
Immobilization of Polyethyleneimine (PEI) on Flat Surfaces and Nanoparticles Affects Its Ability to Disrupt Bacterial Membranes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental
3. Results and Discussion
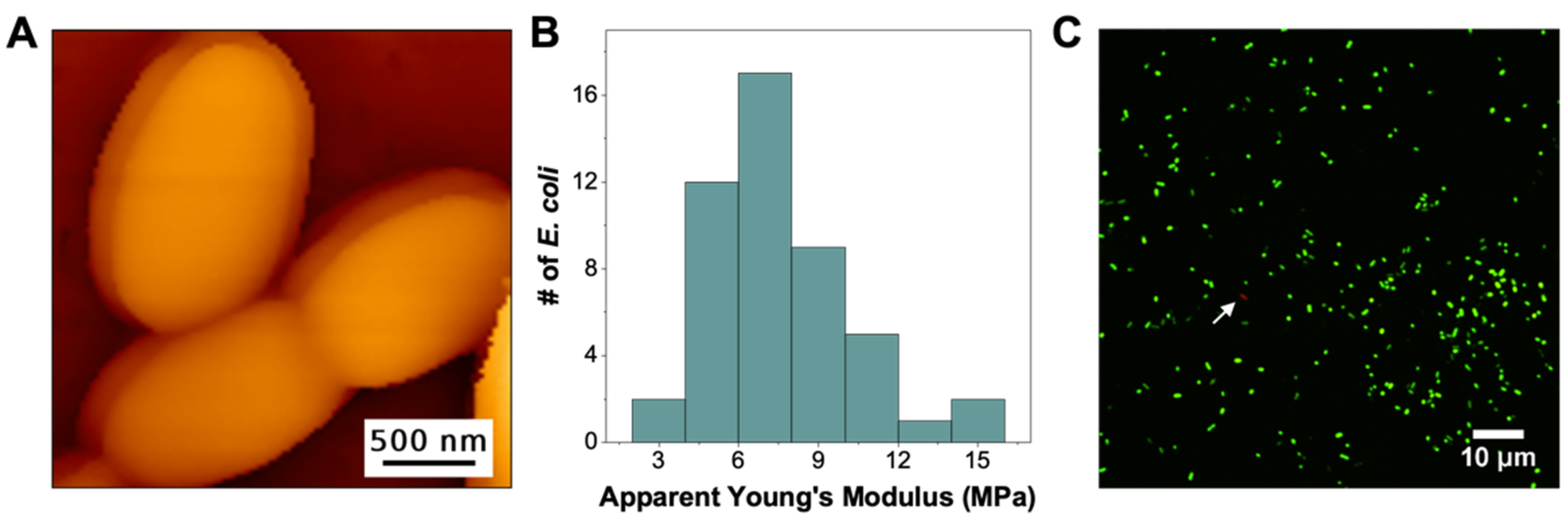
3.1. PEI Immobilization Affects Its Ability to Change the Structure and Mechanical Stability of E. coli
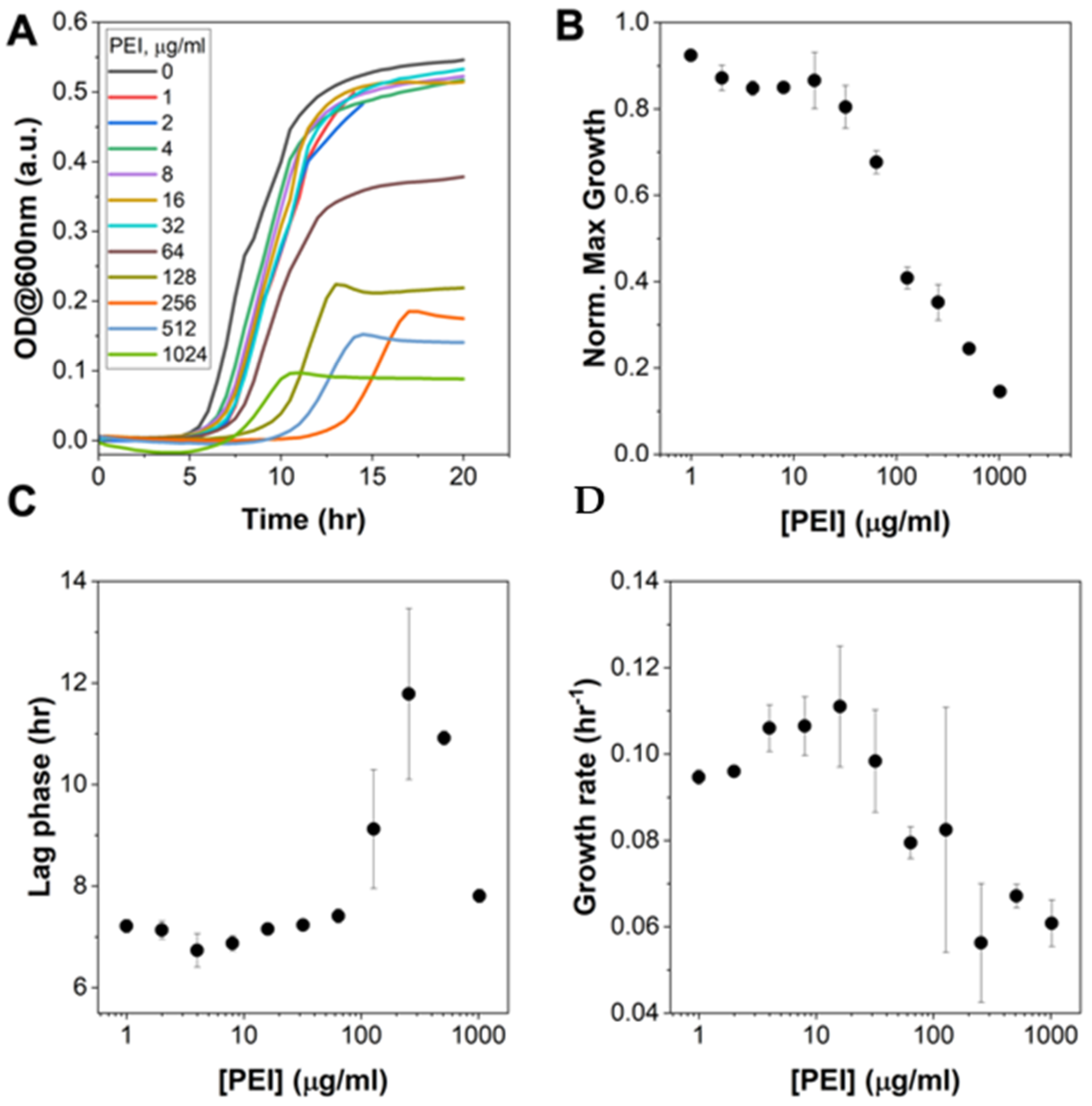
3.2. Pre-Treatment of E. coli with hMW-bPEI Does Affect Bacterial Growth, but Is Not Capable of Completely Killing the Bacteria Even at Very High Concentrations
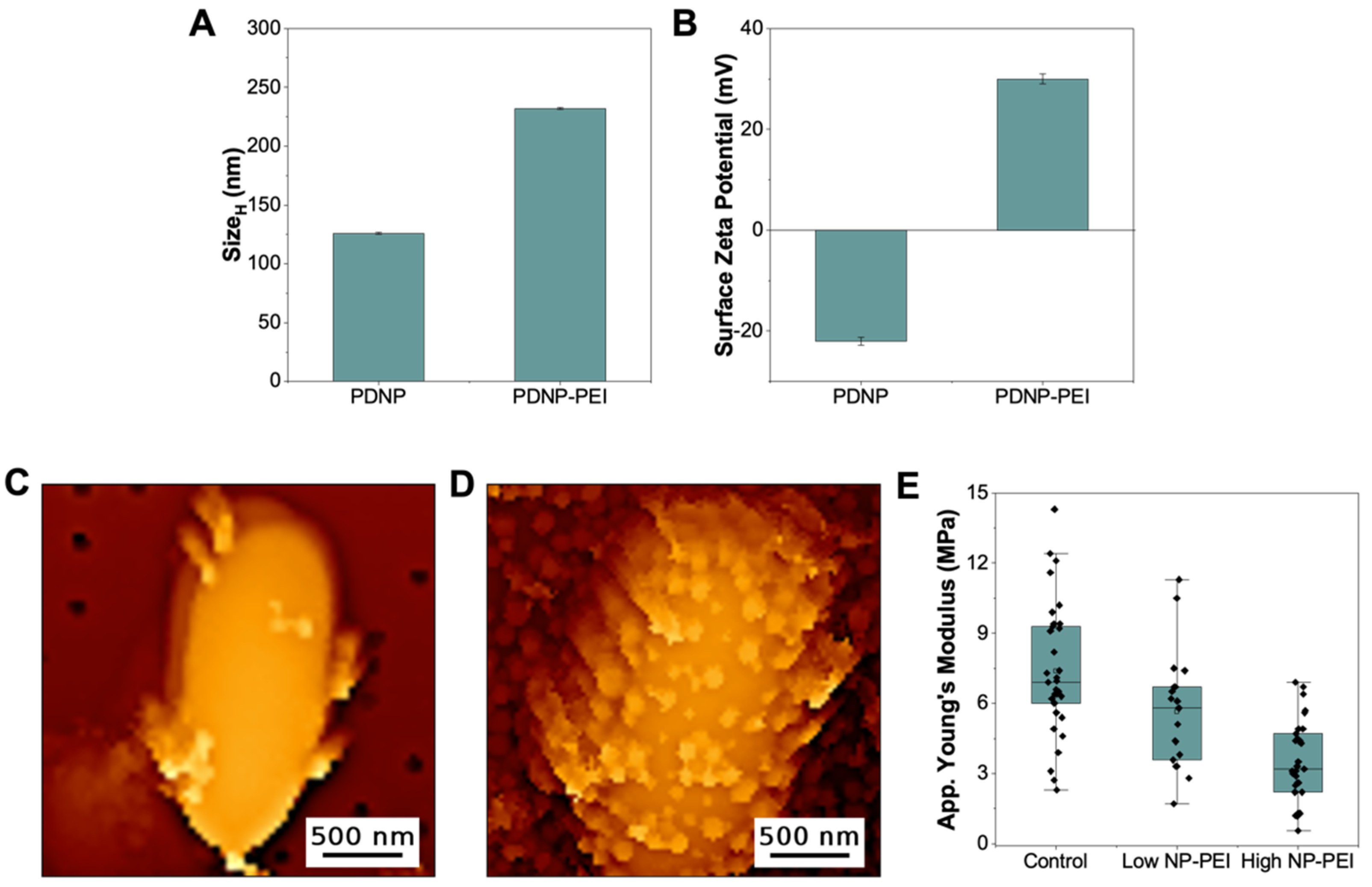
3.3. PEI-Decorated Polydopamine Nanoparticles Can Efficiently Target E. coli and Alter Cellular Mechanics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fischer, D.; Li, Y.; Ahlemeyer, B.; Krieglstein, J.; Kissel, T. In Vitro Cytotoxicity Testing of Polycations: Influence of Polymer Structure on Cell Viability and Hemolysis. Biomaterials 2003, 24, 1121–1131. [Google Scholar] [CrossRef]
- Hong, S.; Leroueil, P.R.; Janus, E.K.; Peters, J.L.; Kober, M.M.; Islam, M.T.; Orr, B.G.; Baker, J.R., Jr.; Holl, M.M.B. Interaction of Polycationic Polymers with Supported Lipid Bilayers and Cells: Nanoscale Hole Formation and Enhanced Membrane Permeability. Bioconjug. Chem. 2006, 17, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Purohit, G.; Sakthivel, T.; Florence, A.T. Interaction of Cationic Partial Dendrimers with Charged and Neutral Liposomes. Int. J. Pharm. 2001, 214, 71–76. [Google Scholar] [CrossRef]
- Yaroslavov, A.A.; Kuchenkova, O.Y.; Okuneva, I.B.; Melik-Nubarov, N.S.; Kozlova, N.O.; Lobyshev, V.I.; Menger, F.M.; Kabanov, V.A. Effect of Polylysine on Transformations and Permeability of Negative Vesicular Membranes. Biochim. Biophys. Acta 2003, 1611, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Mintzer, M.A.; Simanek, E.E. Nonviral Vectors for Gene Delivery. Chem. Rev. 2009, 109, 259–302. [Google Scholar] [CrossRef]
- Xia, T.; Kovochich, M.; Liong, M.; Meng, H.; Kabehie, S.; George, S.; Zink, J.I.; Nel, A.E. Polyethyleneimine Coating Enhances the Cellular Uptake of Mesoporous Silica Nanoparticles and Allows Safe Delivery of Sirna and DNA Constructs. ACS Nano 2009, 3, 3273–3286. [Google Scholar] [CrossRef]
- Lungwitz, U.; Breunig, M.; Blunk, T.; Gopferich, A. Polyethylenimine-Based Non-Viral Gene Delivery Systems. Eur. J. Pharm. Biopharm. 2005, 60, 247–266. [Google Scholar] [CrossRef]
- Dick, C.R.; Ham, G.E. Characterization of Polyethylenimine. J. Macromol. Sci. Part A Chem. 1970, 4, 1301–1314. [Google Scholar] [CrossRef]
- Wang, Y.K.; Krasnopeeva, E.; Lin, S.Y.; Bai, F.; Pilizota, T.; Lo, C.J. Comparison of Escherichia Coli Surface Attachment Methods for Single-Cell Microscopy. Sci. Rep. 2019, 9, 19418. [Google Scholar] [CrossRef] [Green Version]
- Gibney, K.A.; Sovadinova, I.; Lopez, A.I.; Urban, M.; Ridgway, Z.; Caputo, G.A.; Kuroda, K. Poly(Ethylene Imine)S as Antimicrobial Agents with Selective Activity. Macromol. Biosci. 2012, 12, 1279–1289. [Google Scholar] [CrossRef] [Green Version]
- Helander, I.M.; Latva-Kala, K.; Lounatmaa, K. Permeabilizing Action of Polyethyleneimine on Salmonella Typhimurium Involves Disruption of the Outer Membrane and Interactions with Lipopolysaccharide. Microbiology 1998, 144 Pt 2, 385–390. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Qiu, S.; Lewis, K.; Klibanov, A.M. Bactericidal Properties of Flat Surfaces and Nanoparticles Derivatized with Alkylated Polyethylenimines. Biotechnol. Prog. 2002, 18, 1082–1086. [Google Scholar] [CrossRef] [PubMed]
- Domb, A.J.; Yudovin-Farber, I.; Golenser, J.; Beyth, N.; Weiss, E.I. Quaternary Ammonium Polyethyleneimine: Antibacterial Activity. J. Nanomater. 2010, 2010, 826343. [Google Scholar]
- Lin, J.; Qiu, S.; Lewis, K.; Klibanov, A.M. Mechanism of Bactericidal and Fungicidal Activities of Textiles Covalently Modified with Alkylated Polyethylenimine. Biotechnol. Bioeng. 2003, 83, 168–172. [Google Scholar] [CrossRef]
- Liu, H.; Elkin, I.; Chen, J.; Klibanov, A.M. Why Do Some Immobilized N-Alkylated Polyethylenimines Far Surpass Others in Inactivating Influenza Viruses? Biomacromolecules 2015, 16, 351–356. [Google Scholar] [CrossRef]
- Dague, E.; Le, D.T.; Zanna, S.; Marcus, P.; Loubiere, P.; Mercier-Bonin, M. Probing In Vitro Interactions between Lactococcus Lactis and Mucins Using Afm. Langmuir 2010, 26, 11010–11017. [Google Scholar] [CrossRef]
- Jauvert, E.; Palleau, E.; Dague, E.; Ressier, L. Directed Assembly of Living Pseudomonas Aeruginosa Bacteria on Pei Patterns Generated by Nanoxerography for Statistical Afm Bioexperiments. ACS Appl. Mater. Interfaces 2014, 23, 21230–21236. [Google Scholar] [CrossRef]
- Zhavnerko, G.; Poleschuyk, N.N. Mycobacterium under Afm Tip: Advantages of Polyelectrolyte Modified Substrate. Int. J. Mycobacteriol. 2012, 1, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Chopinet, L.; Formosa, C.; Rols, M.P.; Duval, R.E.; Dague, E. Imaging Living Cells Surface and Quantifying Its Properties at High Resolution Using Afm in Qi Mode. Micron 2013, 48, 26–33. [Google Scholar] [CrossRef]
- Mulansky, S.; Saballus, M.; Friedrichs, J.; Bley, T.; Boschke, E. A Novel Protocol to Prepare Cell Probes for the Quantification of Microbial Adhesion and Biofilm Initiation on Structured Bioinspired Surfaces Using Afm for Single-Cell Force Spectroscopy: Dedicated to Prof. Em. Dr. Dr. H.C. Karl Schugerl on the Occasion of His 90th Birthday. Eng. Life Sci. 2017, 17, 833–840. [Google Scholar] [PubMed] [Green Version]
- Helander, I.M.; Alakomi, H.L.; Latva-Kala, K.; Koski, P. Polyethyleneimine Is an Effective Permeabilizer of Gram-Negative Bacteria. Microbiology 1997, 143 Pt 10, 3193–3199. [Google Scholar] [CrossRef] [Green Version]
- Foxley, M.A.; Wright, S.N.; Lam, A.K.; Friedline, A.W.; Strange, S.J.; Xiao, M.T.; Moen, E.L.; Rice, C.V. Targeting Wall Teichoic Acid in Situ with Branched Polyethylenimine Potentiates Beta-Lactam Efficacy against Mrsa. ACS Med. Chem. Lett. 2017, 8, 1083–1088. [Google Scholar] [CrossRef]
- Khalil, H.; Chen, T.; Riffon, R.; Wang, R.; Wang, Z. Synergy between Polyethylenimine and Different Families of Antibiotics against a Resistant Clinical Isolate of Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 2008, 5, 1635–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutter, J.L.; Bechhoefer, J. Calibration of Atomic-Force Microscope Tips. Rev. Sci. Instrum. 1993, 64, 1868–1873. [Google Scholar] [CrossRef] [Green Version]
- Andoy, N.M.O.; Jeon, K.; Kreis, C.T.; Sullan, R.M.A. Multifunctional and Stimuli-Responsive Polydopamine Nanoparticle-Based Platform for Targeted Antimicrobial Applications. Adv. Funct. Mater. 2020, 30, 2004503. [Google Scholar] [CrossRef]
- Kreis, C.T.; Sullan, R.M.A. Interfacial Nanomechanical Heterogeneity of the E. Coli Biofilm Matrix. Nanoscale 2020, 12, 16819–16830. [Google Scholar] [CrossRef] [PubMed]
- Kasas, S.; Ikai, A. A Method for Anchoring Round Shaped Cells for Atomic Force Microscope Imaging. Biophys. J. 1995, 68, 1678–1680. [Google Scholar] [CrossRef] [Green Version]
- Tuson, H.H.; Auer, G.K.; Renner, L.D.; Hasebe, M.; Tropini, C.; Salick, M.; Crone, W.C.; Gopinathan, A.; Huang, K.C.; Weibel, D.B. Measuring the Stiffness of Bacterial Cells from Growth Rates in Hydrogels of Tunable Elasticity. Mol. Microbiol. 2012, 84, 874–891. [Google Scholar] [CrossRef] [Green Version]
- Bagheri, M.; Beyermann, M.; Dathe, M. Immobilization Reduces the Activity of Surface-Bound Cationic Antimicrobial Peptides with No Influence Upon the Activity Spectrum. Antimicrob. Agents Chemother. 2009, 53, 1132–1141. [Google Scholar] [CrossRef] [Green Version]
- Alakomi, H.L.; Paananen, A.; Suihko, M.L.; Helander, I.M.; Saarela, M. Weakening Effect of Cell Permeabilizers on Gram-Negative Bacteria Causing Biodeterioration. Appl. Environ. Microbiol. 2006, 72, 4695–4703. [Google Scholar] [CrossRef] [Green Version]
- Krapf, M.E.; Lartiges, B.; Merlin, C.; Francius, G.; Ghanbaja, J.; Duval, J.F. Deciphering the Aggregation Mechanism of Bacteria (Shewanella Oneidensis Mr1) in the Presence of Polyethyleneimine: Effects of the Exopolymeric Superstructure and Polymer Molecular Weight. Coll. Surf. B Biointerfaces 2016, 139, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Krapf, M.E.; Lartiges, B.S.; Merlin, C.; Francius, G.; Ghanbaja, J.; Duval, J.F. Polyethyleneimine-Mediated Flocculation of Shewanella Oneidensis Mr-1: Impacts of Cell Surface Appendage and Polymer Concentration. Water Res. 2012, 46, 1838–1846. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.H.; Messersmith, P.B.; Lee, H. Polydopamine Surface Chemistry: A Decade of Discovery. ACS Appl. Mater. Interfaces 2018, 10, 7523–7540. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andoy, N.M.O.; Patel, M.; Lui, C.L.J.; Sullan, R.M.A. Immobilization of Polyethyleneimine (PEI) on Flat Surfaces and Nanoparticles Affects Its Ability to Disrupt Bacterial Membranes. Microorganisms 2021, 9, 2176. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102176
Andoy NMO, Patel M, Lui CLJ, Sullan RMA. Immobilization of Polyethyleneimine (PEI) on Flat Surfaces and Nanoparticles Affects Its Ability to Disrupt Bacterial Membranes. Microorganisms. 2021; 9(10):2176. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102176
Chicago/Turabian StyleAndoy, Nesha May Octavio, Meera Patel, Ching Lam Jane Lui, and Ruby May Arana Sullan. 2021. "Immobilization of Polyethyleneimine (PEI) on Flat Surfaces and Nanoparticles Affects Its Ability to Disrupt Bacterial Membranes" Microorganisms 9, no. 10: 2176. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102176