
CsrA Regulates Swarming Motility and Carbohydrate and Amino Acid Metabolism in Vibrio alginolyticus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, and Media
2.2. Mutant and Plasmid Construction
2.3. Motility
2.4. Growth Measurement
2.5. RNA Isolation and Quantitative Reverse Transcription PCR (qRT-PCR) Analysis
2.6. RNA Stability Measurement
3. Results
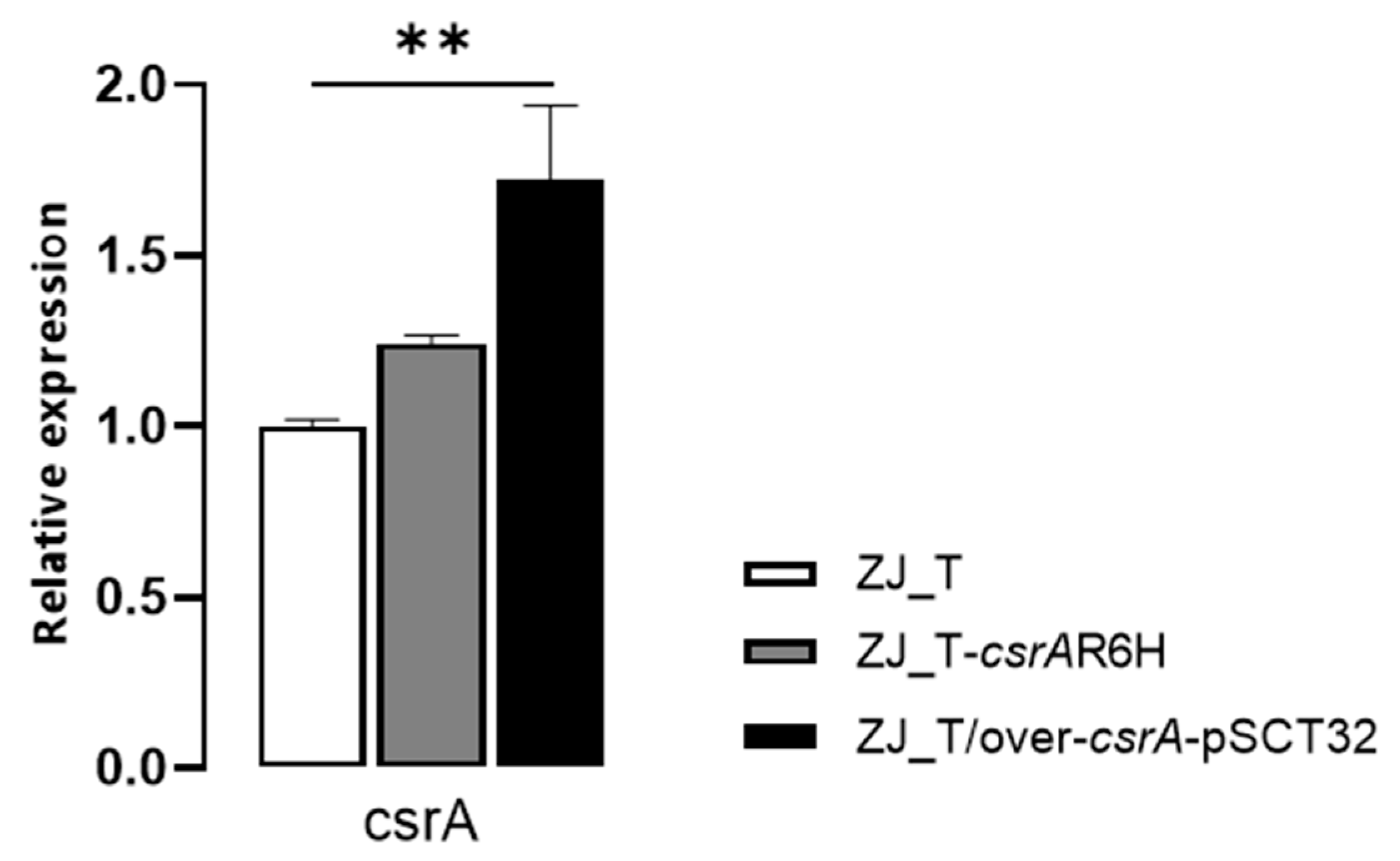
3.1. Construction of csrA Pointmutant and Overexpression Strain
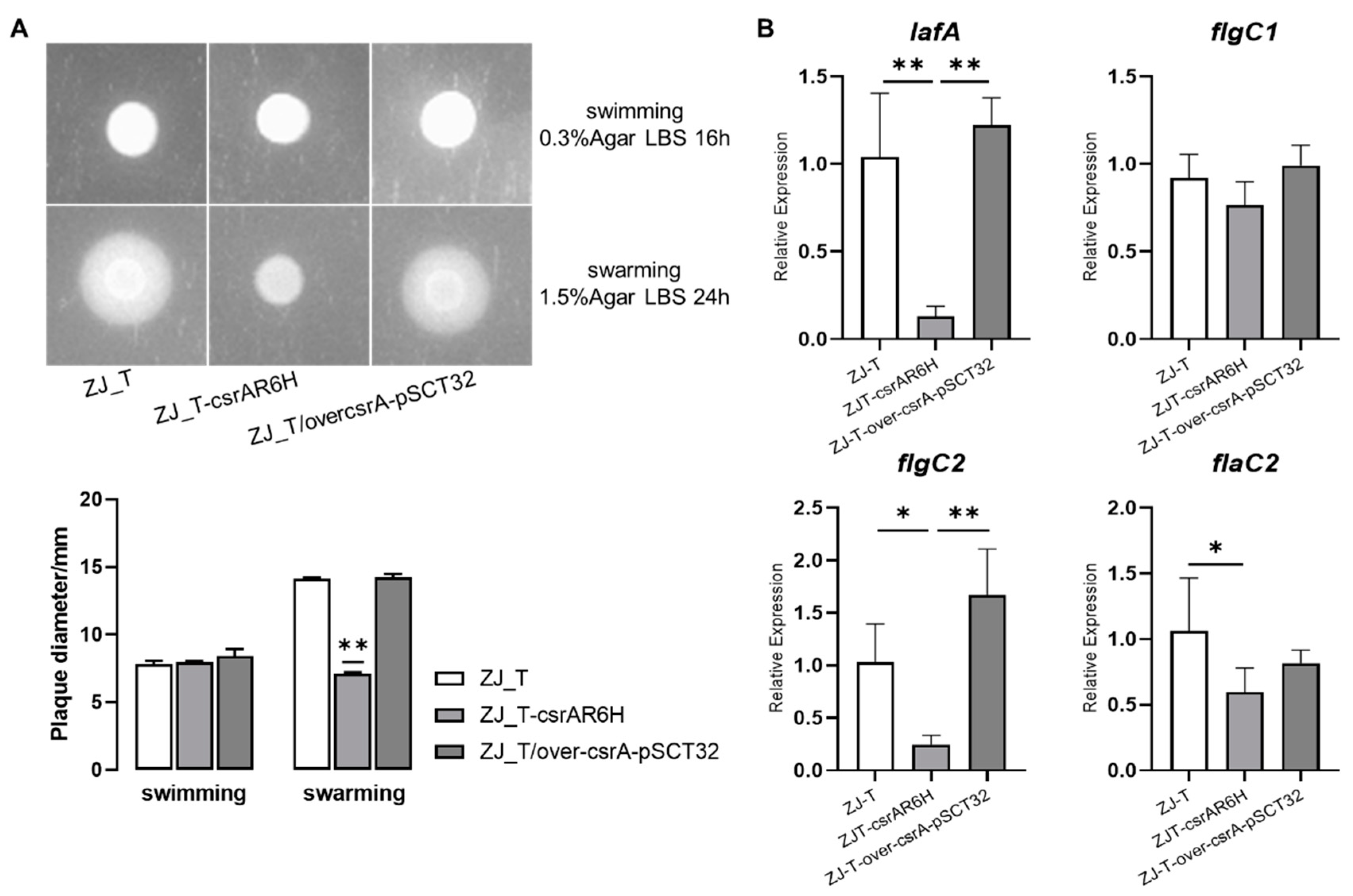
3.2. CsrA Positively Regulates Swarming but Not Swimming Motility in V. alginolyticus
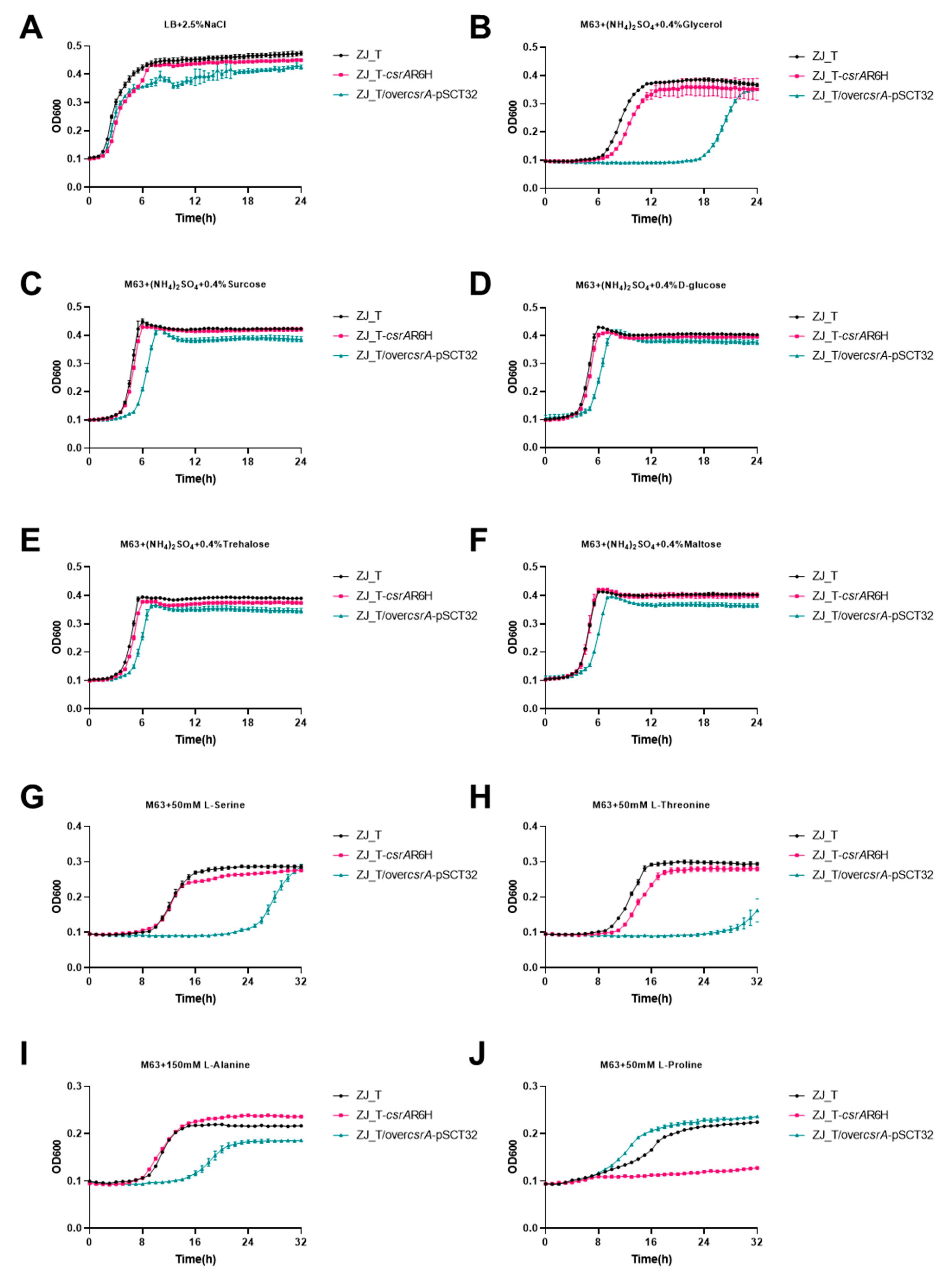
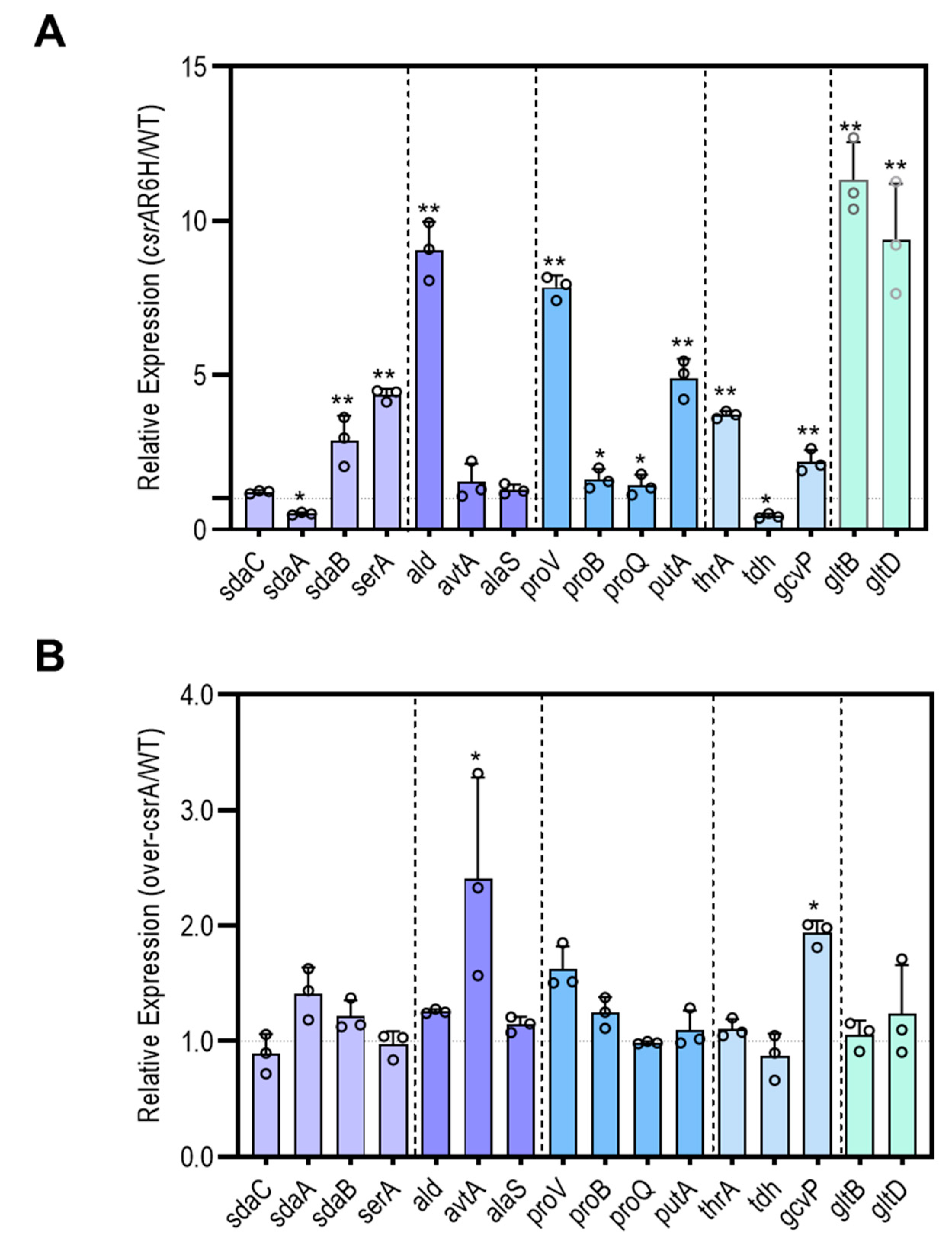
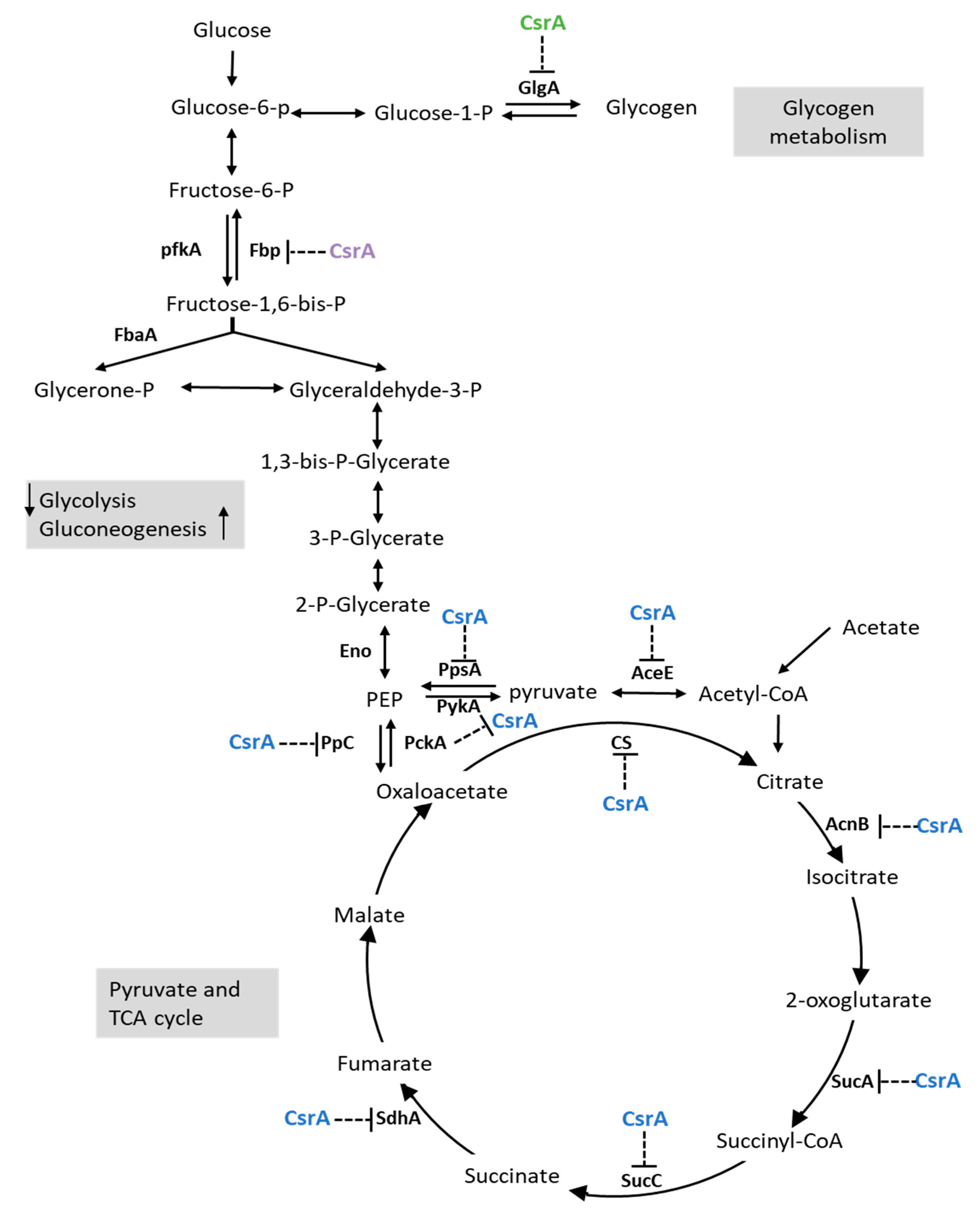
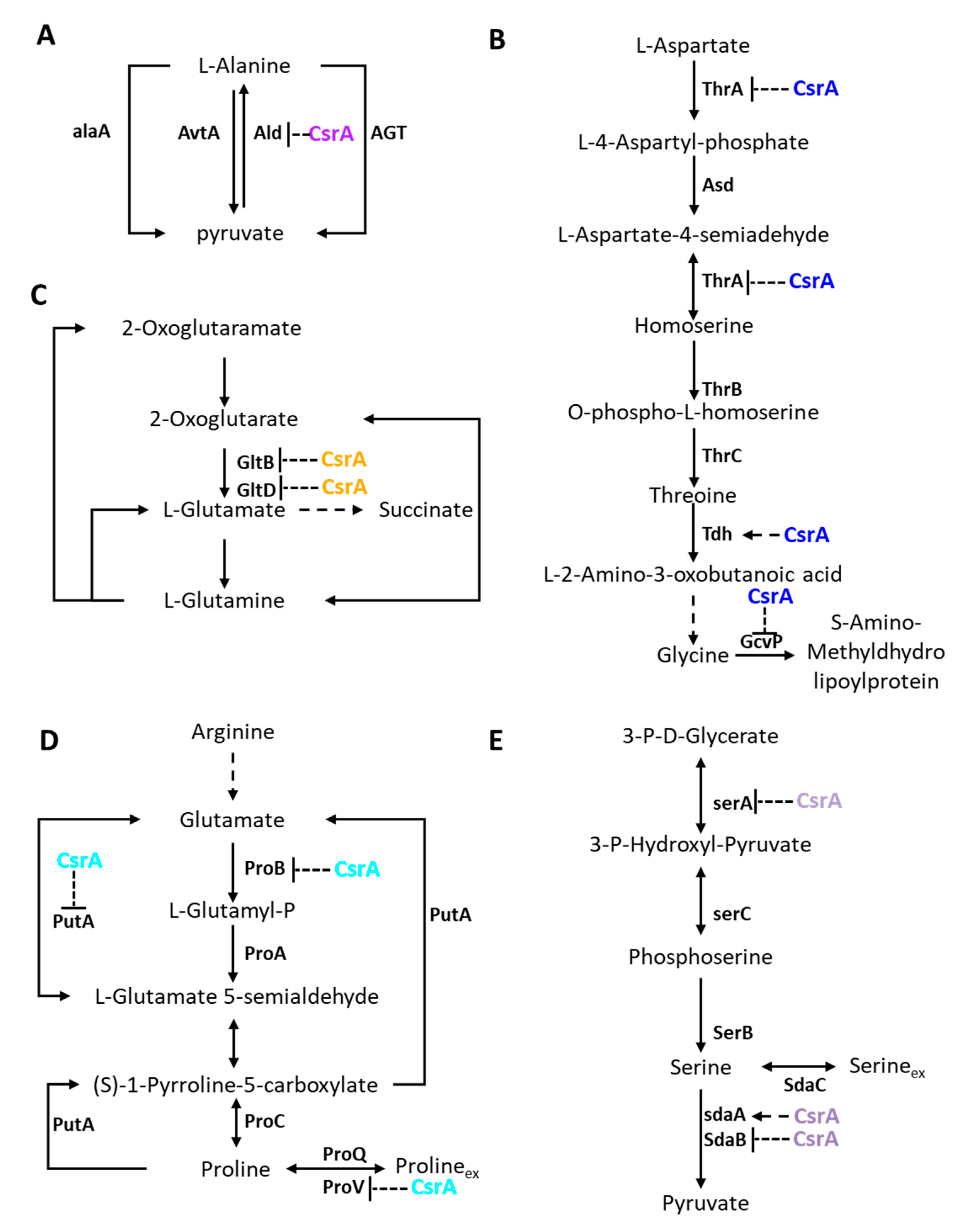
3.3. CsrA Is Extensively Involved in Carbon and Nitrogen Metabolism
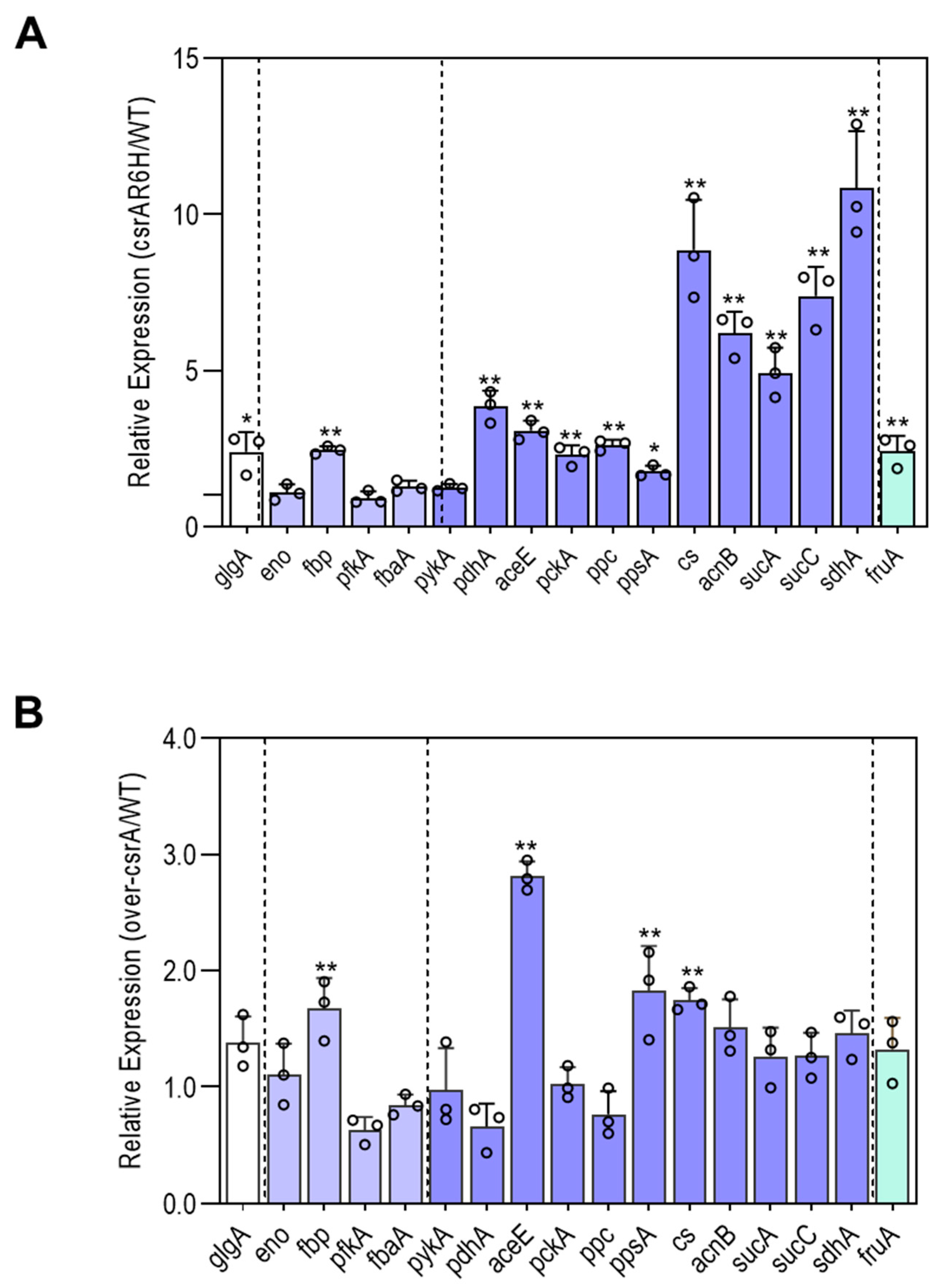
3.4. CsrA Has a Significant Impact on the Transcript Abundance of Genes Involved in Central Metabolism
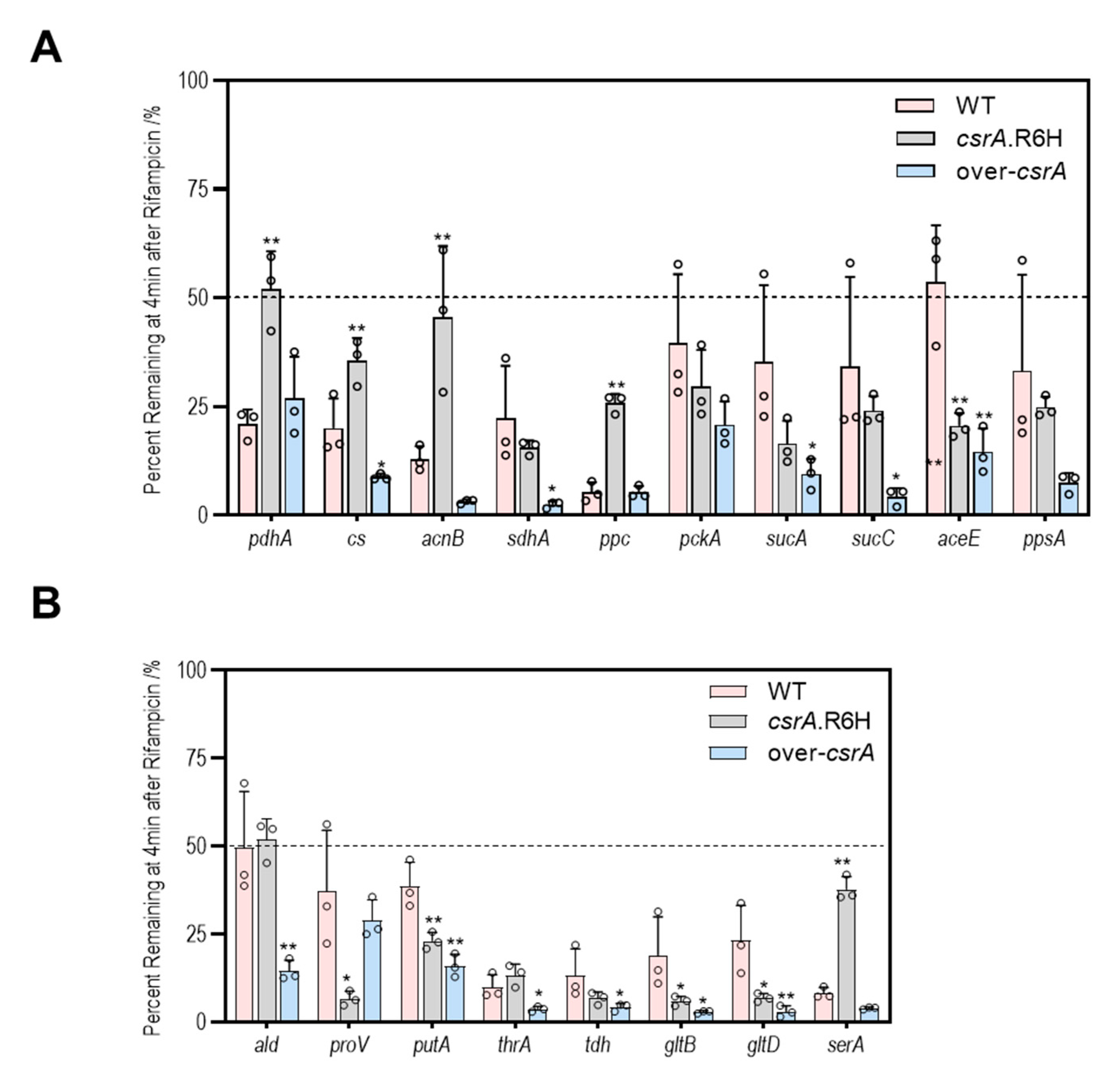
3.5. CsrA Changes the Transcipt Abundance of Genes Involved in Central Metabolism Likely by Alternation of Their mRNA Stability
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xie, J.; Bu, L.; Jin, S.; Wang, X.; Zhao, Q.; Zhou, S.; Xu, Y. Outbreak of vibriosis caused by Vibrio harveyi and Vibrio alginolyticus in farmed seahorse Hippocampus kuda in China. Aquaculture 2020, 523, 735168. [Google Scholar] [CrossRef]
- Xie, Z.; Ke, S.; Hu, C.; Zhu, Z.; Wang, S.; Zhou, Y. First Characterization of Bacterial Pathogen, Vibrio alginolyticus, for Porites andrewsi White Syndrome in the South China Sea. PLoS ONE 2013, 8, e75425. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.; Mcdonald, M.; Russ, G.; Carson, M.; Lukaczynski, E. Vibrio alginolyticus peritonitis associated with ambulatory peritoneal dialysis. Br. Med. J. 1981, 283, 275. [Google Scholar] [CrossRef] [Green Version]
- Opal, S.M.; Saxon, J.R. Intracranial infection by Vibrio alginolyticus following injury in salt water. J. Clin. Microbiol. 1986, 23, 373–374. [Google Scholar] [CrossRef] [Green Version]
- Barbarossa, V.; Kucisec-Tepes, N.; Aldova, E.; Matek, D.; Stipoljev, F. Ilizarov technique in the treatment of chronic osteomyelitis caused by Vibrio alginolyticus. Croat. Med. J. 2002, 43, 346–349. [Google Scholar]
- Chien, J.; Shih, J.; Hsueh, P.; Yang, P.; Luh, K. Vibrio alginolyticus as the cause of pleural empyema and bacteremia in an immunocompromised patient. Eur. J. Clin. Microbiol. Infect. Dis. 2002, 21, 401–403. [Google Scholar] [CrossRef]
- Feingold, M.H.; Kumar, M.L. Otitis media associated with Vibrio alginolyticus in a child with pressure-equalizing tubes. Pediatric Infect. Dis. J. 2004, 23, 475–476. [Google Scholar] [CrossRef]
- Li, X.C.; Xiang, Z.Y.; Xu, X.M.; Yan, W.H.; Ma, J.M. Endophthalmitis Caused by Vibrio alginolyticus. J. Clin. Microbiol. 2009, 47, 3379–3381. [Google Scholar] [CrossRef] [Green Version]
- Hong, G.L.; Dai, X.Q.; Lu, C.J.; Liu, J.M.; Zhao, G.J.; Wu, B.; Li, M.F.; Lu, Z.Q. Temporizing surgical management improves outcome in patients with Vibrio necrotizing fasciitis complicated with septic shock on admission. Burns 2014, 40, 446–454. [Google Scholar] [CrossRef]
- Pourciau, C.; Lai, Y.-J.; Gorelik, M.; Babitzke, P.; Romeo, T. Diverse Mechanisms and Circuitry for Global Regulation by the RNA-Binding Protein CsrA. Front. Microbiol. 2020, 11, 601352. [Google Scholar] [CrossRef]
- Sabnis, N.A.; Yang, H.; Romeo, T. Pleiotropic regulation of central carbohydrate metabolism in Escherichia coli via the gene csrA. J. Biol. Chem. 1995, 270, 29096–29104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altier, C.; Suyemoto, M.; Lawhon, S.D. Regulation of Salmonella enterica serovar typhimurium invasion genes by csrA. Infect. Immun. 2000, 68, 6790–6797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, D.W.; Suzuki, K.; Oakford, L.; Simecka, J.W.; Hart, M.E.; Romeo, T. Biofilm Formation and Dispersal under the Influence of the Global Regulator CsrA of Escherichia coli. J. Bacteriol. 2002, 184, 290–301. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Cui, Y.Y.; Liu, Y.; Dumenyo, C.K.; Chatterjee, A.K. Global regulation in Erwinia species by Erwinia carotovora rsmA, a homologue of Escherichia coli csrA: Repression of secondary metabolites, pathogenicity and hypersensitive reaction. Microbiology 1996, 142, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Romeo, T.; Gong, M.; Liu, M.Y.; Brunzinkernagel, A.M. Identification and molecular characterization of csrA a pleiotropic gene from Escherichia coli that affects glycogen biosynthesis, gluconeogenesis, cell-size, and surface-properties. J. Bacteriol. 1993, 175, 4744–4755. [Google Scholar] [CrossRef] [Green Version]
- Wei, B.; Brun-Zinkernagel, A.M.; Simecka, J.W.; Prüss, B.; Romeo, T. Positive regulation of motility and flhDC expression by the RNA-binding protein CsrA of Escherichia coli. Mol. Microbiol. 2010, 40, 245–256. [Google Scholar] [CrossRef]
- Gudapaty, S.; Suzuki, K.; Wang, X.; Babitzke, P.; Romeo, T. Regulatory Interactions of Csr Components: The RNA Binding Protein CsrA Activates csrB transcription in Escherichia coli. J. Bacteriol. 2001, 183, 6017–6027. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M. Towards bacterial strains overproducing L-tryptophan and other aromatics by metabolic engineering. Appl. Microbiol. Biotechnol. 2006, 69, 615. [Google Scholar] [CrossRef] [Green Version]
- Romeo, T.; Vakulskas, C.A.; Babitzke, P. Post-transcriptional regulation on a global scale: Form and function of Csr/Rsm systems. Environ. Microbiol. 2013, 15, 313–324. [Google Scholar] [CrossRef]
- Weilbacher, T.; Suzuki, K.; Dubey, A.K.; Wang, X.; Romeo, T. A novel sRNA component of the carbon storage regulatory system of Escherichia coli. Mol. Microbiol. 2003, 48, 657–670. [Google Scholar] [CrossRef]
- Mey, A.R.; Butz, H.A.; Payne, S.M. Vibrio cholerae CsrA Regulates ToxR Levels in Response to Amino Acids and Is Essential for Virulence. mBio 2015, 6, e01064-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butz, H.A.; Mey, A.R.; Ciosek, A.L.; Payne, S.M. Vibrio cholerae CsrA Directly Regulates varA to Increase Expression of the Three Nonredundant Csr Small RNAs. mBio 2019, 10, e01042-19. [Google Scholar] [CrossRef] [Green Version]
- Butz, H.A.; Mey, A.R.; Ciosek, A.L.; Crofts, A.A.; Davies, B.W.; Payne, S.M. Regulatory Effects of CsrA in Vibrio cholerae. mBio 2021, 12, e03380-20. [Google Scholar] [CrossRef] [PubMed]
- Val, M.E.; Skovgaard, O.; Ducos-Galand, M.; Bland, M.J.; Mazel, D. Genome engineering in Vibrio cholerae: A feasible approach to address biological issues. PLoS Genet. 2012, 8, e1002472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neidhardt, F.C.; Bloch, P.L.; Smith, D.F. Culture Medium for Enterobacteria. J. Bacteriol. 1974, 119, 736–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.; Jin, X.; Hu, C. Phenotypic and genetic differences between opaque and translucent colonies of Vibrio alginolyticus. Biofouling 2009, 25, 525–531. [Google Scholar] [CrossRef]
- Nguyen, A.N.; Disconzi, E.; Charriere, G.M.; Destoumieux-Garzon, D.; Bouloc, P.; Le Roux, F.; Jacq, A. csrB Gene Duplication Drives the Evolution of Redundant Regulatory Pathways Controlling Expression of the Major Toxic Secreted Metalloproteases in Vibrio tasmaniensis LGP32. mSphere 2018, 3, e00582-18. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-J.; Chen, G.; Lin, H.; Wang, P.; Kuang, B.; Liu, J.; Chen, S. Development of a regulatable expression system for the functional study of Vibrio vulnificus essential genes. Antonie Leeuwenhoek 2017, 110, 607–614. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ulitzur, S. The mechanism of swarming of Vibrio alginolyticus. Arch. Microbiol. 1975, 104, 67–71. [Google Scholar] [CrossRef]
- Atsumi, T.; Maekawa, Y.; Yamada, T.; Kawagishi, I.; Imae, Y.; Homma, M. Effect of viscosity on swimming by the lateral and polar flagella of Vibrio alginolyticus. J. Bacteriol. 1996, 178, 5024–5026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogure, K.; Ikemoto, E.; Morisaki, H. Attachment of Vibrio alginolyticus to glass surfaces is dependent on swimming speed. J. Bacteriol. 1998, 180, 932–937. [Google Scholar] [CrossRef] [Green Version]
- Lenz, D.H.; Miller, M.B.; Zhu, J.; Kulkarni, R.V.; Bassler, B.L. CsrA and three redundant small RNAs regulate quorum sensing in Vibrio cholerae. Mol. Microbiol. 2005, 58, 1186–1202. [Google Scholar] [CrossRef] [PubMed]
- Mey, A.R.; Craig, S.A.; Payne, S.M. Effects of Amino Acid Supplementation on Porin Expression and ToxR Levels in Vibrio cholerae. Infect. Immun. 2012, 80, 518–528. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.K.; Warner, E.B.; Oliver, J.D. csrA Inhibits the Formation of Biofilms by Vibrio vulnificus. Appl. Environ. Microbiol. 2008, 74, 7064–7066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakhnin, A.V.; Baker, C.S.; Vakulskas, C.A.; Yakhnin, H.; Babitzke, P. CsrA activates flhDC expression by protecting flhDC mRNA from RNase E-mediated cleavage. Mol. Microbiol. 2013, 87, 851–866. [Google Scholar] [CrossRef] [Green Version]
- Weinstock, M.T.; Hesek, E.D.; Wilson, C.M.; Gibson, D.G. Vibrio natriegens as a fast-growing host for molecular biology. Nat. Methods 2016, 13, 849–851. [Google Scholar] [CrossRef]
- Jacobsen, T.; Koch, A.G. Influence of different histories of the inoculum on lag phase and growth of Listeria monocytogenes in meat models. J. Food Prot. 2006, 69, 532–541. [Google Scholar] [CrossRef]
- Yates, G.T.; Smotzer, T. On the lag phase and initial decline of microbial growth curves. J. Theor. Biol. 2007, 244, 511–517. [Google Scholar] [CrossRef]
- Bertrand, R.L. Lag Phase Is a Dynamic, Organized, Adaptive, and Evolvable Period That Prepares Bacteria for Cell Division. J. Bacteriol. 2019, 201, e00697-18. [Google Scholar] [CrossRef]
- Yamamotoya, T.; Dose, H.; Tian, Z.Y.; Faure, A.; Toya, Y.; Honma, M.; Igarashi, K.; Nakahigashi, K.; Soga, T.; Mori, H.; et al. Glycogen is the primary source of glucose during the lag phase of E. coli proliferation. Biochim. Biophys. Acta-Proteins Proteom. 2012, 1824, 1442–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potts, A.H.; Vakulskas, C.A.; Pannuri, A.; Yakhnin, H.; Babitzke, P.; Romeo, T. Global role of the bacterial post-transcriptional regulator CsrA revealed by integrated transcriptomics. Nat. Commun. 2017, 8, 1596. [Google Scholar] [CrossRef] [Green Version]
- Morin, M.; Ropers, D.; Letisse, F.; Laguerre, S.; Portais, J.-C.; Cocaign-Bousquet, M.; Enjalbert, B. The post-transcriptional regulatory system CSR controls the balance of metabolic pools in upper glycolysis of Escherichia coli. Mol. Microbiol. 2016, 100, 686–700. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.B.; Peng, B.; Li, H.; Cheng, Z.X.; Zhang, T.T.; Zhu, J.X.; Li, D.; Li, M.Y.; Ye, J.Z.; Du, C.C.; et al. Pyruvate cycle increases aminoglycoside efficacy and provides respiratory energy in bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, E1578–E1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durica-Mitic, S.; Goepel, Y.; Goerke, B. Carbohydrate Utilization in Bacteria: Making the Most Out of Sugars with the Help of Small Regulatory RNAs. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Hauslein, I.; Sahr, T.; Escoll, P.; Klausner, N.; Eisenreich, W.; Buchrieser, C. Legionella pneumophila CsrA regulates a metabolic switch from amino acid to glycerolipid metabolism. Open Biol. 2017, 7, 170149. [Google Scholar] [CrossRef] [Green Version]
- Lawhon, S.D.; Frye, J.G.; Suyemoto, M.; Porwollik, S.; McClelland, M.; Altier, C. Global regulation by CsrA in Salmonella typhimurium. Mol. Microbiol. 2003, 48, 1633–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brencic, A.; Lory, S. Determination of the regulon and identification of novel mRNA targets of Pseudomonas aeruginosa RsmA. Mol. Microbiol. 2009, 72, 612–632. [Google Scholar] [CrossRef] [Green Version]
- Potts, A.H.; Guo, Y.; Ahmer, B.M.M.; Romeo, T. Role of CsrA in stress responses and metabolism important for Salmonella virulence revealed by integrated transcriptomics. PLoS ONE 2019, 14, e0211430. [Google Scholar] [CrossRef] [Green Version]
- Esquerre, T.; Bouvier, M.; Turlan, C.; Carpousis, A.J.; Girbal, L.; Cocaign-Bousquet, M. The Csr system regulates genome-wide mRNA stability and transcription and thus gene expression in Escherichia coli. Sci. Rep. 2016, 6, 25057. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
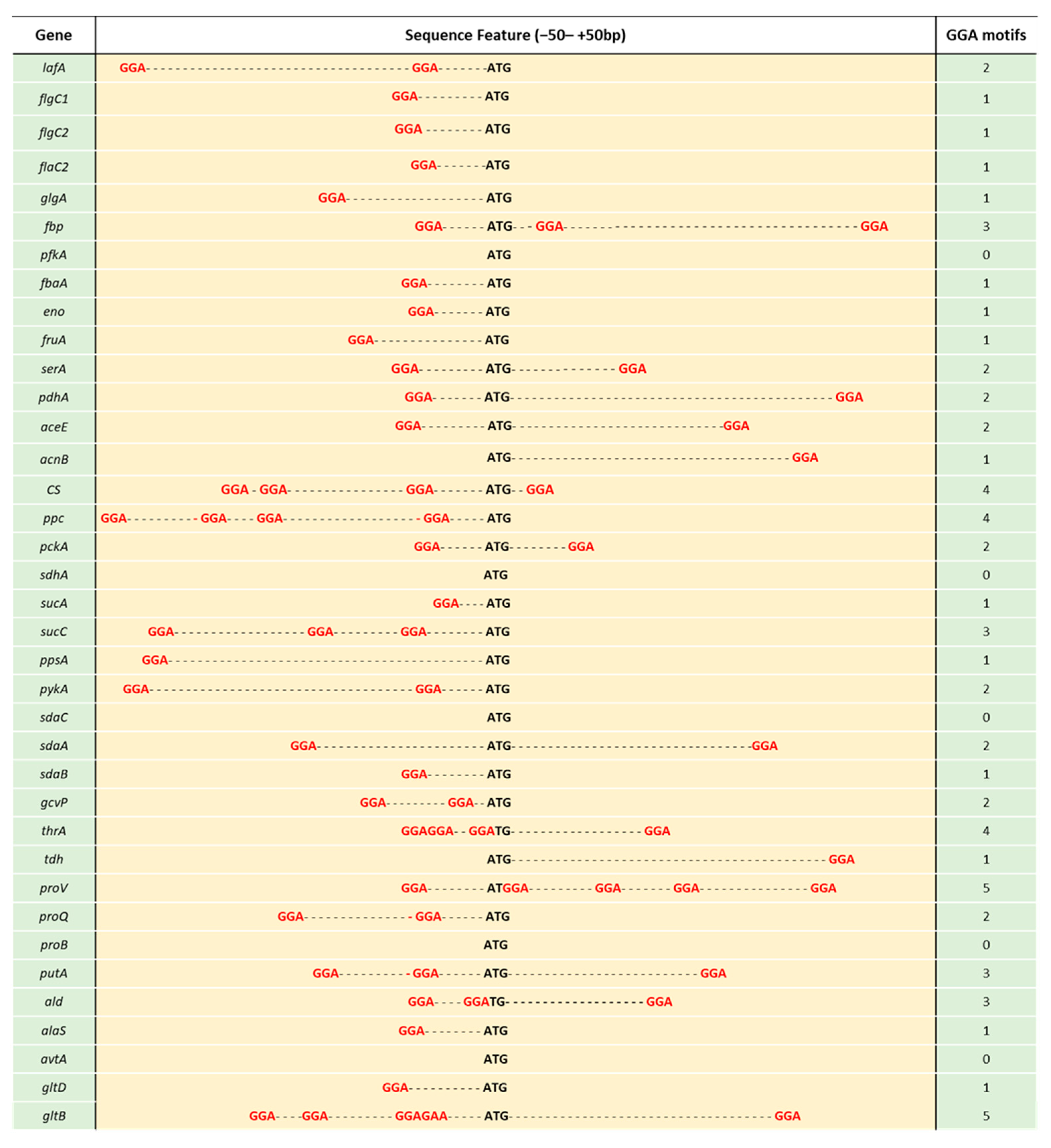{kind=link}
| Strains or Plasmids | Relevant Characteristics | Source |
|---|---|---|
| Vibrio alginolyticus | ||
| ZJ_T | Apr (ampicillin-resistant), translucent/smooth variant of wild strain ZJ51; isolated from diseased Epinephelus coioides off the Southern China coast | [26] |
| ZJ_T-csrA R6H | Apr; ZJ-T carrying a point mutation that replaces the arginine residue at amino acid position 6 with a histidine (R6H). | This study |
| ZJ_T/over-csrA-pSCT32 | Cmr; ZJ-T carrying the CsrA expression plasmid pSCT32- over-csrA | This study |
| E. coli | ||
| GEB883 | WT; E.coli K12 ΔdapA::ermpir RP4-2 ΔrecA gyrA462, zei298::Tn10; donor strain for conjugation | [27] |
| Plasmids | ||
| pSW7848 | Cmr; suicide vector with an R6K origin, requiring the Pir protein for its replication, and the ccdB toxin gene | [24] |
| pSW7848-CsrAR6H | Cmr; pSW7848 containing the mutant allele of CsrA | This study |
| pSCT32 | Cmr; expression plasmid with a pBR322 and a f1 origin at the same time and a tac promoter | [28] |
| pSCT32-over csrA | Cmr; pSCT32 containing the WT allele of CsrA | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Gao, Q.; Zhang, X.; Chen, H.; Zhang, Y.; Sun, Y.; Yang, S.; Chen, C. CsrA Regulates Swarming Motility and Carbohydrate and Amino Acid Metabolism in Vibrio alginolyticus. Microorganisms 2021, 9, 2383. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112383
Liu B, Gao Q, Zhang X, Chen H, Zhang Y, Sun Y, Yang S, Chen C. CsrA Regulates Swarming Motility and Carbohydrate and Amino Acid Metabolism in Vibrio alginolyticus. Microorganisms. 2021; 9(11):2383. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112383
Chicago/Turabian StyleLiu, Bing, Qian Gao, Xin Zhang, Huizhen Chen, Ying Zhang, Yuehong Sun, Shan Yang, and Chang Chen. 2021. "CsrA Regulates Swarming Motility and Carbohydrate and Amino Acid Metabolism in Vibrio alginolyticus" Microorganisms 9, no. 11: 2383. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112383