countries reporting malaria data between 2010 and 2020, 11 reported airport malaria cases [25,28]. According to WHO, airport malaria cases have been on the rise since 2015 [42]. In France, mortality due to airport malaria is 20 times higher than that of imported malaria [24].
3.3. Facilities and Tools for Diagnosis of Plasmodium spp. Infections
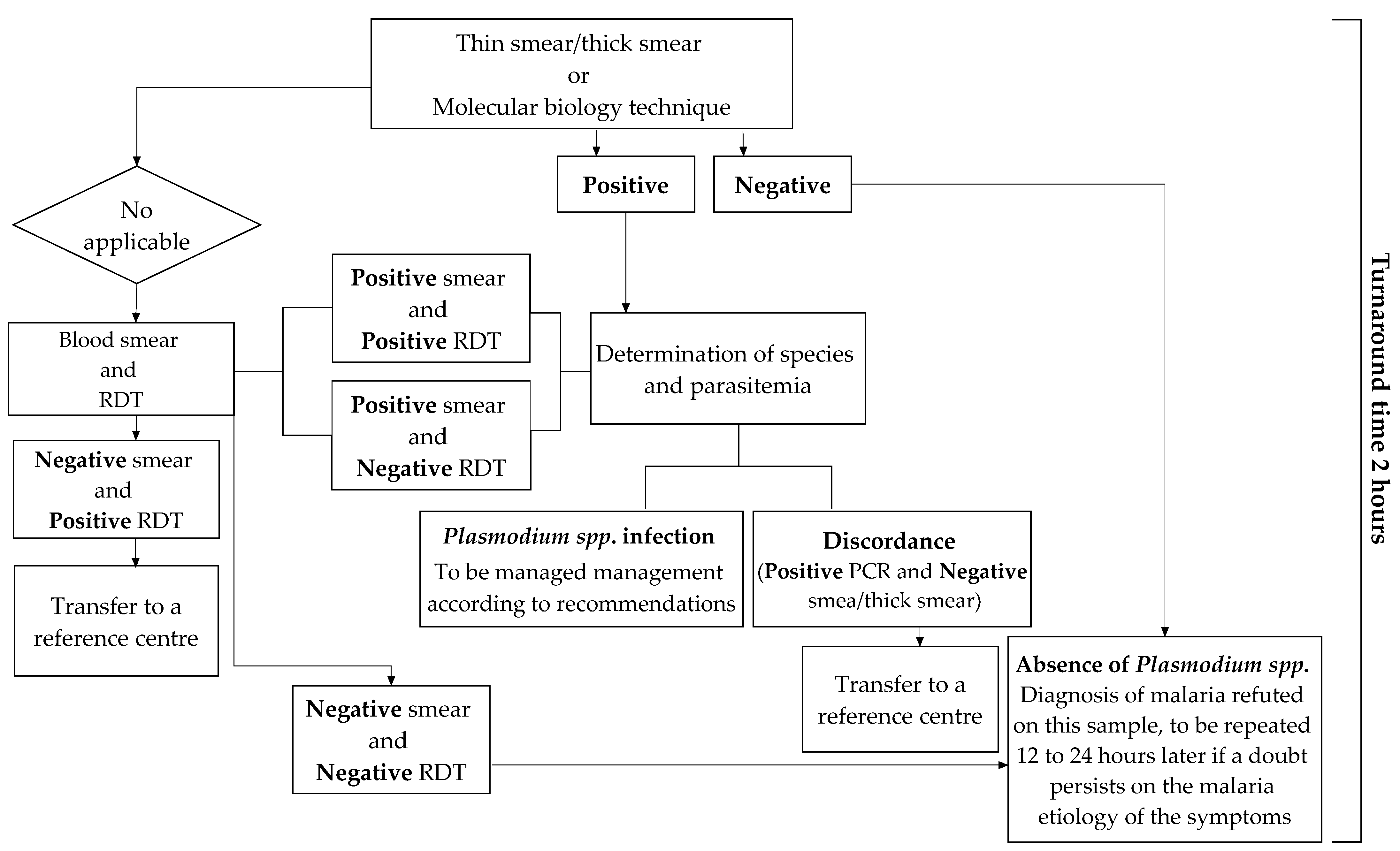
The diagnostic tools that can be used in the laboratory differ in their technologies, sensitivities, interpretations, and costs. The use of either can be prioritized according to the technical resources available and the biologist’s experience in detecting Plasmodium spp. parasites, as shown Figure 5 [35].
The 12th consensus conference on clinical practice organized by the Society of Infectious Pathology of French Language (SIPFL) in 1999 and revised in 2008 recalled that the biological diagnosis of malaria should be microscopic [89]. It is the method that objectively determines the presence of the parasites. This examination must allow to obtain results in less than two hours. However, in airport or imported malaria, low parasitemia often leads to performing a concentration technique (thick drop) or QBC™ Malaria Test as the first line [33]. The QBC™ Malaria Test is the fastest and most sensitive technique. Its extra cost compared to the thick drop is largely balanced with time savings. Still, the blood smear remains essential to determine Plasmodium spp. [24]. These techniques must therefore be perfectly known and mastered by the medical professionals in place; therefore, regular hands-on training is essential for them. However, evaluations conducted in France have shown that nearly 30% of laboratories returned their diagnosis results in more than two hours [33]; furthermore 16.5% of the results were inaccurate [90]. To overcome these challenges, there is a renewed interest in non-microscopic techniques.
Rapid Diagnostic Tests are the most accessible and least expensive; their ease of use makes them of valuable aid [33]. They are highly used by laboratories that rarely see positive blood samples and by biologists or technicians on-call [91]. It should be recalled, however, that this test is less sensitive (e.g., 10 fold less) than the concentration methods; so, its negativity does not make it possible to rule out the diagnosis in the case of very low parasitemia [33]. This is true for P. falciparum and even more so for other species [34,92].
Molecular biology techniques offer the advantage of a very high sensitivity. Most of the time, they are used as confirmatory tests in cases where the interpretation of the thick drop is difficult [33,34]. Venous blood (300 µL) is applied to a filtered paper FTA (Flinders Technology Associates) microcard, which is then stored in an aluminum barrier bag until the PCR (Polymerase Chain Reaction) test is performed [93]. The structure and composition of the FTA blotting paper allows for automatic isolation of DNA from its surface; the DNA is ready to be purified and analyzed after 30 min [93]. However, due to their implementation constraints, these techniques can only be used in equipped centers and during regular working hours. Indeed, they may not be available on call; therefore, their utilization is often delayed.
The diagnosis of Plasmodium spp. in mosquitoes can be determined by three main methods:
- -
- Microscopically examining the salivary glands of mosquitoes after dissection for direct observation of Plasmodium spp. sporozoites;
- -
- The circum-sporozoite antigen enzyme-linked immunosorbent assay (CSP-ELISA);
- -
- Analysis of genomic markers using polymerase chain reaction (PCR).
These methods allow the determination of the sporozoite index (SPI) of Anopheles and the entomological inoculation rate (EIR) [94], which is the number of infective mosquito bites per human per night. Although the first two methods can routinely be used in specialized laboratories of entomology, they are known to be labour intensive and it has been shown that CSP-ELISA [95] overestimates the real infection rate by detecting the CSP from the oocysts bursting 2–3 days before the sporozoites actually reach the salivary glands [96,97]. Research efforts in recent decades have led to the development of molecular biology tools for detecting the five major species of Plasmodium (P. falciparum, P. malariae, P. ovale, P. vivax and P. knowlesi) in human blood [98,99] and in Anopheles mosquito samples [100]. In the multiplex PCR assays, a Small Subunit of ribosomal RNA (SSU rRNA) of each Plasmodium spp. can be detected, but it requires a significant amount of parasite DNA, which is not easily achieved with small tissues such as a single pair of salivary glands.
3.4. Treatment and Management of Malaria Cases
Following the expansion of malaria cases in the European Union, the pharmaceutical policy for the treatment of malaria has gradually changed over the years from chloroquine and mefloquine monotherapies used as first-line treatment for uncomplicated malaria to artemisinin combination therapies (ACT) [35,37,105].
For adult treatment, four antimalarial drugs from three different therapeutic classes are routinely recommended [36,106]; these include Artenimol—piperaquine, Artemether—lumefantrine, and Atovaquone—proguanil; the advantages and disadvantages of each of these drugs are summarized in Table 2.
For pregnant women, because of the embryotoxicity and teratogenicity of artemisinin, ACTs are not recommended in the first trimester of pregnancy. The treatment of uncomplicated P. falciparum malaria is based on quinine or, failing that, atovaquone—proguanil during the first trimester. From the second trimester onwards, artemether—lumefantrine is the preferred treatment [38]. For children, the first-line treatments are artemether—lumefantrine; artenimol—piperaquine as shown in Table 3. Mefloquine and atovaquone—proguanil are second-line drugs used in cases of intolerance, contraindication, or failure of ACT treatment. Oral quinine is the third-line drug [35,38].
The choice of these treatments is based on precise clinical and biological criteria, the existence of possible contraindications, the frequency and importance of side effects (particularly those that are potentially severe), the way the drug is taken, and its cost [35,107]. In young children and others at risk of complications (advanced age, co-morbidities, pregnancy, splenectomy), initial hospitalization in an intensive care unit is recommended for the first 48 h, even in the absence of severe criteria [35]. After a minimum incubation period of at least six days, P. falciparum malaria manifests itself mainly as a fever [108]. However, it may be absent at the time of consultation. The associated clinical signs are not specific but headaches are common. Digestive symptoms (nausea, diarrhea, transit disorders) and cough are often present, whatever the age [31]. Therefore, it is essential to look for signs of severity and bacterial co-infection. During management, clinical examination is repeated to detect symptoms suggestive of an early severe form (somnolence, even minimal vigilance disorders, conjunctival subicterus, haemodynamic disorders, dyspnoea) [107]. However, even in the absence of severity criteria, frail patients are treated with intravenous quinine [105].
3.5. Airport Malaria Mass Prevention Strategies
Malaria prevention at airports relies heavily on vector control with an insecticide [39]. International health regulations recommend that airports and 400 m perimeter surrounding areas be disinsected to make them free of Anopheles vectors of malaria [13]. In addition, all aircraft and ships (civil, military, or private) arriving from areas at risk of mosquito-borne diseases, particularly from Africa, Asia, the Middle East, and the Indian Ocean islands, must be disinsected before landing [39]. The current recommended dose of insecticide for aircraft spraying is 2% permethrin (25/75), i.e., 0.7 g active ingredient per 100 m3 in various parts of the aircraft, including the cockpit, passenger cabin, cargo compartments, toilets, and overhead/side garment compartments [39,40]. Pre-flight spraying is carried out by crew members just before departure [40]; this measure is under the responsibility of the airlines.
3.6. Major Challenges
Currently, all actions undertaken in the fight against malaria at airports encounter biological and/or operational challenges; below are some of the key challenges.
3.6.1. Which Case Definition Should Be Considered?
Annual reporting of suspected malaria cases by the European Centre for Disease Prevention and Control (ECDC) really started in 2007 [25], following an upsurge of malaria and infectious diseases in the European Union (EU). In 2019, 29 EU and European Economic Area (EEA) countries reported malaria data [25]; among these countries, 27 reported case-based data and 2 reported aggregate data (Belgium and Bulgaria). Twenty-six countries used the EU case definition, two (France and Germany) used another case definition, and one (Belgium) did not specify the case definition used [25,26]. Malaria surveillance is comprehensive but essentially passive. Reporting is mandatory in 26 countries, voluntary in France and Belgium, and classified among the “other” diseases in the UK [25].
In Canada, malaria has been a nationally notifiable disease since 1983 [109]. However, only laboratory-confirmed cases with evidence of Plasmodium spp. in a blood smear are reported to the federal authorities. Reported malaria cases are classified into five categories [109], as follows:
- -
- Induced cases, i.e., confirmed cases of malaria contracted through a blood transfusion from a donor with confirmed parasitemia;
- -
- Indigenous cases, i.e., confirmed case of malaria contracted through a mosquito bite in Canada;
- -
- Imported cases, i.e., confirmed case of malaria contracted outside Canada;
- -
- Confirmed congenital cases i.e., confirmed case of malaria in an infant up to 3 months of age who has not left Canada since birth and for whom the parasite has been confirmed in the mother;
- -
- Probable congenital cases, i.e., confirmed case of malaria in an infant up to 3 months of age who has not left Canada since birth, but for whom the parasite has not been confirmed in the mother. Cases of probable malaria are reported for public health management purposes only.
This classification differs from the WHO classification. In areas where laboratory diagnostic services are available, WHO classifies malaria cases as follows: asymptomatic malaria, confirmed uncomplicated malaria, confirmed severe malaria, and confirmed death from malaria [110].
3.6.2. Delay in Diagnosis and Treatment
In areas of moderate to intense transmission of Plasmodium spp. parasites, people may develop a form of immunity (premonition) after several years of regular exposure to infected mosquito bites. This is a state of protection against the disease, but not against the parasite. Parasitemia usually remains very low and the patient does not develop severe forms of the disease.
In non-endemic regions, the entire population is considered systematically as non-immune. The particular severity of malaria in these areas is due to delays in diagnosis and treatment, as well as a lack of malaria immunity in infected individuals [109]. Of the cases reported to the Canadian Malaria Network, only 20% visited a health care facility within 24 h after symptoms onset and 44% waited more than three days [109,111]. Diagnosis by health professionals was delayed for more than 24 h in 34% of cases [111]. Thirty percent (30%) of patients developed the severe forms of the disease [109].
3.6.3. Non-Compliance with Aircraft Disinsection Regulations
Until 1994, all flights from endemic areas were subject to health screening, according to WHO recommendations [112]. But due to the large number of flights today, controls have quickly become random [112]. After the surge of airport malaria cases in France in 1994, the local health authorities called attention of the airlines to the danger of importing mosquito vectors via flights from malaria endemic regions (sub-Saharan Africa, Southeast Asia, and South America) [13]. Subsequently, the frequencies of infringements in controlling at risk flights significantly decreased [13], e.g., from 26% in 1995 to 13% in 1996. The most common offenses were non-disinfection of the cargo hold (48%) and cabin (13%), insufficient use of aerosols (18%) and incomplete forms (12%) [13]. At the global level, WHO provided new international health regulations in 2005, which had a fairly high compliance rate (77%) [113]. Of the 23% non-compliance, 12% were due to poorly applied procedures or documentary traceability problems, and 11% concerned a lack of disinsection [113].
More critically, disinsection of incoming or outgoing aircrafts is not a requirement for some countries. Therefore, aircrafts registered in countries not covered by disinsection regulations, but flying to countries where this procedure is in force, are required to comply with the regulations.
3.6.4. Drug and Insecticide Resistances
According to estimates, the global malaria incidence has declined by 27% between 2010 and 2015, but this downward trend slowed after 2015, falling by less than 2% between 2015 and 2019 [25]. Progress in the fight against malaria is still stagnating, especially in African countries where the malaria burden is very high [1]. Among the biological challenges reported are parasite resistance to drugs and vector resistance to insecticides.
At present, despite the wide range of antimalarial drugs available for the treatment of malaria, the treatment regimens put in place are all rapidly coming up against the development of drug resistance in Plasmodium spp. parasites [114]. Parasite resistance to certain antimalarial drugs, particularly those originating from the regions of Southeast Asia (Greater Mekong) or South America, where there is resistance to quinine, pyrimethamine-sulfadoxine, and artemisinin, is now spreading throughout the world [115,116]. Despite the emergence of resistance in endemic countries, ACTs remain the mainstay treatment for P. falciparum malaria in all EU/EEA countries [37,117]. However, there is very little data currently available on antimalarial drug resistance in these areas. Meanwhile, ACTs can also fail there due to both decreased sensitivity to artemisinin compounds and resistance to the associated drugs [115]. In France, ACTs are increasingly used routinely, delivered by hospitals and private sector pharmacies. This further increases the risk of inappropriate, incomplete, or incorrectly dosed treatment. Under these conditions, resistance to anti-malarial drugs could take hold more quickly.
In vector control, the prioritization of long-lasting insecticide-treated net (LLIN) distribution campaigns and indoor residual spraying (IRS) over the past decade has resulted in a 37% decrease in the number of new cases worldwide and a 65% decrease in the mortality rate among children under 5 years old [118,119]. However, the malaria burden in sub-Saharan Africa is still a great concern due to high disease transmission and the rapid increase of vector resistance to insecticides. In many areas, mosquitoes are resistant to permethrin, the pyrethroid insecticide used to disinfect aircrafts [120]. This resistance, which was first reported in Côte d’Ivoire in the major malaria vector in sub-Saharan Africa An. gambiae, is now widespread in Central, West, and East Africa [120,121,122]. In addition, although pyrethroids are less toxic to mammals than other insecticides, people who are temporarily or regularly exposed to them could be exposed to health risks [123]. Therefore, there is a need for alternative vector control measures in and around airports, e.g., non-chemical interventions to mitigate vector resistance to insecticides.
4. Discussion
From this review, it appears that the risk of malaria transmission by mosquito vectors breeding in non-endemic countries is not zero. Moreover, this risk is more likely associated with infected tropical vectors coming in by air or sea transportation. Indeed, there is ample evidence that airport malaria is re-emerging in non-endemic areas. The number of cases recorded during the last decade was 7.4 times higher than that observed during the 2000–2010 decade. This increase may be due to climate change, intensification of international trade, the decline of aircraft disinsection, in addition to delays in diagnosis and treatment.
Global warming has become an undeniable and tragic reality, more or less perceived by the world’s population as a whole, and whose scale and impacts on malaria transmission are increasing. Since the publication of the first report of the IPCC (Intergovernmental Panel on Climate Change) in 1990, there is a warning about the rise in temperature and its consequences [124]. Previous predictions on the increase of temperature are now becoming a reality. Whether humans will be able to stop this inexorable increase, for which there is no other option than drastically reducing our greenhouse gas emissions, is questionable. According to the French public meteorological institute, the 2010–2020 decade was the hottest period from least since 1980 [125,126]. This has resulted in an increase in the number of days with average temperatures exceeding 25 °C in summer [125]. On the other hand, the cases of airport malaria observed in the last decade occurred mainly in summer between June and September, when climatic conditions were favorable to the survival of the mosquitos (average temperature 23 °C, range: 17–31 °C) [127]. One of the key factors controlling mosquito development both in water and on land, as well as parasite development in mosquitoes, is the temperature. Indeed, the duration of sporogony, mosquito longevity, and the duration of larval development are closely influenced by the temperature [128].
On a global scale, human populations passed the 7 billion mark in 2011 [129]. This has resulted in an increase in the number of international trips and airlines have increased the number of flights across the world’s capitals. It should also be noted that most of the patients suffering with airport malaria live near or worked at an international airport. Field studies revealed that these patients are infected by vectors coming inland by air transportation, i.e., which escaped the chemical treatment of aircrafts. Nowadays, aircraft disinsection is declining in most countries. Although WHO continues to recommend disinsection of aircrafts, there are concerns about the health effects of pyrethroid insecticides used and the resulting lawsuits have led many airlines and governments to stop the spraying practices [6]. To strengthen the capacity of airlines, the ICAO (International Civil Aviation Organization) developed a vector control register in 2016, in accordance with the WHO Manual on Vector Surveillance and Control at Ports, Airports and Border Crossings [130]. Therefore, decisions are made on a global scale, but very few are implemented. More recently in 2019, a number of logistical, practical, and financial challenges in implementing additional disinsection requirements has been identified and recommendations made for further research, development, and implementation actions [130]. Accordingly, current efforts should lead to non-chemical insect control in order to overcome insecticide resistance and better prevent malaria in aircrafts, as well as in and around airports [130], along with early diagnosis and appropriate treatment of cases.
Currently, hopes have turned to biological diagnosis. Unfortunately, biologists in non-endemic areas are facing several shortcomings in performing and reading the thick drop, in detecting parasites on the smear of pauci-parasitic patients, and in determining the species when the parasite density is low [28,33]. These difficulties are sometimes increased by the previous use of drugs that alter the morphology of the parasites. In addition, the use of rapid diagnostic tests (RDT) poses problems in terms of sensitivity due to the deletion of the pfhrp 2/3 genes in P. falciparum [131,132,133]. Currently, more than 28 countries in Africa and southwest Asia have reported deletions of the pfhrp2/3 genes [1,134], which cause false negative RDT results in infection detection. In this situation, the probability of the patient to receive adequate treatment is low. This probability is even lower when it comes to airport malaria in non-endemic areas since most physicians have little reason to suspect it. In addition, the treatment initiated by the physician may be inappropriate due to the resistance of Plasmodium spp. to certain antimalarial drugs [28,88,135]. In the Southeast Asia region, failure rates for artemisinin-based combinations is above 10% [1]. In the Eastern Mediterranean region, the failure of sulfadoxine-pyrimethamine treatments against P. falciparum has led countries such as Somalia and Sudan to recommend artemether-lumefantrine as a first-line treatment [1]. Chloroquine is effective against P. vivax in most countries except Myanmar and East Timor [1]. For a better monitoring of the resistance of Plasmodium spp. to antimalarial drugs, it would be interesting to set up a resistance-mapping tool. The mapping tool will guide physicians in countries where malaria is not endemic to get an idea of the resistance of Plasmodium spp. to antimalarial drugs on a global scale. Nevertheless, the deployment of any intervention must be accompanied by rigorous routine entomological and epidemiological surveillance activities to monitor the success of the intervention and continuously inform the policies.
5. Conclusions
This review provides an update on the status of airport malaria in non-endemic areas. Although significant progress has been made in previous decades to reduce the disease burden worldwide, airport malaria is re-emerging in most countries of the European Union. This increase may be associated with climate change, growth of international travels, failure in aircraft disinsection, on top of delays in case diagnosis and treatment. Key issues hindering malaria case management and vector control interventions in and around the airports, e.g., delay in diagnosis and treatment, drug and insecticide resistance, technical and logistic shortcomings, should further be addressed by policy makers and disease control programmes. Moreover, the deployment of any intervention must be accompanied by rigorous routine entomological and epidemiological surveillance activities to inform policy in real time and monitor the effectiveness of the interventions.
Author Contributions
L.D.A. and J.E. designed and developed the study protocol. L.D.A. interpreted and analyzed the data, and drafted the manuscript. J.E. critically reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The APC were supported by the European Union Horizon 2020 Framework Programme (688207-DMC-MALVEC Project).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets supporting the findings of this article are included within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- WHO. World Malaria Report 2020; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Velasco, E.; Gomez-Barroso, D.; Akikwa, D.; Diaz, O.; Cano, R. Non-imported malaria in non-endemic countries: A review of cases in Spain. Malar. J. 2017, 16. [Google Scholar] [CrossRef]
- Newman, R.D.; Parise, M.E.; Barber, A.M.; Steketee, R.W. Malaria-related deaths among U.S. travelers, 1963–2001. Ann. Intern. Med. 2004, 141, 547–555. [Google Scholar] [CrossRef] [Green Version]
- Diallo, M.L.P. Étudiants Subsahariens à Toulouse et Prévention du Paludisme d’Importation. Practice Thesis, Université Toulouse III, Paul Sabatier, France, 2018; p. 78. Available online: http://thesesante.ups-tlse.fr/2140/ (accessed on 14 August 2021).
- Muentener, P.; Schlagenhauf, P.; Steffen, R. Le paludisme Importé (1985–1995): Tendances et Perspectives. Bull. De L’organisation Mond. De La Santé La Rev. Int. De Santé Publique Recl. D’articles 2000, 2, 57–63. Available online: https://apps.who.int/iris/handle/10665/57460 (accessed on 18 August 2021).
- Tatem, A.J.; Rogers, D.J.; Hay, S.I. Estimating the malaria risk of African mosquito movement by air travel. Malar. J. 2006, 5, 57. [Google Scholar] [CrossRef] [Green Version]
- Lusina, D.; Legros, F.; Esteve, V.; Klerlein, M.; Giacomini, T. Airport malaria: Four new cases in Suburban Paris during summer 1999. Euro Surveill. 2000, 5, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Isaäcson, M. Airport malaria: A Review. Bull. World Health Organ. 1989, 67, 737–743. [Google Scholar] [PubMed]
- Gratz, N.G.; Steffen, R.; Cocksedge, W. Why aircraft disinsection? Bull. World Health Organ. 2000, 78, 995–1004. [Google Scholar] [CrossRef]
- World Health Organization. Airport malaria: A fatal case. Wkly. Epidemiol. Rec. 1996, 71, 358. [Google Scholar]
- Chapuis, Y.; Chaussain, J.-C. Influences du transport aérien sur la santé. Acad. Natl. Méd. 2009. Available online: https://www.academie-medecine.fr/influences-du-transport-aerien-sur-la-sante/ (accessed on 8 March 2021).
- Mier-y-Teran-Romero, L.; Tatem, A.J.; Johansson, M.A. Mosquitoes on a plane: Disinsection will not stop the spread of vector-borne pathogens, a simulation study. PLoS Negl. Trop. Dis. 2017, 11, e0005683. [Google Scholar] [CrossRef]
- Guillet, P.; Germain, M.C.; Giacomini, T.; Chandre, F.; Akogbeto, M.; Faye, O.; Kone, A.; Manga, L.; Mouchet, J. Origin and prevention of airport malaria in France. Trop. Med. Int. Health 1998, 3, 700–705. [Google Scholar] [CrossRef]
- United Nations. Standard Country or Area Codes for Statistical Use (M49). Available online: https://unstats.un.org/unsd/methodology/m49/overview/ (accessed on 25 September 2021).
- Fang, Y.; Shi, W.-Q.; Zhang, Y. Molecular phylogeny of Anopheles hyrcanus group members based on ITS2 RDNA. Parasites Vectors 2017, 10, 417. [Google Scholar] [CrossRef] [Green Version]
- Miaoulis, M.; Giantsis, I.A.; Schaffner, F.; Chaskopoulou, A. Re-examination of the taxonomic status of Anopheles hyrcanus and An. pseudopictus using a multilocus genetic approach. J. Vector Ecol. 2018, 43, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Harbach, R. The classification of genus Anopheles (Diptera: Culicidae): A working hypothesis of phylogenetic relationships. Bull. Entomol. Res. 2004, 94, 537–553. [Google Scholar] [CrossRef]
- Artemov, G.N.; Velichevskaya, A.I.; Bondarenko, S.M.; Karagyan, G.H.; Aghayan, S.A.; Arakelyan, M.S.; Stegniy, V.N.; Sharakhov, I.V.; Sharakhova, M.V. A standard photomap of the ovarian nurse cell chromosomes for the dominant malaria vector in Europe and Middle East Anopheles sacharovi. Malar. J. 2018, 17, 276. [Google Scholar] [CrossRef] [Green Version]
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Kabaria, C.W.; et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: Occurrence data, distribution maps and bionomic précis. Parasites Vectors 2010, 3, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanafi-Bojd, A.A.; Azari-Hamidian, S.; Hassan, V.; Zabihollah, C. Spatio—temporal distribution of malaria vectors (Diptera: Culicidae) across different climatic zones of Iran. Asian Pac. J. Trop. Med. 2011, 4, 498–504. [Google Scholar] [CrossRef] [Green Version]
- Vatandoost, H.; Abai, M.R. Irritability of malaria vector, Anopheles sacharovi to different insecticides in a malaria–prone Area. Asian Pac. J. Trop. Med. 2012, 5, 113–116. [Google Scholar] [CrossRef] [Green Version]
- Birnberg, L.; Aranda, C.; Talavera, S.; Núñez, A.I.; Escosa, R.; Busquets, N. Laboratory colonization and maintenance of Anopheles atroparvus from the Ebro Delta, Spain. Parasites Vectors 2020, 13, 394. [Google Scholar] [CrossRef] [PubMed]
- Hackett, L.W. The present status of our knowledge of the sub-species of Anopheles maculipennis. Trans. R. Soc. Trop. Med. Hyg. 1934, 28, 109–140. [Google Scholar] [CrossRef] [Green Version]
- Danis, M.; Mouchet, M.; Giacomini, T.; Guillet, P.; Legros, F.; Belkaïd, M. Paludisme autochtone et introduit en Europe. Méd. Mal. Infect. 1996, 26, 393–396. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). Malaria Annual Epidemiological Report. 2020. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/AER-malaria-2019.pdf (accessed on 14 August 2021).
- European Centre for Disease Prevention and Control (ECDC). The European Surveillance System (TESSy). 2015. Available online: https://www.ecdc.europa.eu/en/publications-data/european-surveillance-system-tessy (accessed on 31 May 2021).
- Gentilini, M.; Danis, M. Le paludisme autochtone. Méd. Mal. Infect. 1981, 11, 356–362. [Google Scholar] [CrossRef]
- Queyriaux, B.; Pradines, B.; Hasseine, L.; Coste, S.; Rodriguez, P.; Coffinet, T.; Haus-Cheymol, R.; Rogier, C. Paludisme d’aéroport. Presse Med. 2009, 38, 1106–1109. [Google Scholar] [CrossRef] [PubMed]
- Thellier, M.; Lusina, D.; Guiguen, C.; Delamaire, M.; Legros, F.; Cicéron, L.; Klerlein, M.; Danis, M.; Mazier, D. Is airport malaria a transfusion-transmitted malaria risk? Transfusion 2001, 41, 301–302. [Google Scholar] [CrossRef]
- Mace, K.E.; Lucchi, N.W.; Tan, K.R. Malaria Surveillance—United States, 2017. Morb. Mortal. Wkly. Rep. 2021, 70, 1–35. [Google Scholar] [CrossRef]
- Wieters, I.; Eisermann, P.; Borgans, F.; Giesbrecht, K.; Goetsch, U.; Just-Nübling, G.; Kessel, J.; Lieberknecht, S.; Muntau, B.; Tappe, D.; et al. Two cases of airport-associated falciparum malaria in Frankfurt am main, Germany, October 2019. Eurosurveillance 2019, 24, 1900691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capek, I. Epidémiologie Du Paludisme d’importation à Plasmodium falciparum et de la chimiorésistance. Méd. Mal. Infect. 1999, 29, S178–S186. [Google Scholar] [CrossRef]
- De Pina, J.J.; Garnotel, E.; Hance, P.; Vedy, S.; Rogier, C.; Morillon, M. Diagnostic du paludisme d’importation en France. Méd. Mal. Infect. 2009, 37, 710–715. [Google Scholar] [CrossRef]
- Bigaillon, C.; Fontan, E.; Cavallo, J.-D.; Hernandez, E.; Spiegel, A. Ineffectiveness of the Binax NOW Malaria Test for Diagnosis of Plasmodium ovale Malaria. J. Clin. Microbiol 2005, 43, 1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strady, C. Prise en Charge et Prévention du Paludisme D’importation. Groupe Recommandations de la Société de Pathologie In-Fectieuse de Langue Française (SPILF). 2017, p. 71. Available online: https://www.antibioest.org/wp-content/uploads/2018/08/paludisme-Pr-Strady-JRI-2017.pdf (accessed on 14 October 2021).
- Hatz, C.; Soto, J.; Nothdurft, H.D.; Zoller, T.; Weitzel, T.; Loutan, L.; Bricaire, F.; Gay, F.; Burchard, G.-D.; Andriano, K.; et al. Treatment of acute uncomplicated falciparum malaria with artemether-lumefantrine in nonimmune populations: A safety, efficacy, and pharmacokinetic study. Am. J. Trop. Med. Hyg. 2008, 78, 241–247. [Google Scholar] [CrossRef]
- OMS. Traitement du Paludisme—Tour d’Horizon; World Health Organization: Geneva, Switzerland, 2018; Available online: http://www.who.int/malaria/areas/treatment/overview/fr/ (accessed on 25 May 2021).
- European Medicines Agency (EMA). Public Summary of Opinion on Orphan Designation Dihydroartemisin, Piperaquine for the Treatment of Malaria; European Medicines Agency: Amsterdam, The Netherlands, 2011; Volume 4.
- OMS. Surveillance des vecteurs et lutte antivectorielle dans les ports, les aéroports et aux postes-frontières. In Règlement Sanitaire International; World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- Casida, J.E.; Quistad, G.B. Golden age of insecticide research: Past, present, or future? Annu. Rev. Entomol. 1998, 43, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Feachem, R.G.; Phillips, A.A.; Hwang, J.; Cotter, C.; Wielgosz, B.; Greenwood, B.M.; Sabot, O.; Rodriguez, M.H.; Abeyasinghe, R.R.; Ghebreyesus, T.A.; et al. Shrinking the Malaria Map: Progress and Prospects. Lancet 2010, 376, 1566–1578. [Google Scholar] [CrossRef] [Green Version]
- OMS. Rapport Sur le Paludisme Dans le Monde; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Hantosh, H.A.; Hassan, H.M.; Ahma, B.; Al-fatlawy, A. Mosquito species geographical distribution in Iraq 2009. Vector Borne Dis. 2012, 49, 33. Available online: https://www.mrcindia.org/journal/issues/491033.pdf (accessed on 14 October 2021).
- Özbilgin, A.; Topluoglu, S.; Es, S.; Islek, E.; Mollahaliloglu, S.; Erkoc, Y. Malaria in Turkey: Successful control and strategies for achieving elimination. Acta Trop. 2011, 120, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Vatandoost, H.; Ashraf, H.; Lak, S.; Mahdi, R.; Abai, M.; Nazari, M. Factors involved in the re-emergence of malaria in borderline of Iran, Armenia, Azerbaijan and Turkey. Southeast Asian J. Trop. Med. Public Health 2003, 34 (Suppl. S2), 6–14. [Google Scholar] [PubMed]
- Piperaki, E.T.; Daikos, G.L. Malaria in Europe: Emerging threat or minor nuisance? Clin. Microbiol. Infect. 2016, 22, 487–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odolini, S.; Gautret, P.; Parola, P. Epidemiology of imported malaria in the mediterranean region. Mediterr. J. Hematol. Infect. Dis. 2012, 4, e2012031. [Google Scholar] [CrossRef] [PubMed]
- Ponçon, N.; Tran, A.; Toty, C.; Luty, A.J.; Fontenille, D.A. Quantitative risk assessment approach for mosquito-borne diseases: Malaria re-emergence in Southern France. Malar. J. 2008, 7, 147. [Google Scholar] [CrossRef] [Green Version]
- Brugman, V.A.; Hernández-Triana, L.M.; England, M.E.; Medlock, J.M.; Mertens, P.P.C.; Logan, J.G.; Wilson, A.J.; Fooks, A.R.; Johnson, N.; Carpenter, S. Blood-feeding patterns of native mosquitoes and insights into their potential role as pathogen vectors in the Thames estuary region of the United Kingdom. Parasites Vectors 2017, 10, 163. [Google Scholar] [CrossRef] [Green Version]
- Lourenço, P.M.; Sousa, C.A.; Seixas, J.; Lopes, P.; Novo, M.T.; Almeida, A.P.G. Anopheles atroparvus density modeling using MODIS NDVI in a former malarious area in Portugal. J. Vector Ecol. 2011, 36, 279–291. [Google Scholar] [CrossRef]
- Kampen, H.; Schäfer, M.; Zielke, D.E.; Walther, D. The Anopheles maculipennis complex (Diptera: Culicidae) in Germany: An update following recent monitoring activities. Parasitol. Res. 2016, 115, 3281–3294. [Google Scholar] [CrossRef]
- Bueno-Marí, R.; Jiménez-Peydró, R. Study of the malariogenic potential of Eastern Spain. Trop. Biomed. 2012, 29, 39–50. [Google Scholar]
- de Zulueta, J.; Ramsdale, C.D.; Coluzzi, M. Receptivity to malaria in Europe. Bull. World Health Organ. 1975, 52, 109–111. [Google Scholar]
- Cambournac, F.J. Contribution to the history of malaria epidemiology and control in Portugal and some other places. Parassitologia 1994, 36, 215–222. [Google Scholar]
- Sousa, C.A. Malaria Vectorial Capacity and Competence of Anopheles Atroparvus Van Thiel, 1927 (Diptera, Culicidade): Implications for the Potential Re-Emergence of Malaria in Portugal; Institute of Hygiene and Tropical Medicine: Lisboa, Portugal, 2008. [Google Scholar]
- Boccolini, D.; Toma, L.; Luca, M.D.; Severini, F.; Cocchi, M.; Bella, A.; Massa, A.; Barbieri, F.M.; Bongiorno, G.; Angeli, L.; et al. Impact of environmental changes and human-related factors on the potential malaria vector, Anopheles labranchiae (Diptera: Culicidae), in Maremma, Central Italy. J. Med. Entomol. 2012, 49, 833–842. [Google Scholar] [CrossRef]
- d’Alessandro, G.; Smiraglia, C.B.; Lavagnino, A.; World Health Organization. Further Studies on the Biology of Anopheles labranchiae Falleroni in Sicily, WHO/MAL/71.754; World Health Organization: Geneva, Switzerland, 1971.
- Romi, R.; Pierdominici, G.; Severini, C.; Tamburro, A.; Cocchi, M.; Menichetti, D.; Pili, E.; Marchi, A. Status of malaria vectors in Italy. J. Med. Entomol. 1997, 34, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Di Luca, M.; Boccolini, D.; Severini, F.; Toma, L.; Barbieri, F.M.; Massa, A.; Romi, R. A 2-year entomological study of Potential malaria vectors in central Italy. Vector-Borne Zoonotic Dis. 2009, 9, 703–711. [Google Scholar] [CrossRef]
- Aitken, T.H.G. The Culicidae of Sardinia and Corsica (Diptera). Bull. Entomol. Res. 1954, 45, 437–494. [Google Scholar] [CrossRef]
- Lavagnino, A. Anopheles labranchiae in Sicily: Decrease of susceptibility to insecticides. Trans. R. Soc. Trop. Med. Hyg. 1983, 77, 883. [Google Scholar] [CrossRef]
- Toty, C.; Barré, H.; Le Goff, G.; Larget-Thiéry, I.; Rahola, N.; Couret, D.; Fontenille, D. Malaria risk in Corsica, former hot spot of malaria in France. Malar. J. 2010, 9, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, G.M. Geographical Distribution of Arthropod-Borne Diseases and Their Principal Vectors; World Health Organization, Division of Vector Biology and Control: Geneva, Switzerland, 1989; pp. 7–22. [Google Scholar]
- Adamovic, Z. Anopheles messeae Falleroni in Serbia and Macedonia (Diptera, Culicidae). Acta Vet. 1980, 30, 217–223. [Google Scholar]
- Adamovic, Z. Vertical distribution of the anopheline mosquitoes in Montenegro, Yugoslavia. Acta Vet. 1984, 34, 287–293. [Google Scholar]
- Takken, W.; Geene, R.; Adam, W.; Jetten, T.H.; van der Velden, J.A. Distribution and dynamics of larval populations of Anopheles messeae and An. atroparvus in the delta of the rivers rhine and Meuse, The Netherlands. Ambio 2002, 31, 212–218. [Google Scholar] [CrossRef]
- Fyodorova, M.V.; Savage, H.M.; Lopatina, J.V.; Bulgakova, T.A.; Ivanitsky, A.V.; Platonova, O.V.; Platonov, A.E. Evaluation of potential West Nile virus vectors in Volgograd region, Russia, 2003 (Diptera: Culicidae): Species composition, bloodmeal host utilization, and virus infection rates of mosquitoes. J. Med. Entomol. 2006, 43, 552–563. [Google Scholar] [CrossRef]
- Bates, M. Field Studies of the anopheline mosquitoes of Albania. In Proceedings of the Entomological Society of Washington, Washington, DC, USA, 26 March 1941; Volume 43. Available online: https://www.biodiversitylibrary.org/page/16130453#page/81/mode/1up (accessed on 14 October 2021).
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010; Available online: https://books.google.cm/books?id=JIfgVr1f4IQC (accessed on 20 August 2021).
- Daskova, N.G.; Rasnicyn, S.P. Review of data on susceptibility of mosquitoes in the USSR to imported strains of malaria parasites. Bull. World Health Organ. 1982, 60, 893–897. [Google Scholar]
- Linton, Y.; Lee, A.S.; Curtis, C.F. Discovery of a third member of the Maculipennis group in South West England. Eur. Mosq. Bull. 2005, 19, 5–9. [Google Scholar]
- Rueda, L.M.; Kim, H.-C.; Klein, T.A.; Pecor, J.E.; Li, C.; Sithiprasasna, R.; Debboun, M.; Wilkerson, R.C. Distribution and larval habitat characteristics of Anopheles hyrcanus group and related mosquito species (Diptera: Culicidae) in South Korea. J. Vector Ecol. 2006, 31, 198–205. [Google Scholar] [CrossRef]
- Harbach, R.E. Review of the internal classification of the genus Anopheles (Diptera: Culicidae): The foundation for comparative systematics and phylogenetic research. Bull. Entomol. Res. 1994, 84, 331–342. [Google Scholar] [CrossRef]
- Zu-Jie, Z. The malaria situation in the people’s Republic of China. Bull. World Health Organ. 1981, 59, 931–936. [Google Scholar]
- Lee, W.-J.; Klein, T.A.; Kim, H.-C.; Choi, Y.-M.; Yoon, S.-H.; Chang, K.-S.; Chong, S.-T.; Lee, I.-Y.; Jones, J.W.; Jacobs, J.S.; et al. Anopheles kleini, Anopheles pullus, and Anopheles sinensis: Potential Vectors of Plasmodium vivax in the Republic of Korea. J. Med. Entomol. 2007, 44, 1086–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gingrich, J.B.; Weatherhead, A.; Sattabongkot, J.; Pilakasiri, C.; Wirtz, R.A. Hyperendemic malaria in a Thai Village: Dependence of year-round transmission on focal and seasonally circumscribed mosquito (Diptera: Culicidae) habitats. J. Med. Entomol. 1990, 27, 1016–1026. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.S.; Khan, M.G.M.; Chaudhury, N.; Deloer, S.; Nazib, F.; Bangali, A.M.; Haque, R. Prevalence of Anopheline species and Their Plasmodium Infection Status in Epidemic-Prone Border Areas of Bangladesh. Malar. J. 2010, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Robert, V. Les Changements Globaux Aggravent le Risque D’expansion du Paludisme. Available online: http://theconversation.com/les-changements-globaux-aggravent-le-risque-dexpansion-du-paludisme-115951 (accessed on 23 August 2021).
- Rageau, J. The Geographical Distribution of Mosquitoes in New Caledonia and Dependencies with Keys for Their Identification; Commission du Pacifique Sud: Noumea, New Caledonia, 1958; pp. 1–19. (In French) [Google Scholar]
- Institut Pasteur New Caledonia. Annual Report 2016; Institut Pasteur de Nouvelle-Calédonie: Noumea, New Caledonia, 2017; pp. 23–33. [Google Scholar]
- Pol, M.; Kilama, S.; Duperier, S.; Soupé-Gilbert, M.-E.; Calvez, E.; Pocquet, N. Introduction of the Anopheles bancroftii mosquito, a malaria vector, into New Caledonia. Emerg. Infect. Dis. 2018, 24, 604–605. [Google Scholar] [CrossRef]
- Siala, E.; Gamara, D.; Kallel, K.; Daaboub, J.; Zouiten, F.; Houzé, S.; Bouratbine, A.; Aoun, K. Airport malaria: Report of four cases in Tunisia. Malar. J. 2015, 14, 42. [Google Scholar] [CrossRef] [Green Version]
- Jeyaprakasam, N.K.; Liew, J.W.K.; Low, V.L.; Wan-Sulaiman, W.-Y.; Vythilingam, I. Plasmodium knowlesi infecting Humans in Southeast Asia: What’s next? PLoS Negl. Trop. Dis. 2020, 14, e0008900. [Google Scholar] [CrossRef]
- Wesolowski, R.; Wozniak, A.; Mila-Kierzenkowska, C.; Szewczyk-Golec, K. Plasmodium knowlesi as a threat to global public health. Korean J. Parasitol. 2015, 53, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Ooi, C.H.; Bujang, M.A.; Tg Abu Bakar Sidik, T.M.I.; Ngui, R.; Lim, Y.A.-L. Over Two Decades of Plasmodium Knowlesi Infections in Sarawak: Trend and Forecast. Acta Trop. 2017, 176, 83–90. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). Annual Epidemiological Report on Communicable Diseases in Europe; European Centre for Disease Prevention and Control: Solna Municipality, Sweden, 2010.
- Lounibos, L.P. Invasions by insect vectors of human disease. Annu. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef]
- Gardner, L.; Sarkar, S.A. Global airport-based risk model for the spread of dengue infection via the air transport network. PLoS ONE 2013, 8, e72129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Société de Pathologie Infectieuse de Langue Française. 12ème Conférence de Consensus en Thérapeutique Anti-Infectieuse de la Société de Pathologie Infectieuse de Langue Française (SPILF). Prise en Charge et Prévention du Paludisme D’importation à Plasmodium falciparum. Available online: https://www.srlf.org/wp-content/uploads/2015/11/0807-Reanimation-Vol17-N5-pe1_e54.pdf (accessed on 14 October 2021).
- Fromage, M.; Houze, S.; Miegeville, M.; Thellier, M. Annales du Contrôle National de Qualité des Analyses de Biologie Médicale; Agence Française de Sécurité Sanitaire des Produits de Santé: Paris, France, 2010.
- OMS. Diagnostic du Paludisme—Tour D’horizon; World Health Organization: Geneva, Switzerland, 2018; Available online: http://www.who.int/malaria/areas/diagnosis/overview/fr/ (accessed on 15 May 2021).
- Grobusch, M.P.; Hänscheid, T.; Zoller, T.; Jelinek, T.; Burchard, G.D. Rapid immunochromatographic malarial antigen detection unreliable for detecting Plasmodium malariae and Plasmodium ovale. Eur. J. Clin. Microbiol. Infect. Dis. 2002, 21, 818–820. [Google Scholar] [CrossRef] [PubMed]
- Bylicka-Szczepanowska, E.; Lass, A.; Korzeniewski, K. Prevalence of Plasmodium spp. in symptomatic BaAka Pygmies inhabiting the rural Dzanga Sangha Region, Central African Republic. Ann. Agric. Environ. Med. 2021, 28, 483–490. [Google Scholar] [CrossRef]
- MacDonald, G. The Epidemiology and Control of Malaria; Oxford University Press: Oxford, UK, 1957; p. 201. [Google Scholar]
- Burkot, T.R.; Williams, J.L.; Schneider, I. Identification of Plasmodium falciparum-infected mosquitoes by a double antibody enzyme-linked immunosorbent assay. Am. J. Trop. Med. Hyg. 1984, 33, 783–788. [Google Scholar] [CrossRef] [PubMed]
- Fontenille, D.; Meunier, J.Y.; Nkondjio, C.A.; Tchuinkam, T. Use of circumsporozoite protein enzyme-linked immunosorbent assay compared with microscopic examination of salivary glands for calculation of malaria infectivity rates in mosquitoes (diptera: culicidae) from cameroon. J. Med. Entomol. 2001, 38, 451–454. [Google Scholar] [CrossRef]
- Vaughan, J.A.; Noden, B.H.; Beier, J.C. Population dynamics of Plasmodium falciparum sporogony in laboratory-infected Anopheles gambiae. J. Parasitol. 1992, 78, 716–724. [Google Scholar] [CrossRef]
- Zaman, S.; Tan, L.; Chan, H.H.; Aziz, L.; Abdul-Samat, S.; Wahid, R.; Kamal, A.; Ahmed, M.; Zaman, V. The detection of Plasmodium falciparum and P. Vivax in DNA-extracted blood samples using polymerase chain reaction. Trans. R. Soc. Trop. Med. Hyg. 2001, 95, 391–397. [Google Scholar] [CrossRef]
- Padley, D.; Moody, A.H.; Chiodini, P.L.; Saldanha, J. Use of a rapid, single-round, multiplex PCR to detect malarial parasites and identify the species present. Ann. Trop. Med. Parasitol. 2003, 97, 131–137. [Google Scholar] [CrossRef]
- Fabre, R.; Berry, A.; Morassin, B.; Magnaval, J.F. Comparative assessment of conventional PCR with multiplex real-time PCR using SYBR green I detection for the molecular diagnosis of imported malaria. Parasitology 2004, 128, 15–21. [Google Scholar] [CrossRef]
- Swan, H.; Sloan, L.; Muyombwe, A.; Chavalitshewinkoon-Petmitr, P.; Krudsood, S.; Leowattana, W.; Wilairatana, P.; Looareesuwan, S.; Rosenblatt, J. Evaluation of a real-time polymerase chain reaction assay for the diagnosis of malaria in patients from thailand. Am. J. Trop. Med. Hyg. 2005, 73, 850–854. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, N.; Boutet, A.; Bousquet, P.J.; Basset, D.; Douard-Enault, C.; Charachon, S.; Lachaud, L. Comparison of three real-time PCR methods with blood smears and rapid diagnostic test in Plasmodium spp. infection. Clin. Microbiol. Infect. 2010, 16, 1305–1311. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.S.; Ranford-Cartwright, L.C. A real-time PCR assay for quantifying Plasmodium falciparum infections in the mosquito vector. Int. J. Parasitol. 2004, 34, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Boissière, A.; Gimonneau, G.; Tchioffo, M.T.; Abate, L.; Bayibeki, A.; Awono-Ambéné, P.H.; Nsango, S.E.; Morlais, I. Application of a qPCR assay in the investigation of susceptibility to malaria infection of the M and S molecular forms of An. gambiae s.s. in Cameroon. PLoS ONE 2013, 8, e54820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debord, T.; Danis, M. Prise en Charge et Prévention du Paludisme D’importation à Plasmodium Falciparum: Recommendations Pour la Pratique Clinique 2007 (Révision de la Conférence de Consensus 1999) Texte Long; Société de Pathologie Infectieuse de Langue Française: Paris, France, 2008. [Google Scholar]
- van Agtmael, M.; Bouchaud, O.; Malvy, D.; Delmont, J.; Danis, M.; Barette, S.; Gras, C.; Bernard, J.; Touze, J.-E.; Gathmann, I.; et al. The comparative efficacy and tolerability of CGP 56697 (artemether+lumefantrine) versus halofantrine in the treatment of uncomplicated falciparum malaria in travellers returning from the tropics to the Netherlands and France. Int. J. Antimicrob. Agents 1999, 12, 159–169. [Google Scholar] [CrossRef]
- Le Loup, G.; Malvy, D. Paludisme d’importation. Mal. Infect. 2010. Available online: https://www.em-consulte.com/article/237223/paludisme-d-importation (accessed on 19 May 2021).
- Thang, H.D.; Elsas, R.M.; Veenstra, J. Airport malaria: Report of a case and a brief review of the literature. Neth. J. Med. 2002, 60, 441–443. [Google Scholar]
- Canada, A. de la Santé Publique du. Recommandations du CCMTMV Sur le Diagnostic et le Traitement du Paludisme. Available online: https://www.canada.ca/fr/sante-publique/services/rapports-publications/releve-maladies-transmissibles-canada-rmtc/numero-mensuel/2014-40/rmtc-volume-40-7-3-avril-2014/rmtc-volume-40-7-3-avril-2014.html (accessed on 1 June 2021).
- OMS. Normes Recommendées par l’OMS Pour la Surveillance; Organisation Mondiale de la Santé: Genève, Switzerland, 2000; Available online: http://apps.who.int/iris/bitstream/handle/10665/65894/WHO_CDS_CSR_ISR_99.2_pp.1-100_fre.pdf?sequence=1 (accessed on 14 August 2021).
- Boggild, A.; Brophy, J.; Charlebois, P.; Crockett, M.; Geduld, J.; Ghesquiere, W.; McDonald, P.; Plourde, P.; Teitelbaum, P.; Tepper, M.; et al. Summary of Recommendations for the Diagnosis and Treatment of Malaria by the Committee to Advise on Tropical Medicine and Travel (CATMAT). Can. Commun. Dis. Rep. 2014, 40, 133–143. [Google Scholar] [CrossRef]
- Petersen, P.E. Report of the informal consultation on aircraft disinsection. Commun. Dent. Oral Epidemiol. 1995, 31, 471. [Google Scholar] [CrossRef]
- Boulanger, H. Mise en Œuvre du Nouveau Règlement Sanitaire International Dans le Cadre du Contrôle Sanitaire aux Frontières à L’aéroport International de Lyon Saint-Exupéry; Ecole des Hautes études en Santé Publique: Paris, France, 2008; Available online: https://documentation.ehesp.fr/memoires/2008/ies/boulanger.pdf (accessed on 7 June 2021).
- Udom, B.-B. Programmes nationaux de lutte contre le paludisme: Pharmaciens, engagez-vous! ReMeD 2009, 24. Available online: https://remed.org/wp-content/uploads/2017/03/revue39-ReMeD-pharmacien-lutte-contre-paludisme.pdf (accessed on 5 June 2021).
- Nsanzabana, C.; Ariey, F.; Beck, H.-P.; Ding, X.C.; Kamau, E.; Krishna, S.; Legrand, E.; Lucchi, N.; Miotto, O.; Nag, S.; et al. Molecular assays for antimalarial drug resistance surveillance: A target product profile. PLoS ONE 2018, 13, e0204347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menard, D.; Dondorp, A. Antimalarial drug resistance: A threat to malaria elimination. Cold Spring Harb. Perspect. Med. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Orsi, F.; Zimmermann, J.-B. Le marché des antipaludéens, entre régulation et défaillance. Mondes Dev. 2015, 170, 21–40. [Google Scholar] [CrossRef] [Green Version]
- Chebil, M. La lutte contre le paludisme souffre aussi du Covid-19. Available online: https://www.france24.com/fr/afrique/20210425-la-lutte-contre-le-paludisme-souffre-aussi-du-covid-19 (accessed on 9 June 2021).
- Zogo, M.B. Impact Entomologique de Stratégies Complémentaires de Lutte Intégrée Contre Le Paludisme Dans Un Contexte de Résistance Des Vecteurs Aux Insecticides Au Nord de La Côte d’Ivoire. Ph.D. Thesis, Université de Montpellier, Montpellier, France, Université d’Abomey-Calavi, Cotonou, Benin, 2019; 211p. Available online: https://tel.archives-ouvertes.fr/tel-02485637/file/2019_ZOGO_archivage.pdf (accessed on 9 June 2021).
- Groupe Régional WASH. Lutte Antivectorielle Contre le Moustique Anophèles Dans le Cadre de la Lutte Contre le Paludisme. 2017. Available online: https://www.humanitarianresponse.info/sites/www.humanitarianresponse.info/files/documents/files/guidance_technique_lutte_antivectorielle_contre_le_paludisme_-_wash_wcar_group_-_fr_2017.04.07.pdf (accessed on 14 August 2021).
- Elissa, N.; Mouchet, J.; Riviere, F.; Meunier, J.Y.; Yao, K. Resistance of Anopheles gambiae s.s. to pyrethroids in Côte d’Ivoire. Ann. Soc. Belg. Med. Trop. 1993, 73, 291–294. [Google Scholar] [PubMed]
- Menze, B.D.; Wondji, M.J.; Tchapga, W.; Tchoupo, M.; Riveron, J.M.; Wondji, C.S. Bionomics and insecticides resistance profiling of malaria vectors at a selected site for experimental Hut Trials in Central Cameroon. Malar. J. 2018, 17, 317. [Google Scholar] [CrossRef]
- Soderlund, D.M.; Clark, J.M.; Sheets, L.P.; Mullin, L.S.; Piccirillo, V.J.; Sargent, D.; Stevens, J.T.; Weiner, M.L. Mechanisms of pyrethroid neurotoxicity: Implications for cumulative risk assessment. Toxicology 2002, 171, 3–59. [Google Scholar] [CrossRef]
- Groupe d’Experts Intergouvernemental sur l’Evolution du Climat (GIEC). Réchauffement Planétaire. Climat. 2021. Available online: https://climat.be/changements-climatiques/changements-observes/rechauffement-planetaire (accessed on 24 August 2021).
- Breteau, P. Visualisez le réchauffement climatique en France et dans votre ville avec nos «rayures de réchauffement». Le Monde Publ. Du 06 Janvier 2021. Available online: https://www.lemonde.fr/les-decodeurs/article/2021/01/06/visualisez-le-rechauffement-climatique-en-france-et-dans-votre-ville-avec-nos-barres-de-rechauffement_6065388_4355770.html (accessed on 23 August 2021).
- Breteau, P. En France, 2019 a Eté la Troisième Année la Plus Chaude: Visualisez la Hausse Dans Votre Ville. Le Monde. 3 January 2020. Available online: https://www.lemonde.fr/les-decodeurs/article/2020/01/03/2019-est-la-troisieme-annee-la-plus-chaude-en-france-visualisez-la-hausse-dans-votre-ville_6024699_4355770.html (accessed on 23 August 2021).
- Cheikhrouhou, F.; Ammar, R.; Bellili, S.; Jdidi, I.; Bahloul, M.; Ayadi, A.; Bouaziz, M. Paludisme grave d’aéroport à Plasmodium falciparum compliquant un syndrome d’activation macrophagique: A propos d’un cas. La Tunis. Méd. 2019, 97, 379–382. [Google Scholar]
- Craig, M.H.; Kleinschmidt, I.; Nawn, J.B.; Le Sueur, D.; Sharp, B.L. Exploring 30 years of malaria case data in KwaZulu-Natal, South Africa: Part I. The impact of climatic factors. Trop. Med. Int. Health 2004, 9, 1247–1257. [Google Scholar] [CrossRef]
- État de la Population Mondiale. 2011. Available online: https://reliefweb.int/report/world/%C3%A9tat-de-la-population-mondiale-2011-7-milliards-de-personnes-leur-monde-leurs (accessed on 24 August 2021).
- Organisation de l’Aviation Civile Internationale (OACI). Rapport sur Les Mesures de Désinsectisation des Aéronefs et de Lutte Antivectorielle. 2019. Available online: https://www.icao.int/Meetings/a40/Documents/WP/wp_005_fr.pdf (accessed on 26 August 2021).
- Parr, J.B.; Verity, R.; Doctor, S.M.; Janko, M.; Carey-Ewend, K.; Turman, B.J.; Keeler, C.; Slater, H.C.; Whitesell, A.N.; Mwandagalirwa, K.; et al. Pfhrp2-Deleted Plasmodium falciparum parasites in the Democratic Republic of the Congo: A national cross-sectional survey. J. Infect. Dis. 2017, 216, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Golassa, L.; Messele, A.; Amambua-Ngwa, A.; Swedberg, G. High prevalence and extended deletions in Plasmodium falciparum Hrp2/3 genomic loci in Ethiopia. PLoS ONE 2020, 15, e0241807. [Google Scholar] [CrossRef]
- Berhane, A.; Anderson, K.; Mihreteab, S.; Gresty, K.; Rogier, E.; Mohamed, S.; Hagos, F.; Embaye, G.; Chinorumba, A.; Zehaie, A.; et al. Major Threat to malaria control programs by Plasmodium falciparum lacking Histidine-Rich Protein 2, Eritrea. Emerg. Infect. Dis. 2018, 24, 462–470. [Google Scholar] [CrossRef] [Green Version]
- Kojom, L.P.; Singh, V. Prevalence of Plasmodium falciparum field isolates with deletions in Histidine-Rich Protein 2 and 3 genes in Context with Sub-Saharan Africa and India: A systematic review and meta-analysis. Malar. J. 2020, 19, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambou, C. Paludisme du Retour: Une Anthropologie du Risque Palustre Chez Les Voyageurs Migrants Originaires D’Afrique Subsaharienne de Bordeaux. Ph.D. Thesis, Université de Bordeaux, Bordeaux, France, 2018; 334p. Available online: https://tel.archives-ouvertes.fr/tel-02388499/document (accessed on 15 October 2021).
Figure 1.
PRISMA chart showing the steps for the selection of articles.

Figure 2.
Countries in which confirmed or probable cases of airport malaria have been reported in 2006 [6].
Figure 2.
Countries in which confirmed or probable cases of airport malaria have been reported in 2006 [6].

Figure 3.
Airport malaria: period of the year in Europe when the greatest number of infections occurs [6,8,25,26,86,87,88].

Figure 4.
Airport malaria in the European Union: number of cases from 1969 to 2020 per 10-year period [9,25,26,28,86].

Figure 5.
Flow chart showing the process of biological diagnosis of imported malaria [35].
Figure 5.
Flow chart showing the process of biological diagnosis of imported malaria [35].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Articles included in the review.
| Articles Included (n = 27) | Thematic | References |
|---|---|---|
| 11/27 | Vectors bionomics, incrimination, and importation from endemic areas | [6,9,15,16,17,18,19,20,21,22,23,24] |
| 7/27 | Airport malaria epidemiological profiles | [24,25,26,27,28,29,30] |
| 5/27 | Diagnosis of Plasmodium spp. infections | [31,32,33,34,35] |
| 3/27 | Case management and treatment | [36,37,38] |
| 7/27 | Airport malaria mass prevention strategies | [9,22,23,24,25,26,28,39,40] |
| Antimalarial | Treatment Line | Advantages | Disadvantages |
|---|---|---|---|
| Artenimol + Piperaquine | 1st line | - Rapid parasitic clearance - Short treatment - Simple dosage - Taken on an empty stomach - General tolerance | - Conduction disorder - Drug interactions (CYP3A4 inhibitor) |
| Artemether + Lumefantrine | 1st line | - Rapid parasitic clearance - Short treatment - General tolerance | - Low bioavailability (Lumefantrine) - Conduction disorders - Drug interaction (CYP3A4 inhibitor) |
| Atovaquone + Proguanil | 2nd line | - Short treatment - General tolerance - Generics | - Vomiting - Low bioavailability |
| Quinine | 3rd line | - Possible if pregnancy | - Medium tolerance - Long treatment |
CYP3A4: Cytochrome P450 3A4.
Table 3.
Antimalarial drugs for the treatment of childhood malaria [35].
Table 3.
Antimalarial drugs for the treatment of childhood malaria [35].
| Antimalarial | Treatment Line | Advantages | Disadvantages |
|---|---|---|---|
| Artemether + Lumefantrine | 1st line | - Rapid parasitic clearance - General tolerance | - QT interval prolongation without clinical translation - No suitable formulation for infants and small children - Prolonged duration of treatment |
| Artenimol + Piperaquine | 1st line | - Rapid parasitic clearance - General tolerance - Taken on an empty stomach | - QT interval prolongation without clinical translation - No suitable formulation for infants and small children |
| Atovaquone + proguanil | 2nd line | Cardiac tolerance | - No suitable dosage form for infants and small children - Prolonged duration of treatment - Digestive intolerance |
| Mefloquine | 2nd line | - One socket per day - Cardiac tolerance | - No suitable dosage form for infants and small children - Digestive intolerance |
| Quinine | 3rd line | Decline in use | - Cinchonism - Risk of intoxication - Prolonged duration of treatment |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alenou, L.D.; Etang, J. Airport Malaria in Non-Endemic Areas: New Insights into Mosquito Vectors, Case Management and Major Challenges. Microorganisms 2021, 9, 2160. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102160
AMA Style
Alenou LD, Etang J. Airport Malaria in Non-Endemic Areas: New Insights into Mosquito Vectors, Case Management and Major Challenges. Microorganisms. 2021; 9(10):2160. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102160
Chicago/Turabian StyleAlenou, Leo Dilane, and Josiane Etang. 2021. "Airport Malaria in Non-Endemic Areas: New Insights into Mosquito Vectors, Case Management and Major Challenges" Microorganisms 9, no. 10: 2160. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102160
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.