
Occurrence of Hybrid Diarrhoeagenic Escherichia coli Associated with Multidrug Resistance in Environmental Water, Johannesburg, South Africa


Abstract
:1. Introduction
2. Materials and Methods
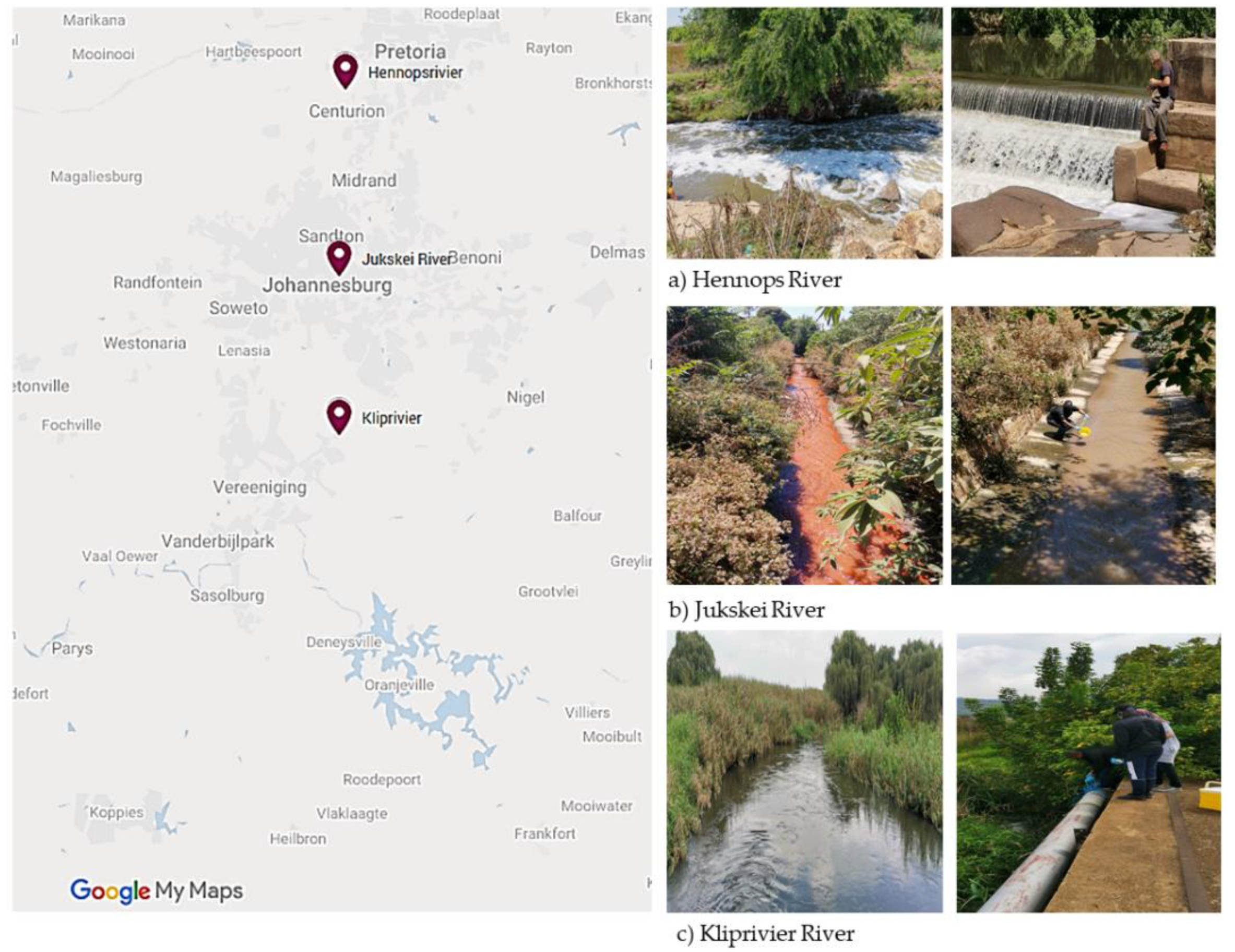
2.1. Ethical Consideration and Sample Collection
2.2. Bacterial Isolation
2.3. DNA Extraction from Escherichia coli Isolates
2.4. Multiplex PCR for the Detection of Virulence Profile of Escherichia coli
2.5. Singleplex PCR for the Confirmation of Escherichia coli
2.6. Visualisation of PCR Products
2.7. Control Strains and Data Analysis
2.8. Antibiotic Susceptibility Testing of Escherichia coli Isolates
3. Results
3.1. Isolation and Detection of E. coli Isolates from Environmental Water Samples
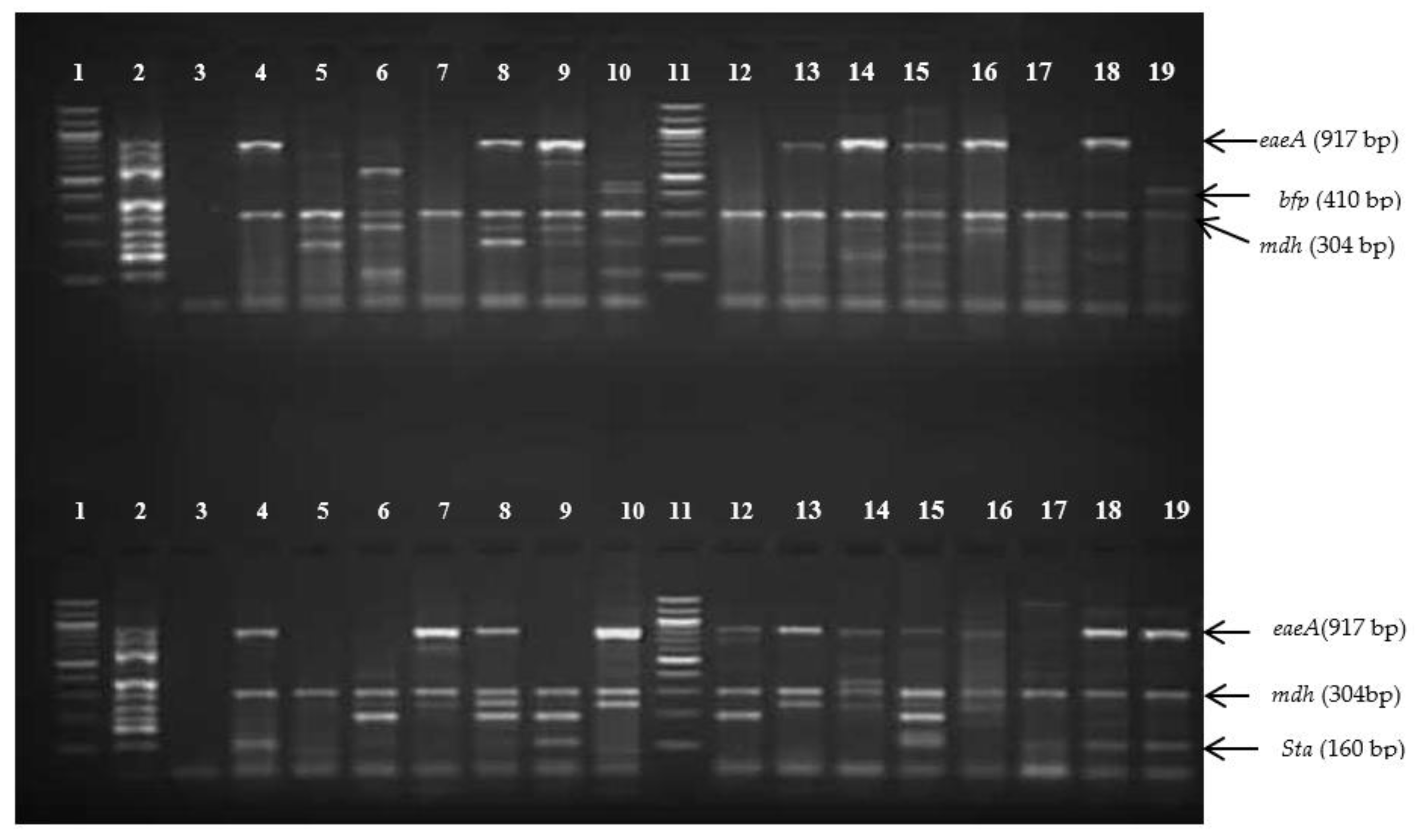
3.2. Detection of Pathogenic Types of E. coli Isolates by Multiplex PCR
3.2.1. Escherichia coli (Single Pathogenic Type) Detected in This Study
3.2.2. Hybrid Pathogenic Types (Two Pathotypes) Detected in This Study
3.2.3. Hybrid Pathogenic Types (Three Pathotypes) Detected in This Study
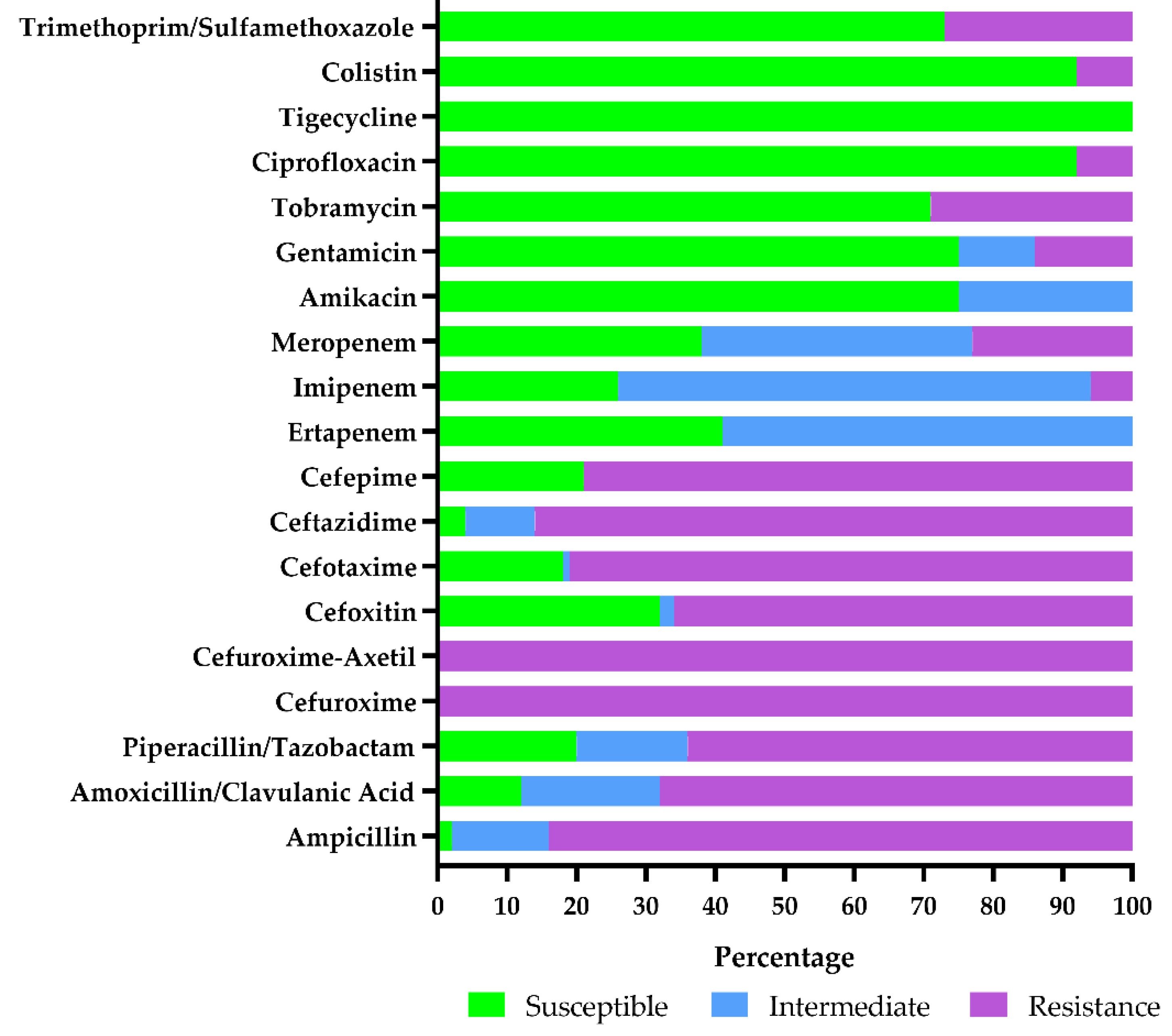
3.2.4. Antimicrobial Susceptibility Testing Results of Escherichia coli Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khalil, I.A.; Troeger, C.; Blacker, B.F.; Rao, P.C.; Brown, A.; Atherly, D.E.; Brewer, T.G.; Engmann, C.M.; Houpt, E.R.; Kang, G.; et al. Morbidity and mortality due to shigella and enterotoxigenic Escherichia coli diarrhoea: The Global Burden of Disease Study 1990–2016. Lancet Infect. Dis. 2018, 18, 1229–1240. [Google Scholar] [CrossRef] [Green Version]
- The United Nations International Children’s Emergency Fund. One Is too Many: Ending Child Deaths from Pneumonia and Diarrhoea; UNICEF: New York, NY, USA, 2016; pp. 1–74. [Google Scholar]
- Bailey, E.S.; Beetsch, N.; Wait, D.A.; Oza, H.H.; Ronnie, N.; Sobsey, M.D. Methods, protocols, guidance and standards for performance evaluation for point-of-use water treatment technologies: History, current status, future needs and directions. Water 2021, 13, 1094. [Google Scholar] [CrossRef]
- Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The population genetics of commensal Escherichia coli. Nat. Rev. Microbiol. 2010, 8, 207–217. [Google Scholar] [CrossRef]
- Wolny-Koładka, K.A.; Zdaniewicz, M. Antibiotic resistance of Escherichia coli isolated from processing of brewery waste with the addition of bulking agents. Sustainability 2021, 13, 10174. [Google Scholar] [CrossRef]
- Molina, F.; López-Acedo, E.; Tabla, R.; Roa, I.; Rebolo, J.E. Improved detection of Escherichia coli and coliform bacteria by multiplex PCR. BMC Biotechnol. 2015, 15, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokdad, A. Estimates of global, regional and national morbidity, mortality and aetiologies of diarrhoeal diseases: A systematic analysis for the global burden of disease Study 2015. Lancet Infect. Dis. 2017, 17, 909–948. [Google Scholar]
- Park, J.; Kim, J.S.; Kim, S.; Shin, E.; Oh, K.-Y.; Kim, Y.; Kim, C.H.; Hwang, M.A.; Jin, C.M.; Na, K.; et al. A waterborne outbreak of multiple diarrhoeagenic Escherichia coli infections associated with drinking water at a school camp. Int. J. Infect. Dis. 2018, 66, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toma, C.; Lu, Y.; Higa, N.; Nakasone, N.; Chinen, I.; Baschkier, A.; Rivas, M.; Iwanaga, M. Multiplex PCR assay for identification of human diarrhoeagenic Escherichia coli. J. Clin. Microbiol. 2003, 41, 2669–2671. [Google Scholar] [CrossRef] [Green Version]
- Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 26, 822–880. [Google Scholar]
- Chique, C.; Hynds, P.; Burke, L.P.; Morris, D.; Ryan, M.P.; O’Dwyer, J. Contamination of domestic groundwater systems by verotoxigenic Escherichia coli (VTEC), 2003–2019: A global scoping review. Water Res. 2021, 188, 116496. [Google Scholar] [CrossRef]
- Cabal, A.; García-Castillo, M.; Cantón, R.; Gortázar, C.; Domínguez, L.; Álvarez, J. Prevalence of Escherichia coli virulence genes in patients with diarrhoea and a subpopulation of healthy volunteers in Madrid, Spain. Front. Microbiol. 2016, 7, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Zhang, J.; Ambikan, A.; Jernberg, C.; Ehricht, R.; Scheutz, F.; Xiong, Y.; Matussek, A. Molecular characterisation and comparative genomics of clinical hybrid Shiga toxin-producing and enterotoxigenic Escherichia coli (STEC/ETEC) strains in Sweden. Nat. Sci. Rep. 2019, 9, 5619. [Google Scholar] [CrossRef] [Green Version]
- Gioia-Di Chiacchio, R.M.; Cunha, M.; de Sá, L.; Davies, Y.M.; Pereira, C.; Martins, F.H.; Munhoz, D.D.; Abe, C.M.; Franzolin, M.R.; Dos Santos, L.F.; et al. Novel Hybrid of Typical Enteropathogenic Escherichia coli and Shiga-Toxin-Producing E. coli (tEPEC/STEC) Emerging from Pet Birds. Front. Microbiol. 2018, 9, 2975. [Google Scholar] [CrossRef]
- García, A.; Fox, J.G.; Besser, T.E. Zoonotic enterohemorrhagic Escherichia coli: A One Health perspective. ILAR J. 2010, 51, 221–232. [Google Scholar] [CrossRef]
- Lan, R.; Alles, M.C.; Donohoe, K.; Martinez, M.B.; Reeves, P.R. Molecular evolutionary relationships of enteroinvasive Escherichia coli and Shigellaspp. Infect. Immun. 2004, 72, 5080–5088. [Google Scholar] [CrossRef] [Green Version]
- Newitt, S.; MacGregor, V.; Robbins, V.; Bayliss, L.; Chattaway, M.A.; Dallman, T.; Ready, D.; Aird, H.; Puleston, R.; Hawker, J. Two Linked Enteroinvasive Escherichia coli Outbreaks, Nottingham, UK, June 2014. Emerg. Infect. Dis. 2016, 22, 1178–1184. [Google Scholar] [CrossRef] [Green Version]
- Nakhjavani, F.A.; Emaneini, M.; Hosseini, H.; Iman-Eini, H.; Aligholi, M.; Jabalameli, F.; Haghi-Ashtiani, M.T.; Taherikalani, M.; Mirsalehian, A. Molecular analysis of typical and atypical enteropathogenic Escherichia coli (EPEC) isolated from children with diarrhoea. J. Med. Microbiol. 2013, 62, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Hazen, T.H.; Michalski, J.; Luo, Q.; Shetty, A.C.; Daugherty, S.C.; Fleckenstein, J.M.; Rasko, D.A. Comparative genomics and transcriptomics of Escherichia coli isolates carrying virulence factors of both enteropathogenic and enterotoxigenic Escherichia coli. Sci. Rep. 2017, 7, 3513. [Google Scholar] [CrossRef] [PubMed]
- Leonard, S.R.; Mammel, M.K.; Rasko, D.A.; Lacher, D.W. Hybrid Shiga Toxin-Producing and Enterotoxigenic Escherichia sp. Cryptic Lineage 1 Strain 7v Harbors a Hybrid Plasmid. Appl. Environ. Microbiol. 2016, 82, 4309–4319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, S.; Pazhani, G.P.; Nataro, J.P.; Ramamurthy, T. Heterogenic virulence in a diarrheagenic Escherichia coli: Evidence for an EPEC expressing heat-labile toxin of ETEC. Int. J. Med. Microbiol. 2015, 305, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Santos, F.F.; Silva, R.M.; Gomes, T. Diversity of Hybrid- and Hetero-Pathogenic Escherichia coli and their potential implication in more severe diseases. Front. Cell. Infect. Microbiol. 2020, 10, 339. [Google Scholar] [CrossRef]
- Aref, N.M.; Abdel-Raheem, A.A.; Kamaly, H.F.; Hussien, S.Z. Clinical and sero-molecular characterization of Escherichia coli with an emphasis on hybrid strain in healthy and diarrheic neonatal calves in Egypt. Open Vet. J. 2018, 8, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Bai, X.; Zhang, J.; Sun, H.; Fu, S.; Fan, R.; He, X.; Scheutz, F.; Matussek, A.; Xiong, Y. Escherichia coli strains producing a novel Shiga toxin 2 subtype circulate in China. Int. J. Med. Microbiol. 2020, 310, 151377. [Google Scholar] [CrossRef]
- Bielaszewska, M.; Mellmann, A.; Zhang, W.; Köck, R.; Fruth, A.; Bauwens, A.; Peters, G.; Karch, H. Characterisation of the Escherichia coli strain associated with an outbreak of haemolytic uraemic syndrome in Germany: A microbiological study. Lancet Infect. Dis. 2011, 11, 671–676. [Google Scholar] [CrossRef] [Green Version]
- The Centers for Disease Control and Prevention. The Antibiotic Resistance Threats in the United States. 2019. Available online: www.cdc.gov/DrugResistance/Biggest-Threats.html (accessed on 22 June 2021).
- Wolny-Koładka, K.A.; Lenart-Boroń, A. Phenotypic and molecular assessment of drug resistance profile and genetic diversity of waterborne Escherichia coli. Water Air Soil Pollut. 2016, 227, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lamprecht, C.; Romanis, M.; Huisamen, N.; Carinus, A.; Schoeman, N.; Sigge, G.O.; Britz, T.J. Escherichia coli with virulence factors and multidrug resistance in the Plankenburg River. S. Afr. J. Sci. 2014, 110, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Morel, C. Transmission of antimicrobial resistance from livestock agriculture to humans and from humans to animals. In Food, Agriculture and Fisheries Papers, No. 133; OECD Publishing: Paris, France, 2019. [Google Scholar] [CrossRef]
- Msolo, L.; Iweriebor, B.C.; Okoh, A.I. Antimicrobial resistance profiles of diarrhoeagenic E. coli (DEC) and Salmonella species recovered from diarrhoeal patients in selected rural communities of the Amathole District Municipality, Eastern Cape province, South Africa. Infect. Drug Resist. 2020, 13, 4615–4626. [Google Scholar] [CrossRef] [PubMed]
- Wolny-Koładka, K.A.; Lenart-Boroń, A. Antimicrobial resistance and the presence of extended-spectrum beta-lactamase genes in Escherichia coli isolated from the environment of horse riding centers. Environ. Sci. Pollut. Res. 2018, 25, 21789–21800. [Google Scholar] [CrossRef] [PubMed]
- Omar, K.B.; Barnard, T.G. Detection of diarrhoeagenic Escherichia coli in clinical and environmental water sources in South Africa using single-step 11-gene m-PCR. World J. Microbiol. Biotechnol. 2014, 30, 2663–2671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mbene, A.B.; Houreld, N.N.; Abrahamse, H. DNA damage after phototherapy in wounded fibroblast cells irradiated with 16 J/cm2. J. Photochem. Photobiol. 2009, 94, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Pass, M.A.; Odedra, R.; Batt, R.M. Multiplex PCRs for identification of Escherichia coli virulence genes. J. Clin. Microbiol. 2000, 38, 2001–2004. [Google Scholar] [CrossRef] [Green Version]
- Clinical Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. M100-S22, M2–7 and M–7; Clinical Laboratory Standards Institute: Wayne, PA, USA, 2021. [Google Scholar]
- Potgieter, N.; Karambwe, S.; Mudau, L.S.; Barnard, T.G.; Traore, A. Human enteric pathogens in eight rivers used as rural household drinking water sources in the northern region of South Africa. Int. J. Environ. Res. Public Health 2020, 17, 2079. [Google Scholar] [CrossRef] [Green Version]
- Cooley, M.B.; Jay-Russell, M.; Atwill, E.R.; Carychao, D.; Nguyen, K.; Quinones, B.; Patel, R.; Walker, S.; Swimley, M.; Pierre-Jerome, E.; et al. Development of a robust method for isolation of Shiga toxin-positive Escherichia coli from faecal, plant, soil and water samples from a leafy greens production region in California. PLoS ONE 2013, 8, e65716. [Google Scholar] [CrossRef] [Green Version]
- Lim, M.A.; Kim, J.Y.; Acharya, D.; Bajgain, B.B.; Park, J.H.; Yoo, S.J.; Lee, K. A diarrhoeagenic enteropathogenic Escherichia coli (EPEC) infection outbreak that occurred among elementary school children in Gyeongsangbuk-Do province of South Korea was associated with consumption of water-contaminated food items. Int. J. Environ. Res. Public Health 2020, 17, 3149. [Google Scholar] [CrossRef]
- Pienaar, J.A.; Singh, A.; Barnard, T.G. Acid-happy: Survival and recovery of enteropathogenic Escherichia coli (EPEC) in simulated gastric fluid. Microb. Pathog. 2019, 128, 396–404. [Google Scholar] [CrossRef]
- Ndlovu, T.; Le Roux, M.; Khan, W.; Khan, S. Co-detection of virulent Escherichia coli genes in surface water sources. PLoS ONE 2015, 10, e0116808. [Google Scholar] [CrossRef]
- Sidhu, J.P.S.; Ahmed, W.; Hodgers, L.; Toze, S. Occurrence of virulence genes associated with diarrhoeagenic pathotypes in Escherichia coli isolates from surface water. Appl. Environ. Microbiol. 2013, 79, 328–335. [Google Scholar] [CrossRef] [Green Version]
- Traoré, A.N.; Mulaudzi, K.; Chari, G.J.E.; Foord, S.H.; Mudau, L.S.; Barnard, T.G.; Potgieter, N. The impact of human activities on microbial quality of rivers in the Vhembe District, South Africa; Int. J. Environ. Res. Public Health 2016, 13, 817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johura, F.T.; Parveen, R.; Islam, A.; Sadique, A.; Rahim, M.N.; Monira, S.; Khan, A.R.; Ahsan, S.; Ohnishi, M.; Watanabe, H.; et al. Occurrence of hybrid Escherichia coli strains carrying Shiga toxin and heat-stable toxin in livestock of Bangladesh. Front. Public Health 2017, 4, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nontongana, N.; Sibanda, T.; Ngwenya, E.; Okoh, A.I. Prevalence and antibiogram profiling of Escherichia coli pathotypes isolated from the Kat River and the Fort Beaufort Abstraction Water. Int. J. Environ. Res. Public Health 2014, 11, 8213–8227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canizalez-Roman, A.; Velazquez-Roman, J.; Valdez-Flores, M.A.; Flores-Villaseñor, H.; Vidal, J.E.; Muro-Amador, S.; Guadrón-Llanos, A.M.; Gonzalez-Nuñez, E.; Medina-Serrano, J.; Tapia-Pastrana, G.; et al. Detection of antimicrobial-resistance diarrheagenic Escherichia coli strains in surface water used to irrigate food products in the northwest of Mexico. Int. J. Food Microbiol. 2019, 304, 1–10. [Google Scholar] [CrossRef]
- Aijuka, M.; Santiago, A.E.; Giron, J.A.; Nataro, J.P.; Buys, E.M. Enteroaggregative Escherichia coli is the predominant diarrhoeagenic E. coli pathotype among irrigation water and food sources in South Africa. Int. J. Food Microbiol. 2018, 278, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Tanih, N.F.; Bolick, D.T.; Samie, A.; Nyathi, E.; Dillingham, R.; Pinkerton, R.C.; Guerrant, R.L.; Bessong, P.O. Prevalence of virulence genes in Enteroaggregative E. coli isolates from young children from rural South Africa. Am. J. Trop. Med. Hyg. 2019, 101, 1027–1033. [Google Scholar] [CrossRef]
- Mbanga, J.; Amoako, D.G.; Abia, A.L.K.; Allam, M.; Ismail, A.; Essack, S.Y. Genomic insights of multidrug-resistant Escherichia coli from wastewater sources and their association with clinical pathogens in South Africa. Front. Vet. Sci. 2021, 8, 636715. [Google Scholar] [CrossRef]
- Ramlal, P.S.; Kistnasamy, E.J. An ecological study of diarrhoeagenic Escherichia coli associated with indiscriminate waste dumps and under five diarrhoea in six informal settlements in Durban, South Africa. Int. J. Environ. Waste Manag. 2017, 20, 300–323. [Google Scholar] [CrossRef]
- Cho, S.; Hiott, L.M.; Barrett, J.B.; McMillan, E.A.; House, S.L.; Humayoun, S.B.; Adams, E.S.; Jackson, C.R.; Frye, J.G. Prevalence and characterization of Escherichia coli isolated from the upper Oconee watershed in Northeast Georgia. PLoS ONE 2018, 13, e0197005. [Google Scholar] [CrossRef] [Green Version]
- Bonkoungou, I.J.O.; Somda, N.S.; Traoré, O.; Zoma, B.S.; Garba, Z.; Drabo, K.M.; Barro, N. Detection of diarrhoeagenic Escherichia coli in human diarrhoeic stool and drinking water samples in Ouagadougou, Burkina Faso. Afr. J. Infect. Dis. 2021, 15, 53–58. [Google Scholar] [PubMed]
- Nyholm, O.; Heinikainen, S.; Pelkonen, S.; Hallanvuo, S.; Haukka, K.; Siitonen, A. Hybrids of Shigatoxigenic and Enterotoxigenic Escherichia coli (STEC/ETEC) among human and animal isolates in Finland. Zoonoses Public Health 2015, 62, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Patzi-Vargas, S.; Zaidi, M.B.; Perez-Martinez, I.; León-Cen, M.; Michel-Ayala, A.; Chaussabel, D.; Estrada-Garcia, T. Diarrheagenic Escherichia coli carrying supplementary virulence genes are an important cause of moderate to severe diarrhoeal disease in Mexico. PLoS Negl. Trop. Dis. 2015, 9, e0003510. [Google Scholar] [CrossRef] [PubMed]
- Badouei, M.A.; Morabito, S.; Najafifar, A.; Mazandarani, E. Molecular characterisation of enterohemorrhagic Escherichia coli haemolysin gene (EHEC-hlyA)-harboring isolates from cattle reveals a diverse origin and hybrid diarrheagenic strains. Infect. Genet. Evol. 2016, 39, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Antikainen, J.; Tarkka, E.; Haukka, K.; Siitonen, A.; Vaara, M.; Kirveskari, J. The new 16-plex PCR method for rapid detection of diarrhoeagenic Escherichia coli directly from stool samples. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 899–908. [Google Scholar] [CrossRef] [PubMed]
- The World Health Organization. Antibiotic Resistance. 2020. Available online: www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 26 June 2021).
- The World Health Organization. Global Antimicrobial Resistance Surveillance System (GLASS) Report. Early Implementation. 2021. Available online: www.who.int/publications/i/item/9789240027336 (accessed on 14 September 2021).
- Bezuidenhout, C.C.; O’Reilly, G.; Sigudu, M.V.; Ncube, E.J. A Scoping Study on the Levels of Antimicrobials and Presence of Antibiotic Resistant Bacteria in Drinking Water; WRC Report No. KV 360/16; Water Research Commission: Lynnwood, South Africa, 2016; ISBN 978-1-4312-0823-4. [Google Scholar]
- Hogan, C.A.; Watz, N.; Budvytiene, I.; Banaei, N. Rapid antimicrobial susceptibility testing by VITEK®2 directly from blood cultures in patients with gram-negative rod bacteremia. Diagnost. Microbiol. Infect. Dis. 2019, 94, 116–121. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Pathogen | Gene Primers | Primer Sequence (5′– 3′) | Size (bp) | Reference |
|---|---|---|---|---|
| Internal control | mdh (F) | GGT ATG GAT CGT TCC GAC CT | 304 | [32] |
| mdh (R) | GGC AGA ATG GTA ACA CCA GAG T | |||
| External control | gapdh (F) | GAG TCA ACG GAT TTG GTC GT | 238 | [33] |
| gapdh (R) | TTG ATT TTG GAG GGA TCT CG | |||
| EIEC | ial (F) | GGT ATG ATG ATG ATG AGT CCA | 650 | [34] |
| ial (R) | GGA GGC CAA CAA TTA TTT CC | |||
| EHEC | stx1 (F) | ACA CTG GAT GAT CTC AGT GG | 614 | [32] |
| stx1 (R) | CTG AAT CCC CCT CCA TTA TG | |||
| stx2 (F) | CCA TGA CAA CGG ACA GCA GTT | 779 | ||
| stx2 (R) | CCT GTC AAC TGA GCA CTT TG | |||
| EHEC/aEPEC | eaeA (F) | CTG AAC GGC GAT TAC GCG AA | 917 | [34] |
| eaeA (R) | CCA GAC GAT ACG ATC CAG | |||
| tEPEC | bfpA (F) | AAT GGT GCT TGC GCT TGC TGC | 410 | |
| bfpM (R) | TAT TAA CAC CGT AGC CTT TCG CTG AAG TAC CT | |||
| EAEC | eagg (F) | AGA CTC TGG CGA AAG ACT GTA TC | 194 | [34] |
| eagg (R) | ATG GCT GTC TGT AAT AGA TGA GAA C | |||
| ETEC | lt-1 (F) | TGG ATT CAT CAT GCA CCA CAA GG | 360 | [34] |
| lt-1 (R) | CCA TTT CTC TTT TGC CTG CCA TC | |||
| sta (F) | TTT CCC CTC TTT TAG TCA GTC AAC TG | 160 | ||
| sta(R) | GGC AGG ATT ACA ACA AAG TTC ACA | |||
| E. coli toxin | astA (F) | GCC ATC AAC ACA GTA TAT CC | 106 | [32] |
| astA (R) | GAG TGA CGG CTT TGT AGT C |
| Pathogenic Types | Genes Detected | n (170) | % |
|---|---|---|---|
| ComEC | mdh | 68 | 40 |
| EPEC | 32 | 19.2 | |
| aEPEC | eaeA | (30) | (18.0) |
| tEPEC | bfp | (1) | (0.6) |
| bfp + eaeA | (1) | (0.6) | |
| ETEC | 19 | 11.4 | |
| lt | (2) | (1.2) | |
| sta | (15) | (9.0) | |
| lt + sta | (2) | (1.2) | |
| EHEC | 5 | 3.0 | |
| stx1 | (2) | (1.2) | |
| stx2 | (2) | (1.2) | |
| stx1 + stx2 | (1) | (0.6) | |
| EAEC | eagg | 10 | 6.0 |
| EIEC | ial | 0 | 0.0 |
| Hybrid Pathotypes | Gene Combinations Detected | n (170) | % |
|---|---|---|---|
| EAEC/aEPEC | eaeA + eagg | 13 | 7.6 |
| EAEC/ETEC | 5 | 3.0 | |
| eagg + sta | (3) | (1.8) | |
| eagg + lt + sta | (2) | (1.2) | |
| EPEC/ETEC | 4 | 2.4 | |
| eaeA + lt | (1) | (0.6) | |
| eaeA + lt + sta | (3) | (1.8) | |
| EHEC/ETEC | 3 | 1.8 | |
| Stx1 + sta | (1) | (0.6) | |
| Stx2 + sta | (1) | (0.6) | |
| Stx1 + lt + sta | (1) | (0.6) | |
| EPEC/EHEC | 3 | 1.8 | |
| aEPEC/EHEC | eaeA + stx2 | (2) | (1.2) |
| tEPEC/EHEC | eaeA + bfp + stx1 + stx2 | (1) | (0.6) |
| Hybrid Pathotypes | Gene Combinations Detected | n (170) | % |
|---|---|---|---|
| EAEC/aEPEC/ETEC | eagg + eaeA + sta | 4 | 2.4 |
| EAEC/EHEC/ETEC | eagg + stx1 + sta | 2 | 1.2 |
| EHEC/ETEC/aEPEC | 3 | 1.8 | |
| eaeA + stx1 + sta | (1) | (0.6) | |
| eaeA + stx1 + stx2 +lt + sta | (2) | (1.2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolukaoto, J.Y.; Singh, A.; Alfinete, N.; Barnard, T.G. Occurrence of Hybrid Diarrhoeagenic Escherichia coli Associated with Multidrug Resistance in Environmental Water, Johannesburg, South Africa. Microorganisms 2021, 9, 2163. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102163
Bolukaoto JY, Singh A, Alfinete N, Barnard TG. Occurrence of Hybrid Diarrhoeagenic Escherichia coli Associated with Multidrug Resistance in Environmental Water, Johannesburg, South Africa. Microorganisms. 2021; 9(10):2163. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102163
Chicago/Turabian StyleBolukaoto, John Y., Atheesha Singh, Ntando Alfinete, and Tobias G. Barnard. 2021. "Occurrence of Hybrid Diarrhoeagenic Escherichia coli Associated with Multidrug Resistance in Environmental Water, Johannesburg, South Africa" Microorganisms 9, no. 10: 2163. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102163