
Genotype-Dependent Immunogenicity of Dengue Virus Type 2 Asian I and Asian/American Genotypes in Common Marmoset (Callithrix jacchus): Discrepancy in Neutralizing and Infection-Enhancing Antibody Levels between Genotypes

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. DENV Infection in Marmoset Models
2.3. Determination of Anti-DENV Immunoglobulin M (IgM) and IgG Antibody
2.4. Determination of Neutralizing Antibody Titers Using a 50% Plaque Reduction Neutralization Test Assay (PRNT50)
2.5. Determination of Infection-Enhancing Activity Using the ADE Assay with BHK-21 and FcγR-Expressing BHK Cells
2.6. Viral RNA Extraction
2.7. Quantification of Viral RNA Copy Number
2.8. Determination of DENV Titer Using Plaque Assay
2.9. Determination of Dengue NS1 Antigen
2.10. Data Analysis
2.11. Ethical Approval
3. Results
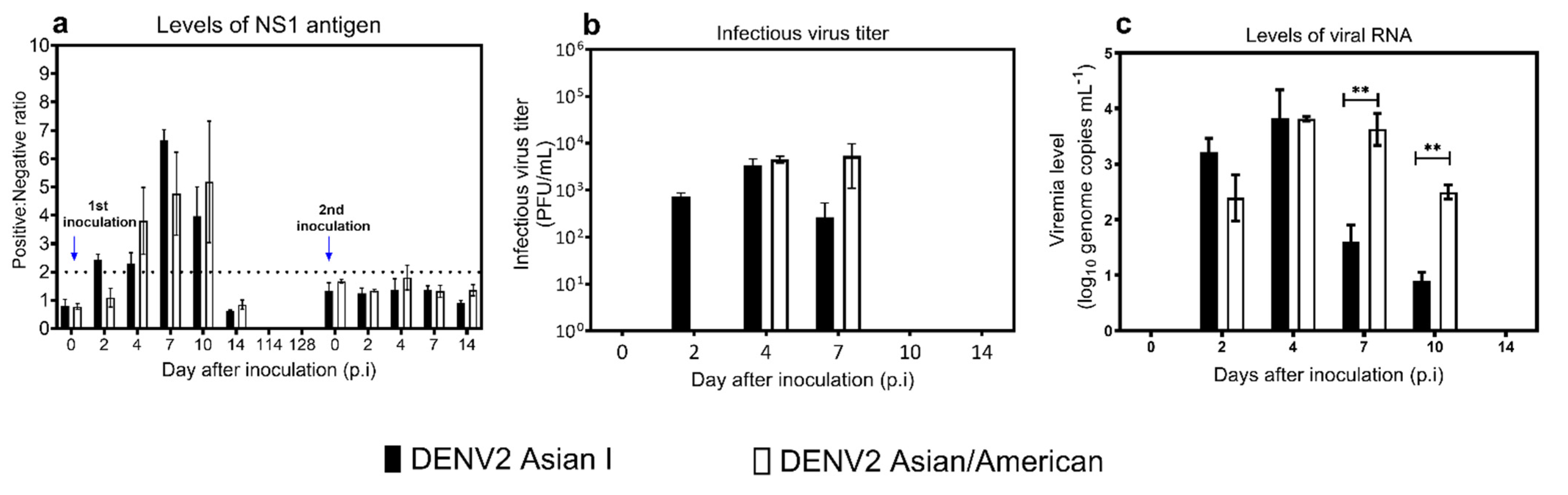
3.1. Viremia Levels
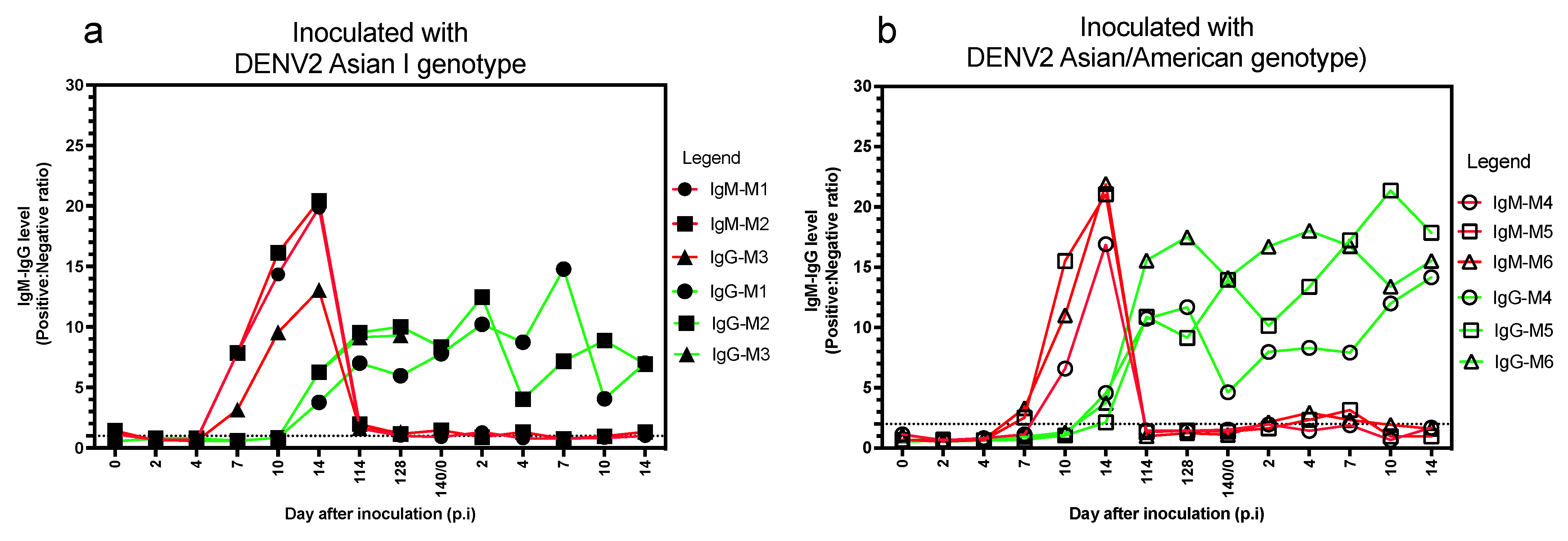
3.2. Anti-DENV IgM and IgG Antibody Levels
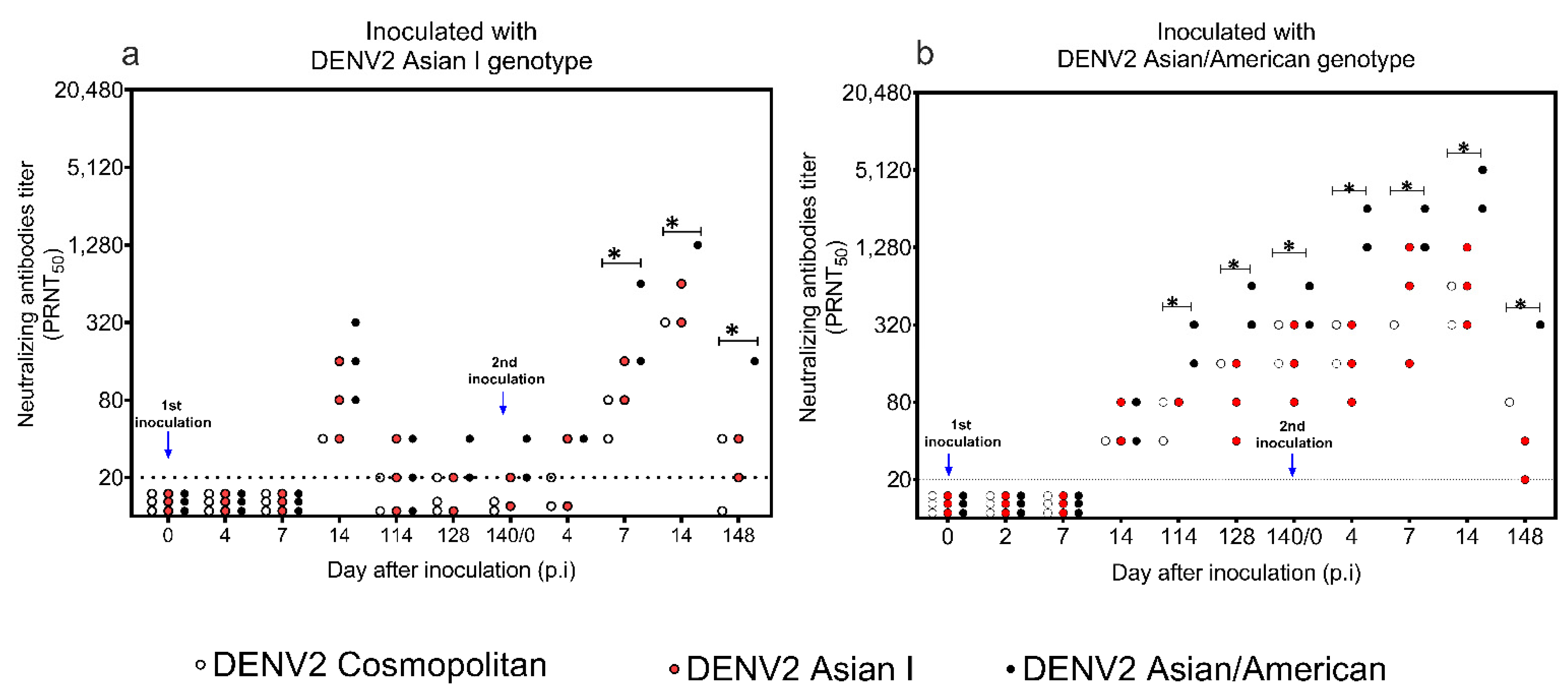
3.3. Levels of Neutralizing Antibody to DENV2 Asian I, Cosmopolitan, and AA Genotypes
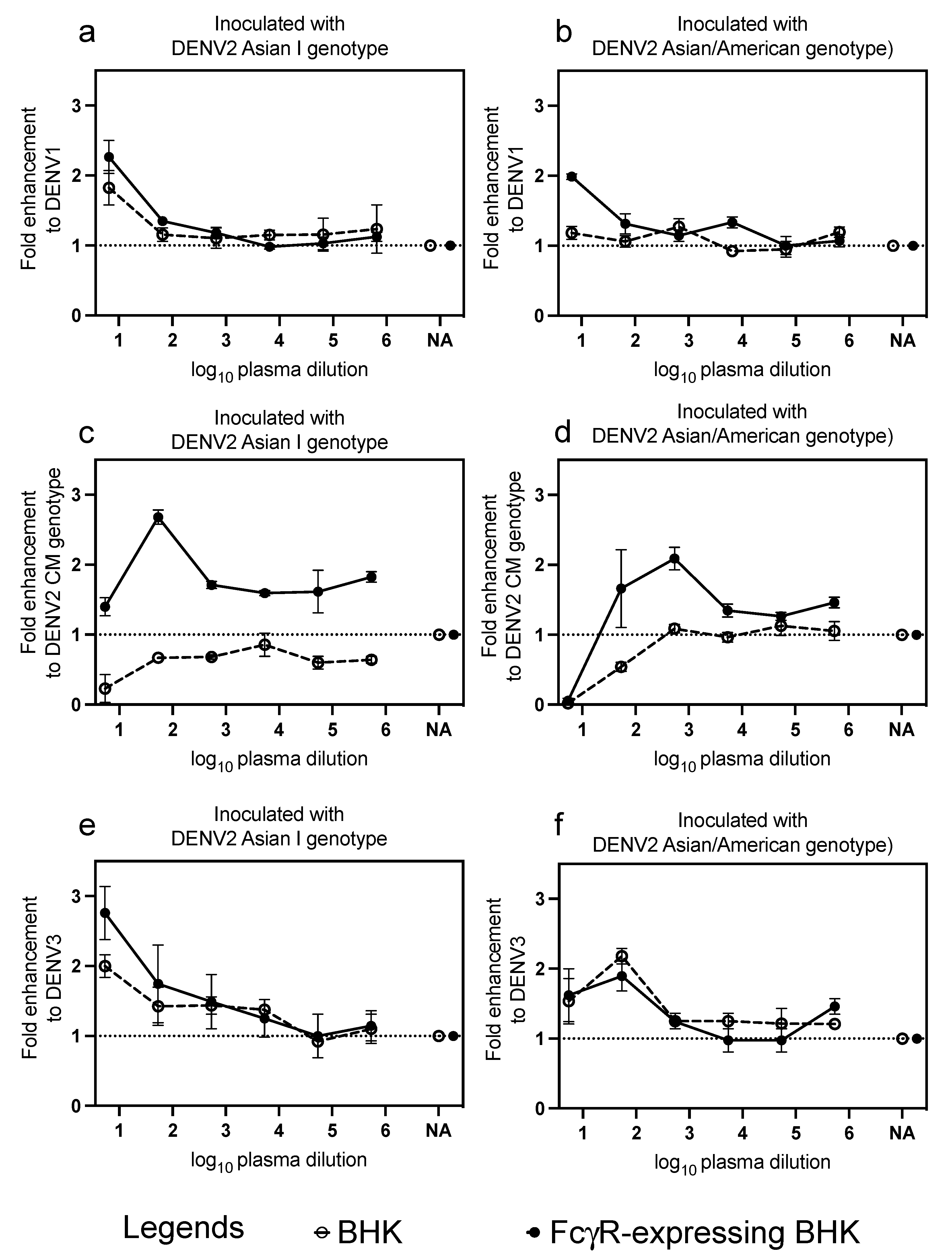
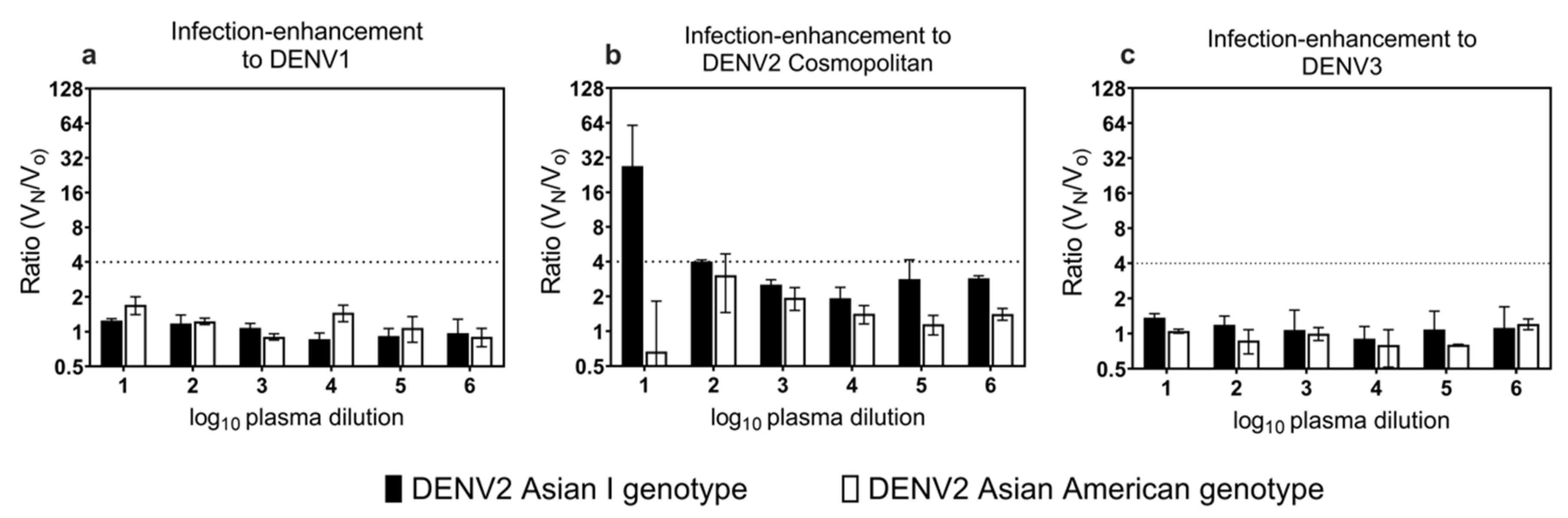
3.4. Cross-Reactive Neutralizing Antibodies and Infection-Enhancing Activity to Heterologous Genotype and Serotype
3.5. Cross-Reactive Neutralizing Antibodies to ZIKV and JEV
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rothman, A.L. Immunity to dengue virus: A tale of original antigenic sin and tropical cytokine storms. Nat. Rev. Immunol. 2011, 11, 532–543. [Google Scholar] [CrossRef] [PubMed]
- Cedillo-Barrón, L.; García-Cordero, J.; Bustos-Arriaga, J.; León-Juárez, M.; Gutiérrez-Castañeda, B. Antibody response to dengue virus. Microbes Infect. 2014, 16, 711–720. [Google Scholar] [CrossRef]
- Drumond, B.P.; Mondini, A.; Schmidt, D.J.; Bronzoni, R.V.; Bosch, I.; Nogueira, M.L. Circulation of different lineages of dengue virus 2, genotype American/Asian in Brazil: Dynamics and molecular and phylogenetic characterization. PLoS ONE 2013, 8, e59422. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ge, P.; Yu, X.; Brannan, J.M.; Bi, G.; Zhang, Q.; Schein, S.; Zhou, Z.H. Cyro-EM structure of the mature dengue virus at 3.5-A resolution. Nat. Struct. Mol. Biol. 2013, 20, 105–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathew, A.; Rothman, A.L. Understanding the contribution of cellular immunity to dengue disease pathogenesis. Immunol. Rev. 2008, 225, 300–313. [Google Scholar] [CrossRef] [PubMed]
- Forshey, B.M.; Reiner, R.C.; Olkowski, S.; Morrison, A.C.; Espinoza, A.; Long, K.C.; Vilcarromero, S.; Casanova, W.; Wearing, H.J.; Halsey, E.S.; et al. Incomplete protection against dengue virus type 2 re-infection in Peru. PLoS Negl. Trop. Dis. 2016, 10, e0004398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinz, F.X.; Stiasny, K. Flaviviruses and their antigen structure. J. Clin. Virol. 2012, 55, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Hadinegoro, S.R.S. The revised WHO dengue case classification: Does the system need to be modified? Paediatr. Int. Child Health 2012, 32, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman, M.G.; Harris, E. Dengue. Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef]
- Guzman, M.G.; Halstead, S.B.; Artsob, H.; Buchy, P.; Farrar, J.; Gubler, D.J.; Hunsperger, E.; Kroeger, A.; Margolis, H.S.; Martínez, E.; et al. Dengue: A continuing global threat. Nat. Rev. Microbiol. 2010, 8, S7–S16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dengue Guidelines, for Diagnosis, Treatment, Prevention and Control; World Health Organization: Geneva, Switzerland, 2009.
- Dengue Guidelines for Patient Care in the Region of the Americans, 2nd ed.; Pan American Health Organization: Washington, DC, USA; World Health Organization: Geneva, Switzerland, 2016.
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Zhou, Z.; Wen, Z.; Liu, Y.; Zeng, C.; Xiao, D.; Ou, M.; Han, Y.; Huang, S.; Liu, D.; et al. Global epidemiology of dengue outbreaks in 1990-2015: A systematic review and meta-analysis. Front. Cell. Infect. Microbiol. 2017, 7, 317. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Li, Y. Global evolutionary history and spatio-temporal dynamics of dengue virus type 2. Sci. Rep. 2017, 7, 45505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twiddy, S.S.; Farrar, J.J.; Vinh Chau, N.; Wills, B.; Gould, E.A.; Gritsun, T.; Lloyd, G.; Holmes, E.C. Phylogenetic relationships and differential selection pressures among genotypes of dengue-2 virus. Virology 2002, 298, 63–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waman, V.P.; Kolekar, P.; Ramtirthkar, M.R.; Kale, M.M.; Kulkarni-Kale, U. Analysis of genotype diversity and evolution of dengue virus serotype 2 using complete genomes. PeerJ 2016, 4, e2326. [Google Scholar] [CrossRef] [Green Version]
- Salazar, M.; Pino, M.A.L.; Ale, J.A.F.; Olson, K.E.; Beaty, B.J. American and American/Asian genotypes of dengue virus differ in mosquito infection efficiency: Candidate molecular determinants of productive vector infection. Rev. Biomédica 2010, 21, 121–135. [Google Scholar]
- Leitmeyer, K.C.; Vaughn, D.W.; Watts, D.M.; Salas, R.; Villalobos, I.; de Chacon; Ramos, C.; Rico-Hesse, R. Dengue virus structural differences that correlate with pathogenesis. J. Virol. 1999, 73, 4738–4747. [Google Scholar] [CrossRef] [Green Version]
- Vu, T.T.H.; Holmes, E.C.; Duong, V.; Nguyen, T.Q.; Tran, T.H.; Quail, M.; Churcher, C.; Parkhill, J.; Cardosa, J.; Farrar, J.; et al. Emergence of the Asian 1 genotype of dengue virus serotype 2 in Viet Nam: In vivo fitness advantage and lineage replacement in South-East Asia. PLoS Negl. Trop. Dis. 2010, 4, e757. [Google Scholar] [CrossRef]
- Wang, C.; Katzelnick, L.C.; Montoya, M.; Hue, K.D.T.; Simmons, C.P.; Harris, E. Evolutionarily successful Asian 1 dengue virus 2 lineages contain one substitution in envelope that increases sensitivity to polyclonal antibody neutralization. J. Infect. Dis. 2015, 213, 975–984. [Google Scholar] [CrossRef] [Green Version]
- Brien, J.D.; Austin, S.K.; Sukupolvi-Petty, S.; O’Brien, K.M.; Johnson, S.; Fremont, D.H.; Diamond, M.S. Genotype-specific neutralization and protection by antibodies against dengue virus type 3. J. Virol. 2010, 84, 10630–10643. [Google Scholar] [CrossRef] [Green Version]
- Zompi, S.; Harris, E. Original antigenic sin in dengue revisited. Proc. Natl. Acad. Sci. USA 2013, 110, 8761–8762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, G.; Si, L.; Wang, Y.; Zhou, J.; Yan, H.; Jiang, L. Antibody-dependent enhancement (ADE) of dengue virus: Identification of the key amino acid that is vital in DENV vaccine research. J. Gene Med. 2021, 23, e3297. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.B. Vaccine-associated enhanced viral disease: Implication for viral vaccine development. BioDrugs 2021, 35, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, M.; Stollenwerk, N.; Halstead, S.B. The risks behind Dengvaxia recommendation. Lancet Infect. Dis. 2016, 16, 882–883. [Google Scholar] [CrossRef]
- Hadinegoro, S.R.; Arredondo-García, J.L.; Capeding, M.R.; Deseda, C.; Chotpitayasunondh, T.; Dietze, R.; Hj Muhammad Ismail, H.I.; Reynales, H.; Limkittikul, K.; Rivera-Medina, D.M.; et al. Efficacy and long-term safety of a dengue vaccine in regions of endemic disease. N. Engl. J. Med. 2015, 373, 1195–1206. [Google Scholar] [CrossRef] [Green Version]
- Azami, N.A.M.; Moi, M.L.; Ami, Y.; Suzaki, Y.; Lim, C.-K.; Taniguchi, S.; Saijo, M.; Takasaki, T.; Kurane, I. Genotype-specific and cross-reactive neutralizing antibodies induced by dengue virus infection: Detection of antibodies with different levels of neutralizing activities against homologous and heterologous genotypes of dengue virus type 2 in common marmoset (Callithrix jacchus). Virol. J. 2018, 15, 51. [Google Scholar] [CrossRef]
- Omatsu, T.; Moi, M.L.; Hirayama, T.; Takasaki, T.; Nakamura, S.; Tajima, S.; Ito, M.; Yoshida, T.; Saito, A.; Katakai, Y.; et al. Common marmoset (Callithrix jacchus) as a primate model of dengue virus infection: Development of high levels of viraemia and demonstration of protective immunity. J. Gen. Virol. 2011, 92, 2272–2280. [Google Scholar] [CrossRef]
- Moi, M.L.; Ami, Y.; Azami, N.A.M.; Shirai, K.; Yoksan, S.; Suzaki, Y.; Kitaura, K.; Lim, C.-K.; Saijo, M.; Suzuki, R.; et al. Marmosets (Callithrix jacchus) as a non-human primate model for evaluation of candidate dengue vaccines: Induction and maintenance of specific protective immunity against challenges with clinical isolates. J. Gen. Virol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Yamada, K.I.; Takasaki, T.; Pandey, B.; Nerome, R.; Tajima, S.; Morita, K.; Kurane, I. Phylogenetics analysis of dengue viruses isolated from imported dengue patients: Possible aid for determining the countries where infections occurred. J. Travel Med. 2007, 14, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA 6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moi, M.L.; Lim, C.K.; Kotaki, A.; Takasaki, T.; Kurane, I. Development of an antibody-dependent enhancement assay for dengue virus using stable BHK-21 cell lines expressing Fc gamma RIIA. J. Virol. Methods 2010, 163, 205–209. [Google Scholar] [CrossRef]
- Moi, M.L.; Lim, C.K.; Kotaki, A.; Takasaki, T.; Kurane, I. Detection of higher levels of dengue viremia using FcγR-expressing BHK-21 cells than FcγR-negative cells in secondary infection but not in primary infection. J. Infect. Dis. 2011, 203, 1405–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, M.; Takasaki, T.; Yamada, K.; Nerome, R.; Tajima, S.; Kurane, I. Development and evaluation of fluorogenic TaqMan reverse transcriptase PCR assays for detection of dengue virus types 1 to 4. J. Clin. Microbiol. 2004, 42, 5935–5937. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marmoset ID | Levels of Neutralizing Antibody (PRNT50) | |||||
|---|---|---|---|---|---|---|
| DENV1 | DENV2 AI | DENV2 CM | DENV2 AA | DENV3 | DENV4 | |
| Inoculated with DENV2 Asian I | ||||||
| M1 | <5 | 5 | <5 | 20 | 5 | <5 |
| M2 | <5 | 20 | 5 | 40 | <5 | <5 |
| Inoculated with DENV2 Asian/American | ||||||
| M4 | <5 | 80 | 160 | 1280 | 5 | 10 |
| M5 | 5 | 160 | 320 | 640 | 10 | <5 |
| M6 | 5 | 320 | 320 | 640 | 10 | 10 |
| Marmoset ID | Levels of Neutralizing Antibodies (PRNT50) | |
|---|---|---|
| ZIKV | JEV | |
| Inoculated with DENV2 Asian I | ||
| M1 | <5 | <5 |
| M2 | <5 | <5 |
| Inoculated with DENV2 Asian/American | ||
| M4 | <5 | <5 |
| M5 | <5 | <5 |
| M6 | <5 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azami, N.A.M.; Moi, M.L.; Ami, Y.; Suzaki, Y.; Taniguchi, S.; Tajima, S.; Saijo, M.; Takasaki, T.; Kurane, I.; Lim, C.-K. Genotype-Dependent Immunogenicity of Dengue Virus Type 2 Asian I and Asian/American Genotypes in Common Marmoset (Callithrix jacchus): Discrepancy in Neutralizing and Infection-Enhancing Antibody Levels between Genotypes. Microorganisms 2021, 9, 2196. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112196
Azami NAM, Moi ML, Ami Y, Suzaki Y, Taniguchi S, Tajima S, Saijo M, Takasaki T, Kurane I, Lim C-K. Genotype-Dependent Immunogenicity of Dengue Virus Type 2 Asian I and Asian/American Genotypes in Common Marmoset (Callithrix jacchus): Discrepancy in Neutralizing and Infection-Enhancing Antibody Levels between Genotypes. Microorganisms. 2021; 9(11):2196. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112196
Chicago/Turabian StyleAzami, Nor Azila Muhammad, Meng Ling Moi, Yasushi Ami, Yuriko Suzaki, Satoshi Taniguchi, Shigeru Tajima, Masayuki Saijo, Tomohiko Takasaki, Ichiro Kurane, and Chang-Kweng Lim. 2021. "Genotype-Dependent Immunogenicity of Dengue Virus Type 2 Asian I and Asian/American Genotypes in Common Marmoset (Callithrix jacchus): Discrepancy in Neutralizing and Infection-Enhancing Antibody Levels between Genotypes" Microorganisms 9, no. 11: 2196. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112196