
Evidence of HIV-1 Genital Shedding after One Year of Antiretroviral Therapy in Females Recently Diagnosed in Bamako, Mali

 , ,
, ,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Patients and Methods
2.1. Patients
2.2. Samples
2.3. CD4+ Cell Count
2.4. HIV RNA Viral Load
2.5. Genotyping Resistance Assay
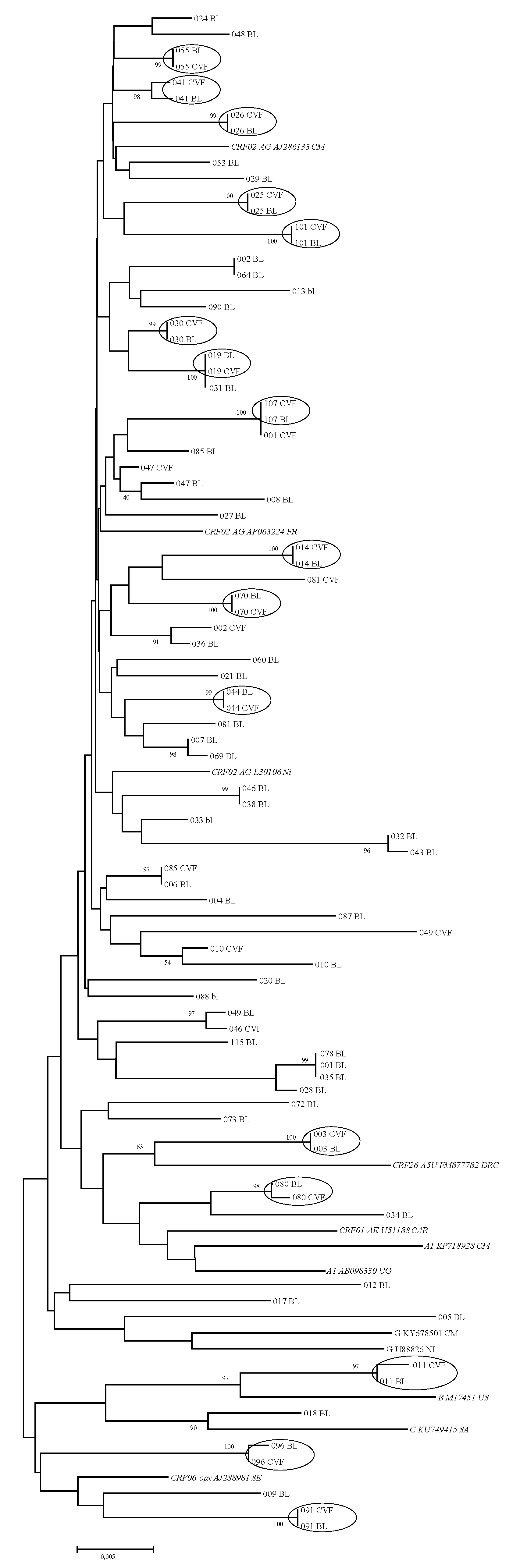
2.6. Phylogenetic Analysis
2.7. Nucleotide Sequence Accession Numbers
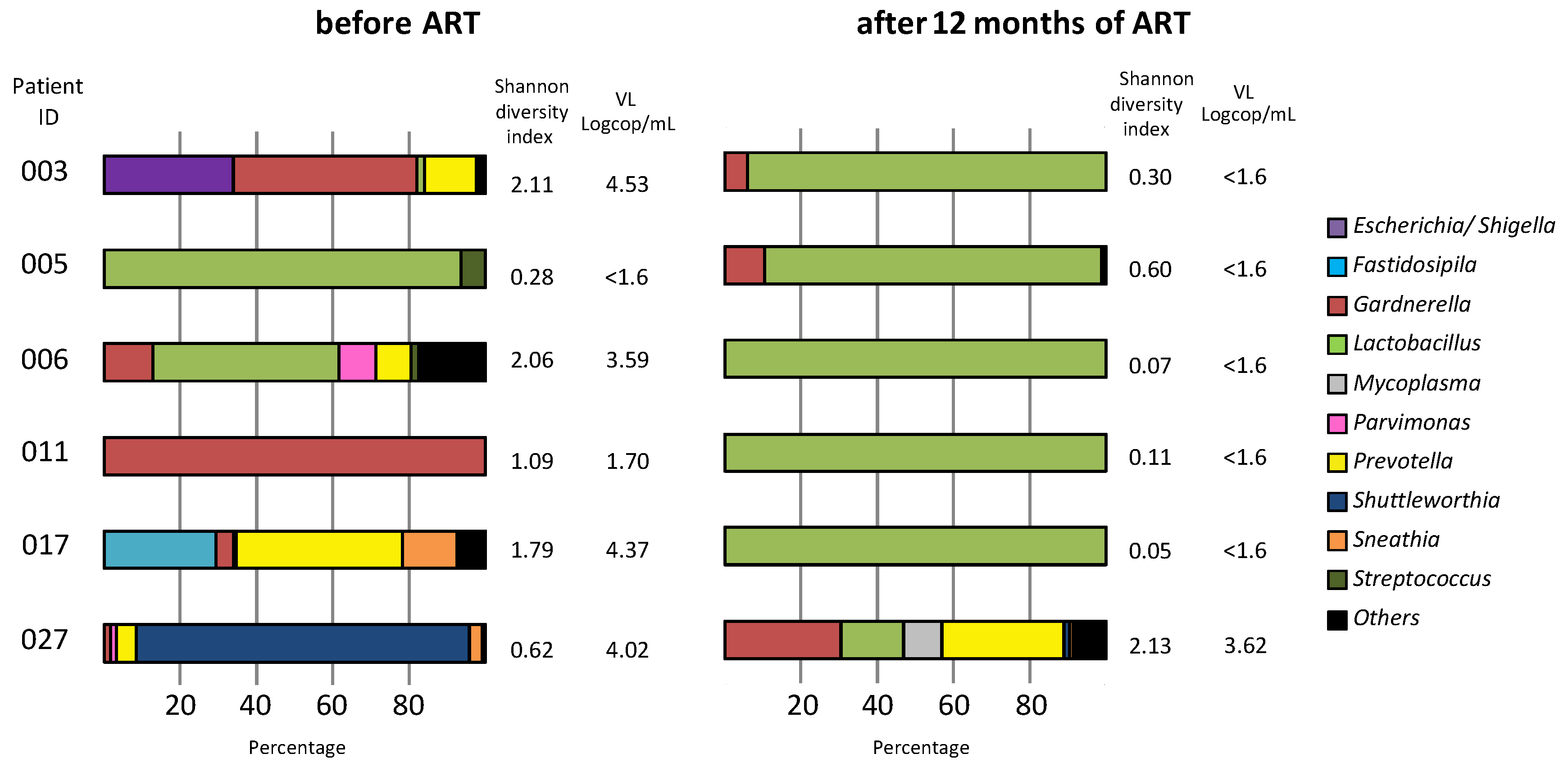
2.8. Vaginal Microbiota Analysis
2.9. Statistical Analysis
3. Results
3.1. Characteristics of the Studied Population at the Time of Diagnosis
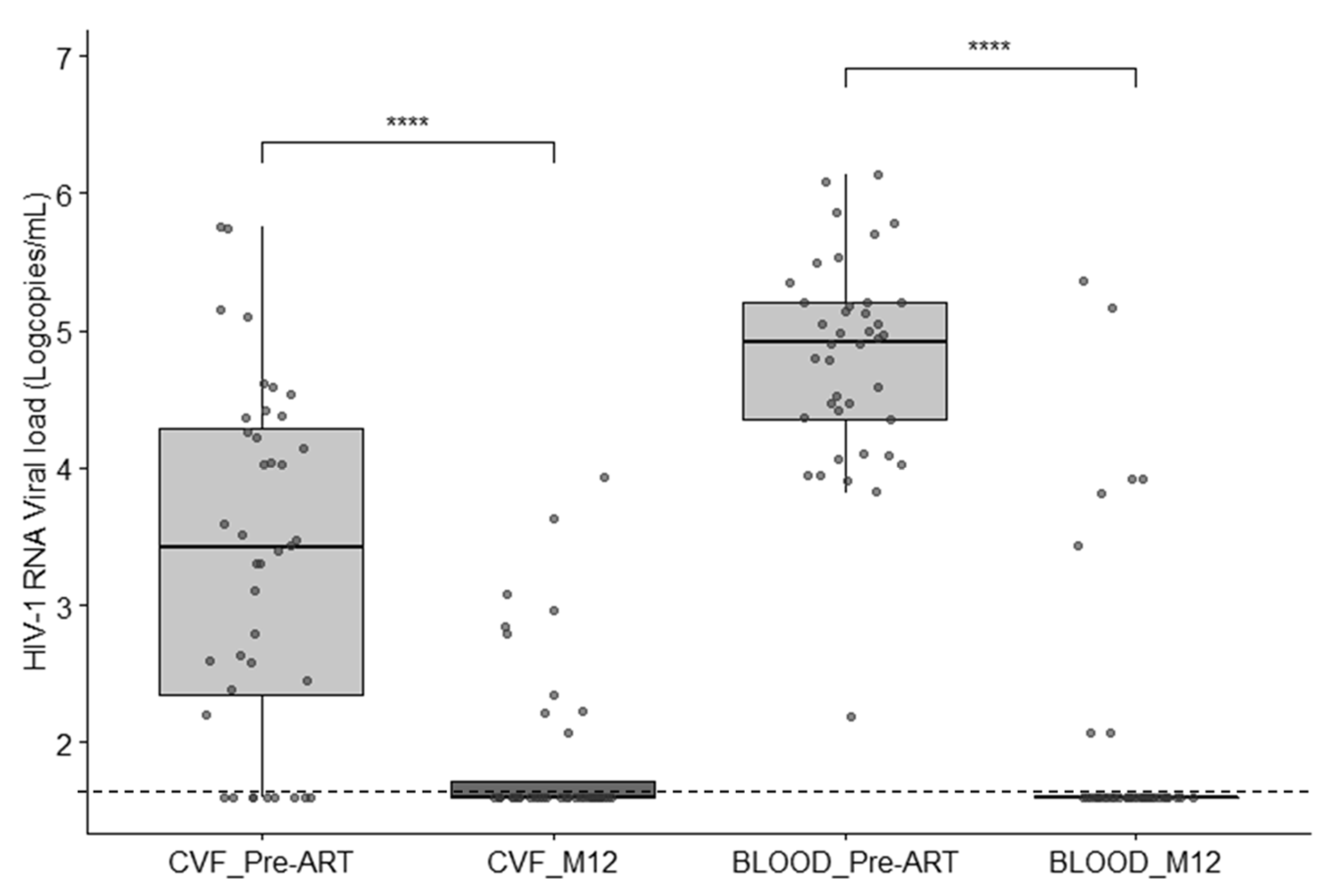
3.2. HIV-1 Viral Load in Blood and Cervicovaginal Fluids
3.3. HIV-1 Subtypes and Phylogenetic Analysis
3.4. Pretreatment Drug Resistance Mutations in Blood and CVF
3.5. Longitudinal Follow-Up after ART Initiation
3.6. Vaginal Microbiota Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations General Assembly. Resolution No A/RES/70/266, Political Declaration on HIV and AIDS: On the Fast-Track to Accelerate the Fight against HIV and to End the AIDS Epidemic by 2030. 2016, pp. 1–26. Available online: https://www.unaids.org/sites/default/files/media_asset/2016-political-declaration-HIV-AIDS_en.pdf (accessed on 20 June 2021).
- World Health Organization. Guideline on When to Start Antiretroviral Therapy and on Pre-Exposure Prophylaxis for HIV, WHO. 2015, pp. 1–78. Available online: http://apps.who.int/iris/bitstream/handle/10665/186275/9789241509565_eng.pdf;jsessionid=1DF46A4F6059616EE796E462BBDDF1EF?sequence=1 (accessed on 19 June 2021).
- UNAIDS Data 2020. Available online: https://www.unaids.org/sites/default/files/media_asset/2020_aids-data-book_en.pdf (accessed on 21 June 2021).
- Hamers, R.L.; Sigaloff, K.C.; Kityo, C.; Mugyenyi, P.; de Wit, T.F. Emerging HIV-1 drug resistance after roll-out of antiretroviral therapy in sub-Saharan Africa. Curr. Opin. HIV AIDS 2013, 8, 19–26. [Google Scholar] [CrossRef]
- Aghokeng, A.F.; Monleau, M.; Eymard-Duvernay, S.; Dagnra, A.; Kania, D.; Ngo-Giang-Huong, N.; Toni, T.D.; Touré-Kane, C.; Truong, L.X.; Delaporte, E.; et al. Extraordinary heterogeneity of virological outcomes in patients receiving highly antiretroviral therapy and monitored with the World Health Organization public health approach in sub-saharan Africa and southeast Asia. Clin. Infect. Dis. 2014, 58, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngo-Giang-Huong, N.; Huynh, T.H.K.; Dagnra, A.Y.; Thomas-d’Aquin, T.; Maiga, A.I.; Kania, D.; Eymard-Duvernay, S.; Peeters, M.; Soulie, C.; Peytavin, G.; et al. Prevalence of pretreatment HIV drug resistance in West African and Southeast Asian countries. J. Antimicrob. Chemother. 2019, 74, 462–467. [Google Scholar] [CrossRef]
- Critchlow, C.W.; Kiviat, N.B. Detection of human immunodeficiency virus type 1 and type 2 in the female genital tract: Implications for the understanding of virus transmission. Obstet. Gynecol. Surv. 1997, 52, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.S.; Chen, Y.Q.; McCauley, M.; Gamble, T.; Hosseinipour, M.C.; Kumarasamy, N.; Hakim, J.G.; Kumwenda, J.; Grinsztejn, B.; Pilotto, J.H.; et al. Prevention of HIV-1 infection with early antiretroviral therapy. N. Engl. J. Med. 2011, 365, 493–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, C.E.; Lennox, J.L.; Pratt-Palmore, M.; Wright, T.C.; Schinazi, R.F.; Evans-Strickfaden, T.; Bush, T.J.; Schnell, C.; Conley, L.J.; Clancy, K.A.; et al. Correlation of human immunodeficiency virus type 1 RNA levels in blood and the female genital tract. J. Infect. Dis. 1999, 179, 871–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iversen, A.K.; Larsen, A.R.; Jensen, T.; Fugger, L.; Balslev, U.; Wahl, S.; Gerstoft, J.; Mullins, J.I.; Skinhøj, P. Distinct determinants of human immunodeficiency virus type 1 RNA and DNA loads in vaginal and cervical secretions. J. Infect. Dis. 1998, 177, 1214–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cu-Uvin, S.; DeLong, A.K.; Venkatesh, K.K.; Hogan, J.W.; Ingersoll, J.; Kurpewski, J.; De Pasquale, M.P.; D’Aquila, R.; Caliendo, A.M. Genital tract HIV-1 RNA shedding among women with below detectable plasma viral load. AIDS 2010, 24, 2489–2497. [Google Scholar] [CrossRef] [PubMed]
- Mauck, C.; Chen, P.L.; Morrison, C.S.; Fichorova, R.N.; Kwok, C.; Chipato, T.; Salata, R.A.; Doncel, G.F. Biomarkers of Cervical Inflammation and Immunity Associated with Cervical Shedding of HIV-1. AIDS Res. Hum. Retrovir. 2016, 32, 443–451. [Google Scholar] [CrossRef] [Green Version]
- Brooks, J.P.; Edwards, D.J.; Blithe, D.L.; Fettweis, J.M.; Serrano, M.G.; Sheth, N.U.; Strauss, J.F.; Buck, G.A.; Jefferson, K.K. Effects of combined oral contraceptives, depot medroxyprogesterone acetate and the levonorgestrel-releasing intrauterine system on the vaginal microbiome. Contraception 2017, 95, 405–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdool Karim, S.S.; Baxter, C.; Passmore, J.S.; McKinnon, L.R.; Williams, B.L. The genital tract and rectal microbiomes: Their role in HIV susceptibility and prevention in women. J. Int. AIDS Soc. 2019, 22, e25300. [Google Scholar] [CrossRef] [Green Version]
- Abdool Karim, S.S.; Passmore, J.S.; Baxter, C. The microbiome and HIV prevention strategies in women. Curr. Opin. HIV AIDS 2018, 13, 81–87. [Google Scholar] [CrossRef]
- Baeten, J.M.; Kahle, E.; Lingappa, J.R.; Coombs, R.W.; Delany-Moretlwe, S.; Nakku-Joloba, E.; Mugo, N.R.; Wald, A.; Corey, L.; Donnell, D.; et al. Partners in Prevention HSV/HIV Transmission Study Team. Genital HIV-1 RNA Quantity Predicts Risk of Heterosexual HIV-1 Transmission. Sci. Transl. Med. 2011, 3, 77ra29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keita, A.; Sereme, Y.; Pillet, S.; Coulibaly, S.; Diallo, F.; Pozzetto, B.; Thiero, T.A.; Bourlet, T. Impact of HIV-1 primary drug resistance on the efficacy of a first-line antiretroviral regimen in the blood of newly diagnosed individuals in Bamako, Mali. J. Antimicrob. Chemother. 2019, 74, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Tamura, K.; Jakobsen, I.B.; Nei, M. MEGA 2: Molecular evolutionary genetics analysis software. Bioinformatics 2001, 17, 1244–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes Da Silva, R.K.; Monteiro de Pina Araujo, I.I.; Venegas Maciera, K.; Gonçalves Morgado, M. Genetic Characterization of a New HIV-1 Sub-Subtype A in Cabo Verde, Denominated A8. Viruses 2021, 13, 1093. [Google Scholar] [CrossRef]
- Yan, M.; Pamp, S.J.; Fukuyama, J.; Hwang, P.H.; Cho, D.Y.; Holmes, S.; Relman, D.A. Nasal microenvironments and interspecific interactions influence nasal microbiota complexity and S. aureus carriage. Cell. Host Microbe 2013, 14, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Mori, H.; Maruyama, F.; Kato, H.; Toyoda, A.; Dozono, A.; Ohtsubo, Y.; Nagata, Y.; Fujiyama, A.; Tsuda, M.; Kurokawa, K. Design and experimental application of a novel non-degenerate universal primer set that amplifies prokaryotic 16S rRNA genes with a low possibility to amplify eukaryotic rRNA genes. DNA Res. 2014, 21, 217–227. [Google Scholar] [CrossRef]
- Kone, B.; Maiga, M.; Baya, B.; Sarro, Y.; Coulibaly, N.; Kone, A.; Diarra, B.; Sanogo, M.; Togo, A.; Goita, D.; et al. Establishing Reference Ranges of Hematological Parameters from Malian Healthy Adults. J. Blood Lymph. 2017, 7, 154. [Google Scholar] [CrossRef]
- Theys, K.; Van Laethem, K.; Gomes, P.; Baele, G.; Pineda-Peña, A.C.; Vandamme, A.M.; Camacho, R.J.; Abecasis, A.B. On behalf of the Portuguese HIV-1 Resistance Study Group. Sub-Epidemics Explain Localized High Prevalence of Reduced Susceptibility to Rilpivirine in Treatment-Naive HIV-1-Infected Patients: Subtype and Geographic Compartmentalization of Baseline Resistance Mutations. AIDS Res. Hum. Retrovir. 2016, 32, 427–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvez, V.; Marcelin, A.G.; Vingerhoets, J.; Hill, A.; Hadacek, B.; Moecklinghoff, C. Systematic review to determine the prevalence of transmitted drug resistance mutations to rilpivirine in HIV-infected treatment-naive persons. Antivir. Ther. 2016, 21, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Derache, A.; Traore, O.; Koita, V.; Sylla, A.; Tubiana, R.; Simon, A.; Canestri, A.; Carcelain, G.; Katlama, C.; Calvez, V.; et al. Genetic diversity and drug resistance mutations in HIV type 1 from untreated patients in Bamako, Mali. Antivir. Ther. 2007, 12, 123–129. [Google Scholar]
- Low, A.J.; Konate, I.; Nagot, N.; Weiss, H.A.; Kania, D.; Vickerman, P.; Segondy, M.; Mabey, D.; Pillay, D.; Meda, N.; et al. Cervicovaginal HIV-1 Shedding in Women Taking Antiretroviral Therapy in Burkina Faso: A Longitudinal Study. J. Acquir. Immune Defic. Syndr. 2014, 65, 237–245. [Google Scholar] [CrossRef] [Green Version]
- King, C.C.; Ellington, S.R.; Davis, N.L.; Coombs, R.W.; Pyra, M.; Hong, T.; Mugo, N.; Patel, R.C.; Lingappa, J.R.; Baeten, J.M.; et al. Partners in Prevention HSV/HIV Transmission Study and Partners PrEP Study Teams. Prevalence, Magnitude, and Correlates of HIV-1 Genital Shedding in Women on Antiretroviral Therapy. J. Infect. Dis. 2017, 216, 1534–1540. [Google Scholar] [CrossRef]
- Grignolo, S.; Bruzzone, B.; Laura Gabbi, L.; Gerbaldo, D.; Gallo, F.; Nigro, N.; Icardi, G.; Viscoli, C.; Di Biagio, A. Vaginal HIV-1 shedding among HIV-1 infected women in the current era of combined antiretroviral therapy: A cross sectional study. Virulence 2017, 8, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Graham, S.M.; Holte, S.E.; Peshu, N.M.; Richardson, B.A.; Panteleeff, D.D.; Jaoko, W.G.; Ndinya-Achola, J.O.; Mandaliya, K.N.; Overbaugh, J.M.; McClelland, R.S. Initiation of antiretroviral therapy leads to a rapid decline in cervical and vaginal HIV-1 shedding. AIDS 2007, 21, 501–507. [Google Scholar] [CrossRef]
- Launay, O.; Tod, M.; Tschöpe, I.; Si-Mohamed, A.; Bélarbi, L.; Charpentier, C.; Goujard, C.; Taburet, A.M.; Lortholary, O.; Leroy, V.; et al. Residual HIV-1 RNA and HIV-1 DNA production in the genital tract reservoir of women treated with HAART: The prospective ANRS EP24 GYNODYN study. Antivir. Ther. 2011, 16, 843–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wessels, J.M.; Lajoie, J.; Vitali, D.; Omollo, K.; Kimani, J.; Oyugi, J.; Cheruiyot, J.; Kimani, M.; Mungai, J.N.; Akolo, M.; et al. Association of high-risk sexual behaviour with diversity of the vaginal microbiota and abundance of Lactobacillus. PLoS ONE 2017, 12, e0187612. [Google Scholar] [CrossRef] [Green Version]
- Borgdorff, H.; Tsivtsivadze, E.; Verhelst, R.; Marzorati, M.; Jurriaans, S.; Ndayisaba, G.F.; Schuren, F.H.; van de Wijgert, J.H. Lactobacillus-dominated cervicovaginal microbiota associated with reduced HIV/STI prevalence and genital HIV viral load in African women. ISME J. 2014, 8, 1781–1793. [Google Scholar] [CrossRef] [PubMed]
- Gosmann, C.; Anahtar, M.N.; Handley, S.A.; Farcasanu, M.; Abu-Ali, G.; Bowman, B.A.; Padavattan, N.; Desai, C.; Droit, L.; Moodley, A.; et al. Lactobacillus-Deficient Cervicovaginal Bacterial Communities Are Associated with Increased HIV Acquisition in Young South African Women. Immunity 2017, 46, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Tyssen, D.; Wang, Y.Y.; Hayward, J.A.; Agius, P.A.; DeLong, K.; Aldunate, M.; Ravel, J.; Moench, T.R.; Cone, R.A.; Tachedjian, G. Anti-HIV-1 Activity of Lactic Acid in Human Cervicovaginal Fluid. mSphere 2018, 3, e00055-18. [Google Scholar] [CrossRef] [Green Version]
- Rodger, A.J.; Cambiano, V.; Bruun, T.; Vernazza, P.; Collins, S.; Degen, O.; Corbelli, G.M.; Estrada, V.; Geretti, A.M.; Beloukas, A.; et al. Risk of HIV transmission through condomless sex in serodifferent gay couples with the HIV-positive partner taking suppressive antiretroviral therapy (PARTNER): Final results of a multicentre, prospective, observational study. Lancet 2019, 393, 2428–2438. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| ID | Age (Years) | Clinical Stage (CDC) | CD4+ Cells/ mm3 | Genotype | VL at Entry (Copies/mL) | Resistance-Associated Mutations Polymorphic | Drug Resistance (Stanford HIV db Program Rules) | Homology of Sequence between Blood and CVF Strains (%) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Blood | CVF | Blood | CVF | Blood | CVF | ||||||
| 001 | 43 | I | 988 | CRF02_AG | 87,200 | 1250 | None K20I M36I H69K L89M | None K20I M36I H69K L89M | None None | None None | 92.2 |
| 002 | 37 | III | 9 | CRF02_AG | 49,200 | 380 | None K20I M36I H69K L89M | L74V K20I M36I H69K L89M | None | ABC (intermediate) | 93.6 |
| 003 | 52 | III | 103 | CRF26_AU | 163,000 | 34,000 | None L10I/V M36I H69Q L89M | None L10I M36I H69Q L89M | None | None | 99.2 |
| 010 | 25 | I | 844 | CRF02_AG | 162,000 | 166 | None K20I M36I H69K L89M | None K20I M36I H69K | None | None | 96.9 |
| 011 | 29 | I | 1405 | B | 50,000 | 50 | None None | None None | None | None | 97.6 |
| 014 | 35 | III | 158 | CRF02_AG | 225,000 | 120,000 | None K20I M36I H69K L89M | None K20I M36I H69K L89M | None | None | 100 |
| 019 | 46 | III | 53 | CRF02_AG | 1,620,000 | 14,200 | E138A L10V K20M/V M36I H69K L89M | E138A L10V K20M/V M36I H69K L89M | RPV (low level) ETR (potential low level) | RPV (low level) ETR (potential low level) | 99.7 |
| 025 | 34 | III | 293 | CRF02_AG | 325,000 | 142,000 | None K20I M36I H69K L89M | None K20I M36I H69K L89M | None | None | 100 |
| 026 | 35 | II | 427 | CRF02_AG | 1,200,000 | 3900 | None K20I M36I H69K L89M | None K20I M36I H69K L89M | None | None | 98.8 |
| 030 | 28 | III | 68 | CRF02_AG | 468,000 | 67,500 | None L10V K20I M36I H69K L89M | None L10V M36I H69K L89M | None | None | 98.8 |
| 041 | 49 | II | 357 | CRF02_AG | 10,700 | 13,700 | None K20I M36I H69K L89M | None K20I M36I H69K L89M | None | None | 97.5 |
| 044 | 40 | III | 209 | CRF02_AG | 140,000 | 250 | None K20I M36I H69K L89M V179I | D67N K20I M36I H69K L89M V179I | None | ZDV (low level) | 98.2 |
| 046 | 33 | III | 113 | CRF02_AG | 300,200 | 5000 | K103N M36I H69K L89M | None M36I H69K L89M | None | None | 90.9 |
| 047 | 40 | III | 165 | CRF02_AG | 28,500 | 67,500 | None L10V M36I H69K L89M | D67N L10V M36I H69K L89M | None | ZDV (low level) | 98.1 |
| 049 | 29 | I | 436 | CRF02_AG | 33,000 | 282 | None K20I M36I H69K L89M | None K20I M36I H69K L89M | None | None | 91.3 |
| 055 | 46 | II | 412 | CRF02_AG | 142,000 | 37,700 | None L10V K20I M36I H69K L89M | None L10V K20I M36I H69K L89M | None | None | 99.1 |
| 070 | 26 | IV | 180 | CRF02_AG | 8804 | 16,500 | None K20I M36I H69K L89M | None K20I M36I H69K L89M | None | None | 99.3 |
| 080 | 47 | I | 536 | A1 | 100,000 | 26,400 | None K20I M36I H69K L89M | None K20I M36I H69K L89M | None | None | 98.2 |
| 081 | 34 | IV | 110 | CRF02_AG | 4300 | 27,400 | None L10V K20I M36I H69K L89M V179I | None K20I M36I H69K L89M | None | None | 93.3 |
| 085 | 32 | II | 592 | CRF02_AG | 29,500 | 575,000 | V179E K20I M36I H69K L89M | None M36I H69K L89M | EFV (potential low level) NVP (potential low level) ETR (potential low level) RPV (potential low level) | None | 93.4 |
| 091 | 60 | II | 183 | CRF06_cpx | 7950 | 41,400 | V106I K20I M36I H69K L89M | V106I K20I M36I H69K L89M | DOR (low level) NVP (potential low level) ETR (potential low level) RPV (potential low level) | DOR (low level) NVP (potential low level) ETR (potential low level) RPV (potential low level) | 96.3 |
| 096 | 33 | II | 225 | CRF06_cpx | 758,000 | 23,440 | None L10V K20I M36I H69K L89M | V179E L10V K20I M36I H69K L89M | None | EFV (potential low level) NVP (potential low level) ETR (potential low level) RPV (potential low level) | 95.2 |
| 101 | 25 | III | 13 | CRF02_AG | 241,000 | 13,200 | None L10V K20I M36I H69K L89M | None L10V K20I M36I H69K L89M | None | None | 100 |
| 107 | 18 | II | 787 | CRF02_AG | 19,500 | 8300 | None K20I M36I H69K L89M | None K20I M36I H69K L89M | None | None | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keita, A.; Rigaill, J.; Pillet, S.; Sereme, Y.; Coulibaly, S.; Diallo, F.; Verhoeven, P.; Pozzetto, B.; Thiero, T.A.; Bourlet, T. Evidence of HIV-1 Genital Shedding after One Year of Antiretroviral Therapy in Females Recently Diagnosed in Bamako, Mali. Microorganisms 2021, 9, 2164. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102164
Keita A, Rigaill J, Pillet S, Sereme Y, Coulibaly S, Diallo F, Verhoeven P, Pozzetto B, Thiero TA, Bourlet T. Evidence of HIV-1 Genital Shedding after One Year of Antiretroviral Therapy in Females Recently Diagnosed in Bamako, Mali. Microorganisms. 2021; 9(10):2164. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102164
Chicago/Turabian StyleKeita, Abdelaye, Josselin Rigaill, Sylvie Pillet, Youssouf Sereme, Souleymane Coulibaly, Fodé Diallo, Paul Verhoeven, Bruno Pozzetto, Tenin Aoua Thiero, and Thomas Bourlet. 2021. "Evidence of HIV-1 Genital Shedding after One Year of Antiretroviral Therapy in Females Recently Diagnosed in Bamako, Mali" Microorganisms 9, no. 10: 2164. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102164