1. Introduction
Plastic materials that have been generally used in our daily lives are now causing dangerous environmental problems. Millions of tons of these non-degradable plastics accumulate in the environment per year. Petroleum-based plastics have serious ecological and social impacts because of their non-degradable nature and the leaching of carcinogenic substances when exposed to scratch or heat [
1]. Biopolymers are one type of product that can help to overcome problems caused by petrochemical polymers. Biopolymers are generated from renewable natural sources and are often biodegradable and nontoxic. They are either produced by biological systems (microorganisms, plants, and animals) or produced from substrates obtained from living organisms such as polylactic acid, which can be synthesized from biologically obtained lactic acid [
2]. Among various biodegradable polymer materials are polyhydroxyalkanoates (PHAs) [
3]. PHAs are thermoplastic aliphatic polyesters with linear polymer chains that are manufactured via a microbial process on sugar-based medium, where they act as carbon and energy storage material in bacteria.
The main member of the PHAs family is polyhydroxybutyrate (PHB). It is accumulated in the cells as intracellular granules in the presence of excess carbon source or under different stress conditions, such as a limited amount of macro-components (nitrogen, phosphate, oxygen) or micro-components (sulfate, magnesium ions, and other trace elements) [
4]. Bacterial PHAs could be divided into two groups depending on the number of carbon atoms in the monomeric units: short-chain-length (SCL) PHAs, which consist of 3–5 carbon atoms, and medium-chain-length (MCL) PHAs, which consist of 6–14 carbon atoms [
5].
PHAs are non-toxic, biodegradable, and biocompatible, so they can be used in medical applications as bioimplant materials and in encapsulation of medicines for controlled release. It can be also used in other applications such as bags, bottles, disposable items, items of personal hygiene, films, and food packaging. PHAs degrade naturally and completely to CO
2 and H
2O in a natural environment due to different microorganisms [
6]. Alarfaj et al. [
7] proved the ability of the
B. thuringiensis KSBM 127 strain that was isolated from the mangroves environment of Saudi Arabia to produce biodegradable plastics. The potential of isolated bacteria from the Makkah region, Saudi Arabia, to produce PHBs and the factors affecting the production of PHB were studied by Alshehrei [
8].
A major problem for PHBs production is their high production cost as compared with plastics derived from petrochemicals. Therefore, much effort has been committed to reducing the production cost of PHBs by using low-cost carbon sources [
9].
Bacillus axaraqunsis BIPC01, which was isolated from Bandar Imam, Iran, could be a potent PHB producer using petrochemical wastewater as a carbon source [
10].
Some studies in Egypt were performed on PHBs isolation and characterization. Belal and Farid [
11] isolated
B.cereus from Kafr El-sheikh Governorate, Egypt, which has the ability to produce PHBs when inoculated in production media supplemented with 2% of glucose, xylose, lactose, whey, molasses, sugar cane bagasse, and rice straw hydrolysate. Rice bran, wheat bran, bagasse, cassava powder, potato starch, cassava powder, corn waste, copra oil cake, jack fruit powder, toor powder, fruit waste, and whey wastewater from industrial farming were used to form a low-cost substrate for PHBs production when inoculated with
B. cereus [
12]. Mostafa and his coworkers [
13] isolated novel strains of PHB-producing bacteria (
Tamlana crocina,
Bacillus aquimaris,
Erythrobacter aquimaris, and
Halomonas halophila) from the mangrove rhizosphere, Red Sea, Saudi Arabia, which can make PHB production a decent contender for numerous industrial applications as a substitution for petroleum-based plastics. Rezk et al. (2020) [
14] proved that wheat bran can be used as an alternative to starch nitrate medium for PHB production by
Streptomyces incanus BK128.
Therefore, the aim of the present study was to optimize and characterize PHB produced by a new bacterial stain (Bacillus wiedmanii) with a focus on its production from some low-cost agricultural wastes (fruit peels).
2. Materials and Methods
2.1. Sample Collection and Isolation of PHB-Producing Bacteria
Different samples (soil, wastewater, sewage sludge, and cattle manure) were collected from various sources in Assiut Governorate, Egypt, and used for bacterial isolation. One gram (soil, cattle manure, or sludge samples) or 1 mL (wastewater samples) was added to 9 mL of sterilized distilled water. Samples were subjected to shaking for 30 min on a rotary shaker (150 rpm) at 30 °C. Then, serial dilutions were prepared, followed by streaking on nutrient agar plates. The plates were incubated at 35 °C for 24 h. The isolated strains were purified and maintained on nutrient slant agar and stored at 4 °C.
2.2. Screening the Isolates for PHB Production
The isolated strains were screened for the presence of polyhydroxybutyrate (PHB) granules using the Sudan Black B staining technique. Nutrient agar medium was supplemented with 2% glucose. The plate was divided into equal parts, and in each part, bacterial isolates were spread. The plates were incubated at 30 °C for 24 h. Sudan Black B stain was prepared by dissolution of 0.02 g powdered stain in 100 mL of 70% ethanol. After incubation, Sudan Black B dye was spread over the plates and kept undisturbed for 30 min. Plates were washed with ethanol (96%) to remove the excess stain. Colonies unable to incorporate the Sudan Black B appeared white, while PHB producers appeared bluish-black [
15]. The promising isolate for PHB production was genetically identified by morphological and biochemical test and 16S rRNA gene analysis.
2.3. Morphological and Biochemical Analysis
The isolate was morphologically characterized by observing the standard microbiological markers (Gram reaction, motility, spore formation). The biochemical characterization of the isolate was done by series of biochemical tests including carbohydrate fermentation, H2S production, and catalase test. Oxidase activities and deoxidization of nitrate were also examined.
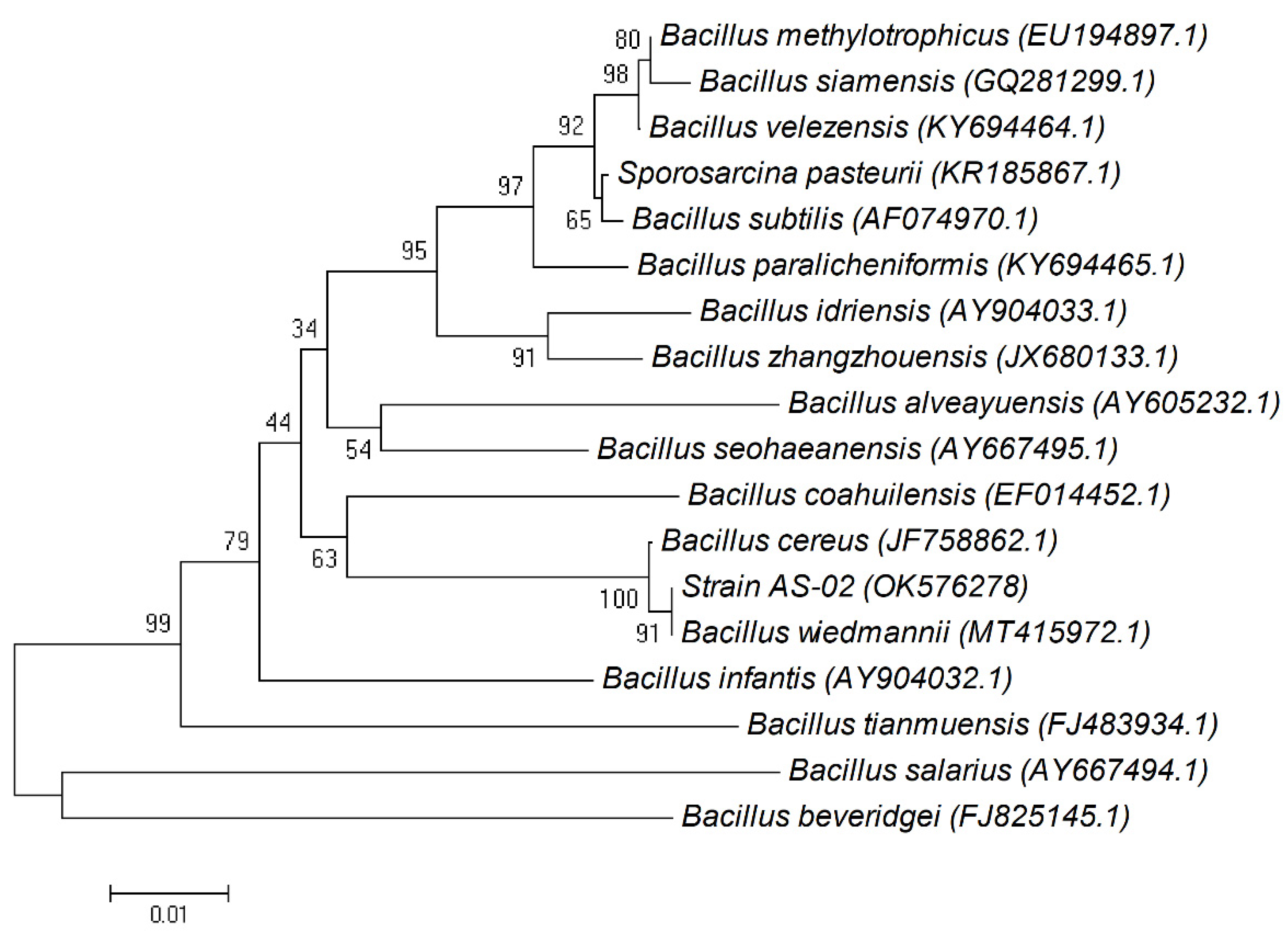
2.4. Molecular Identification of PHB-Accumulating Bacteria by 16S rRNA Gene Analysis
The identification was performed based on 16S rRNA gene sequence analysis. Genomic DNA was extracted from the isolated bacterium strain according to our previously described method [
16]. PCR amplification was performed using common primers: 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-CGGCTACCTTGTTACGACTT-3′). The sequences obtained were then aligned with known 16S rRNA gene sequences in GenBank database using the basic local alignment search tool (BLAST) of the National Center for Biotechnology Information (
http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/BLAST/, accessed on 20 September 2021). To determine the taxonomic position of the isolates, the phylogenetic tree was constructed with MEGA version 4.0 (Auckland, New Zealand) using a neighbor-joining algorithm, and the Jukes–Cantor distance estimation method with bootstrap analyses for 100 replicates was performed [
17].
2.5. Production, Detection, and Extraction of PHB
The pure culture of the promising isolate was inoculated in 5 mL of sterile nutrient broth media. After incubation for 24 h at 30°C, 5% (
v/
v) of the culture was aseptically transferred into 250 mL conical flask containing 50 mL of modified mineral salts medium (pH 7.0) containing (in g/L) 20 glucose, 0.2 MgSO
4, 0.1 NaCl, 0.5 KH
2PO
4, 4.0 peptone, and 2.5 yeast extract [
18]. Then, it was incubated for 72 h at 30 °C and 150 rpm. The culture broth was centrifuged (MPW-260 Refrigerated Laboratory Centrifuge, MPW Med., Warsaw, Poland) at 5000 rpm for 15 min. The supernatant was discarded, and the pellet was dried. Sodium hypochlorite (10 mL) was added to the dried pellet and then incubated for 2 h at 50 °C for lyses of cells. After the incubation period, the tube with the mixture was centrifuged again at 5000 rpm for 15 min. The pellet was washed with distilled water, acetone, and methanol; then, it was dissolved by 5 mL of boiling chloroform. The non-PHB cell matter was removed by filtration using filter paper (Whatman no. 1, WHA1001045, Buckinghamshire, UK). The chloroform was evaporated, and PHB film was stored for further analysis [
12].
2.6. Quantitative Analysis of PHB
Cell culture was grown as described earlier, and the cell pellet was dried using DZF-6020 Laboratory vacuum dry oven over night at 60 °C to estimate the dry cell weight (DCW) in units of g/L. The percentage of intracellular PHBs accumulation is estimated as the percentage composition of PHB present in the dry cell weight [
15].
PHBs accumulation (%) = Dry weight of extracted PHB (g/L)/DCW (g/L) × 100.
2.7. Characterization of PHB
The purified PHB was characterized by the following analytical methods.
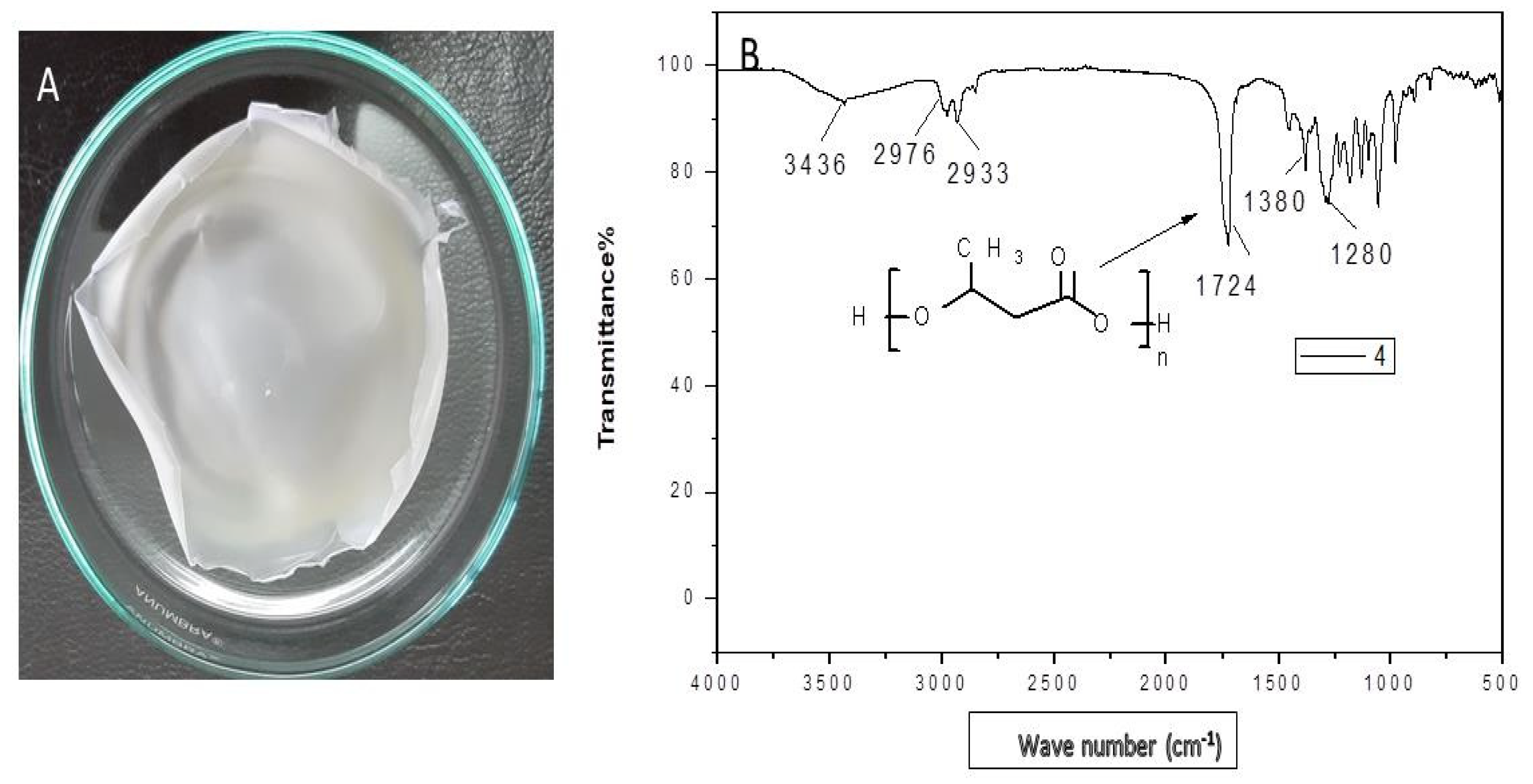
2.7.1. Fourier Transform Infrared Spectroscopy (FTIR)
The instrument used for this analysis was a Nicollet 6700 FTIR spectrophotometer (Assiut University, Egypt, Thermo Fisher Scientific, 168 Third Avenue, Waltham, MA 02451, USA). PHB sample was mixed with potassium bromide (KBr) in a sufficient ratio, and then it was ground. The pellet was kept in the sample holder, and IR rays were passed through it at a range of 4000–400 cm−1. Obtained results were analyzed for the determination of functional groups.
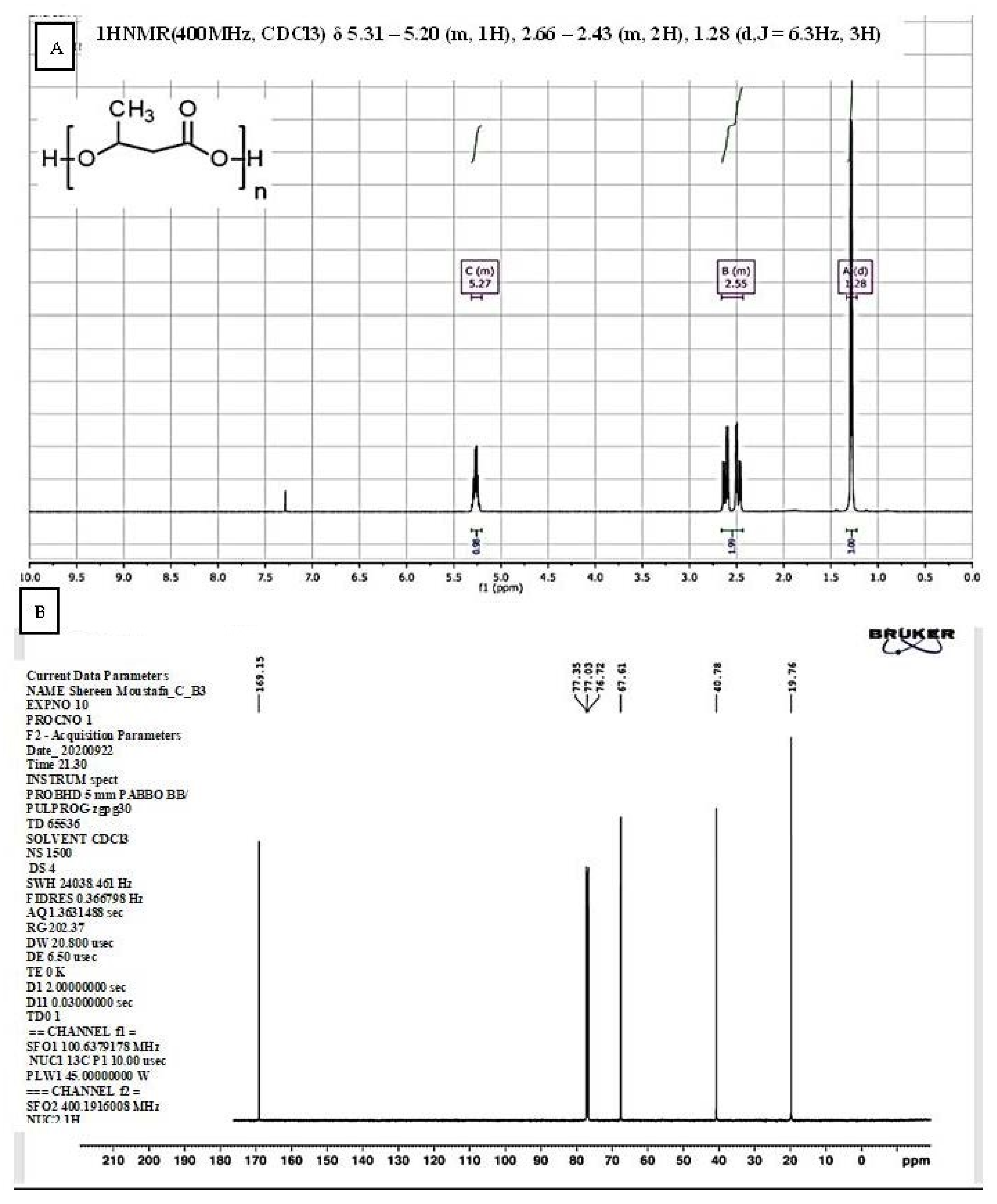
2.7.2. Nuclear Magnetic Resonance (NMR) Spectroscopy
The spectra were recorded for the extracted PHB sample using deuterated chloroform (CDCl3). Five milligrams of the extracted PHB was dissolved in deuterated chloroform (CDCl3), and the solution was transferred to a 5 mm NMR tube for nuclear magnetic resonance measurements. Tetramethyl saline (TMS) was used as an external reference. The instrument used for analysis was Bruker High-Performance Digital FT-NMR Spectrometer Avance (Bruker BioSpin, GmbH, Rheinstetten, Germany) with 400 MHz proton frequency, at the NMR Unit, Faculty of Pharmacy (Cairo University), and operating at the basic frequency of 400.13 MHz for 1H. The spectrometer is equipped with direct-detection broadband observe (BBO) probe. All NMR measurements were acquired at 298 K (25 °C). Data were analyzed using Topspin 3.1 software (Bruker Biosoin, Rheinstetten, Germany). Chemical shifts (δ) are expressed in ppm with reference to the residual solvent signals. Scalar coupling constants (J) are given in Hertz. The following conditions were used for recording of 1H NMR and 13C-NMR spectra: 30 °C pulse experiment; acquisition time of 4.1 s; relaxation delay 1.0 s; sweep width 15.1 ppm (8012 Hz); data points 65536; and dummy scan 2. The data were processed using line broadening 0.1 Hz.
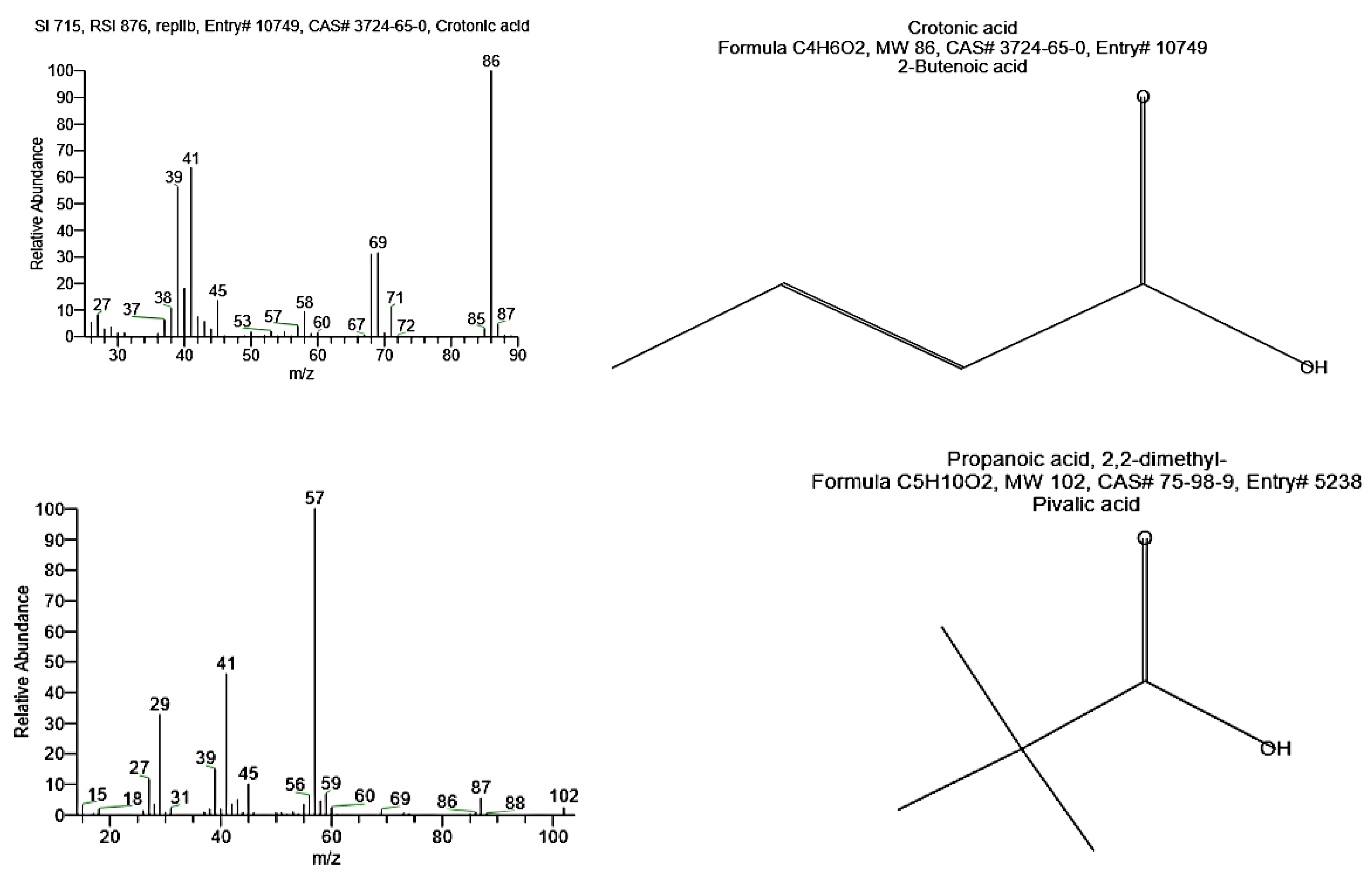
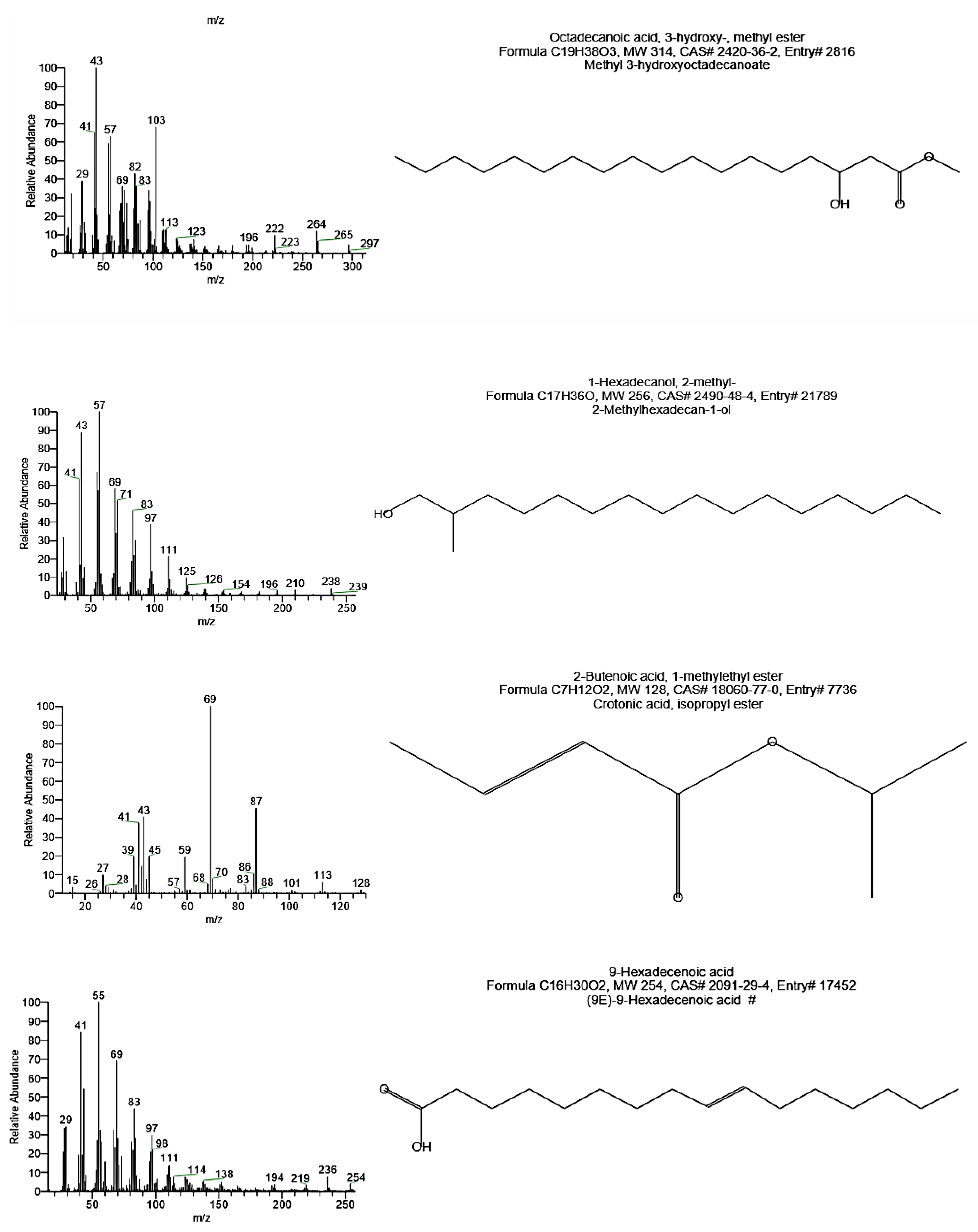
2.7.3. GC-MS Analysis
GC-MS analysis of the sample was carried out after methanolysis. PHB was suspended in 1.0 mL chloroform and 1.0 mL H
2SO
4/methanol (15:85) in a screw-capped tube and then heated to 100 °C for 2 h. After cooling, 0.5 mL of demineralized water was added, and the solution was vortexed for 1 min [
13]. The organic phase containing the resulting methyl esters’ monomers was analyzed using Gas Chromatography-Mass Spectrometric (Varian, CP–3800 GC and Saturn 2200 MS) at Analytical Chemistry Unit, Faculty of Science, Assiut University, Egypt. 7102 Riverwood Drive, Columbia, MD 21046-2502, USA that equipped with a quadrupole ion trap mass detector coupled with CP–Sil 5 CB (0.25 mm i.d × 30 m length) capillary column. Ionization energy was 70 eV, scan interval was 1.5 s, and mass range was from 50–600 amu. The oven temperature was programmed at 50 °C for 1.0 min with 10 °C increments and held at 280 °C for 10 min. The detector and injector temperature was kept at 280 °C, and helium was used as the carrier gas.
2.8. Optimization of Physic-Chemical Parameters for Production of PHB
The influence of various time courses, pH, temperature, different nitrogen and carbon sources, and C/N ratio on PHB production was investigated using nutrient broth media.
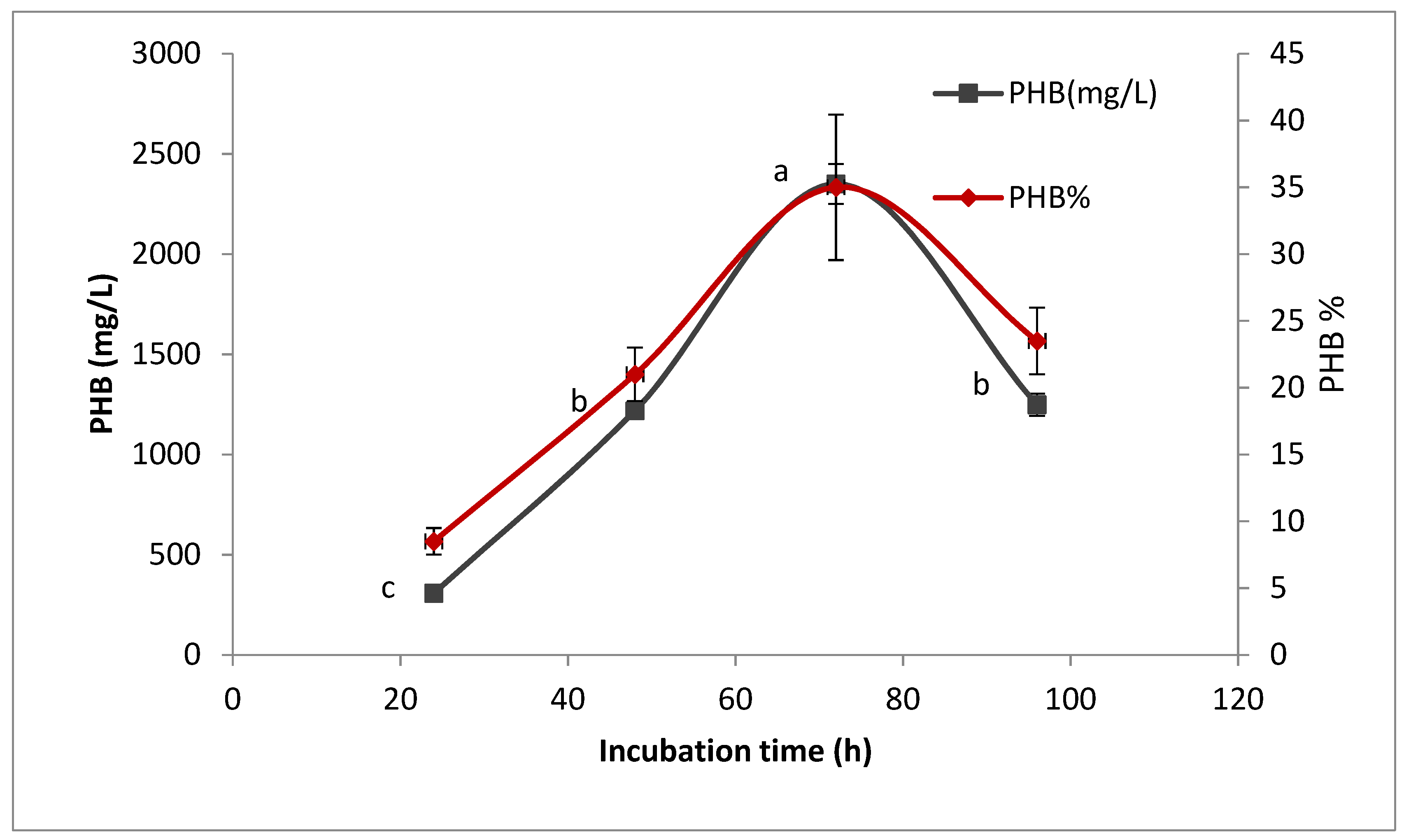
2.8.1. Effect of Incubation Time on PHB Production
To determine the best incubation time for PHB production, 200 mL of sterile production medium with (pH 7) was prepared and inoculated with 5% inoculums. The inoculated media was incubated at 30 °C, with shaking at 150 rpm [
18]. In total, 50 mL of culture was taken periodically at 24 h intervals up to 96 h. The PHB production was determined as described previously.
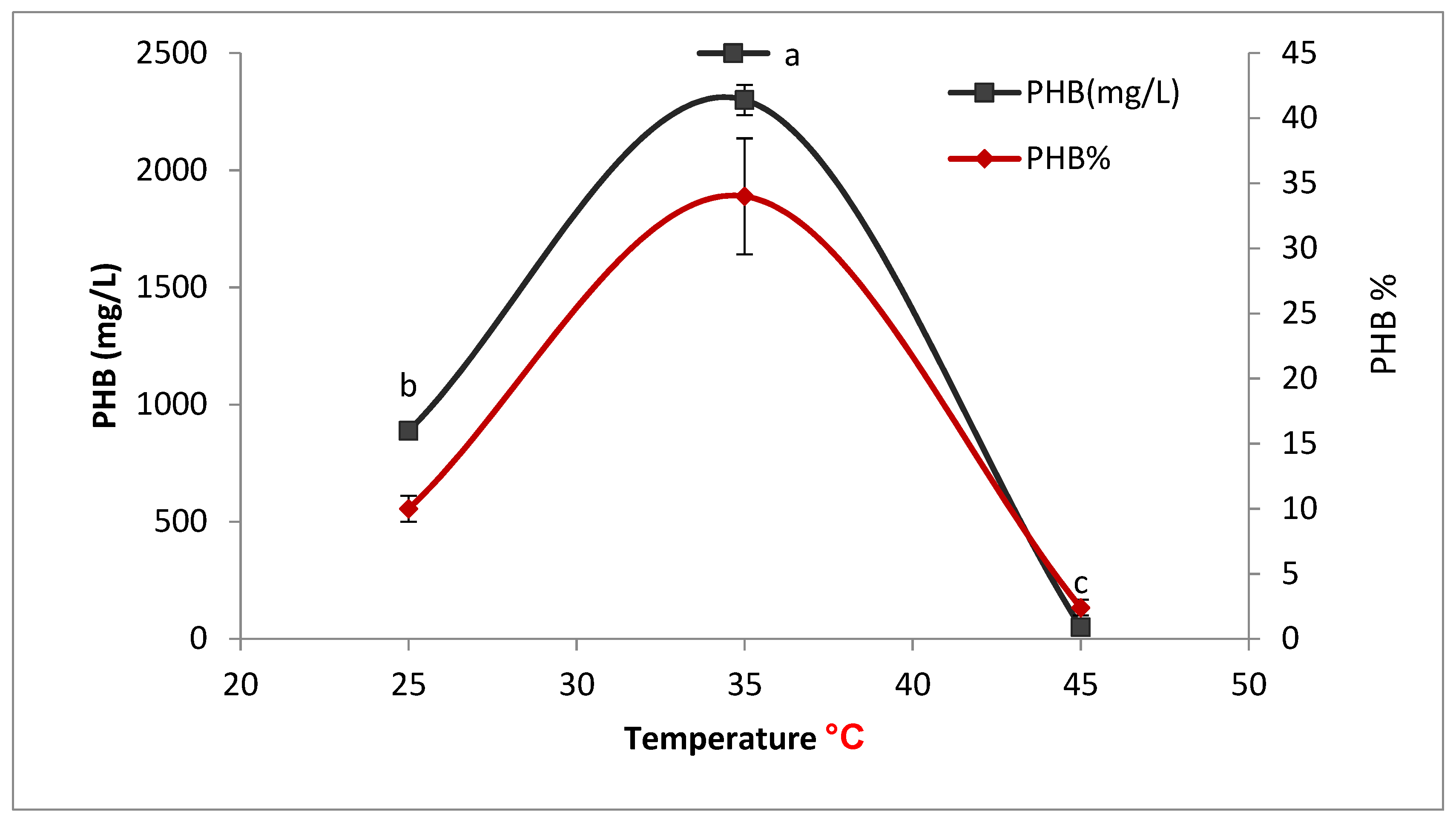
2.8.2. Effect of Temperature on PHB Production
To determine the optimum temperature for PHB production, 50 mL of sterile production media with (pH 7) was prepared in a different conical flask and inoculated with 5% inoculums. Each flask was incubated for 72 h at different temperatures (25, 35, and 45 °C).
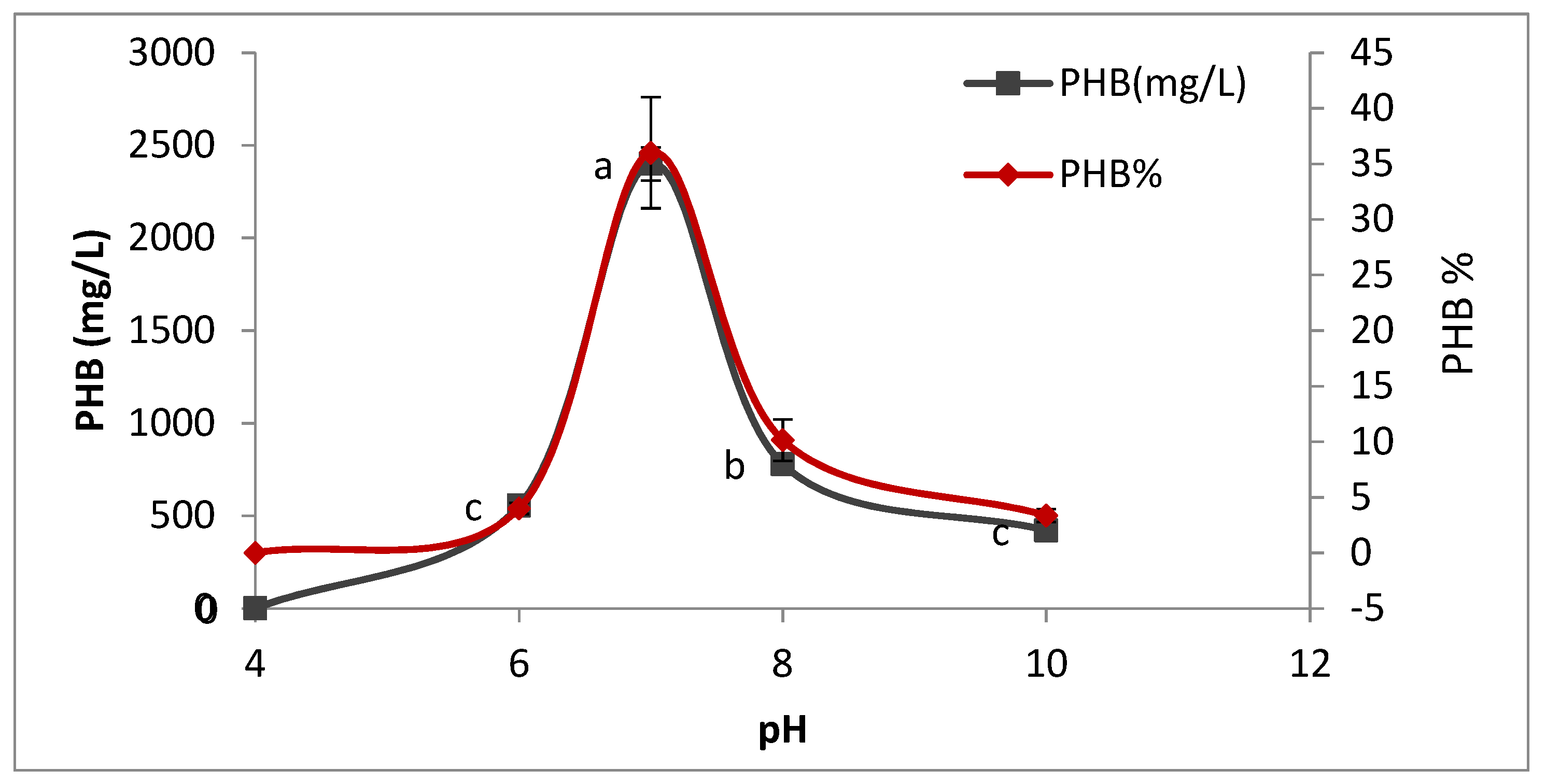
2.8.3. Effect of pH on PHB Production
To determine the optimum (pH) for PHB production, 50 mL of sterile production media was prepared in different conical flasks, and each flask was adjusted to different pH (4, 6, 7, 8, and 10) using 0.1 N NaOH and 0.1 N HCl. The flasks were inoculated with 5% inoculum and incubated for 72 h at 35 °C after sterilization. The PHB production was estimated as described previously.
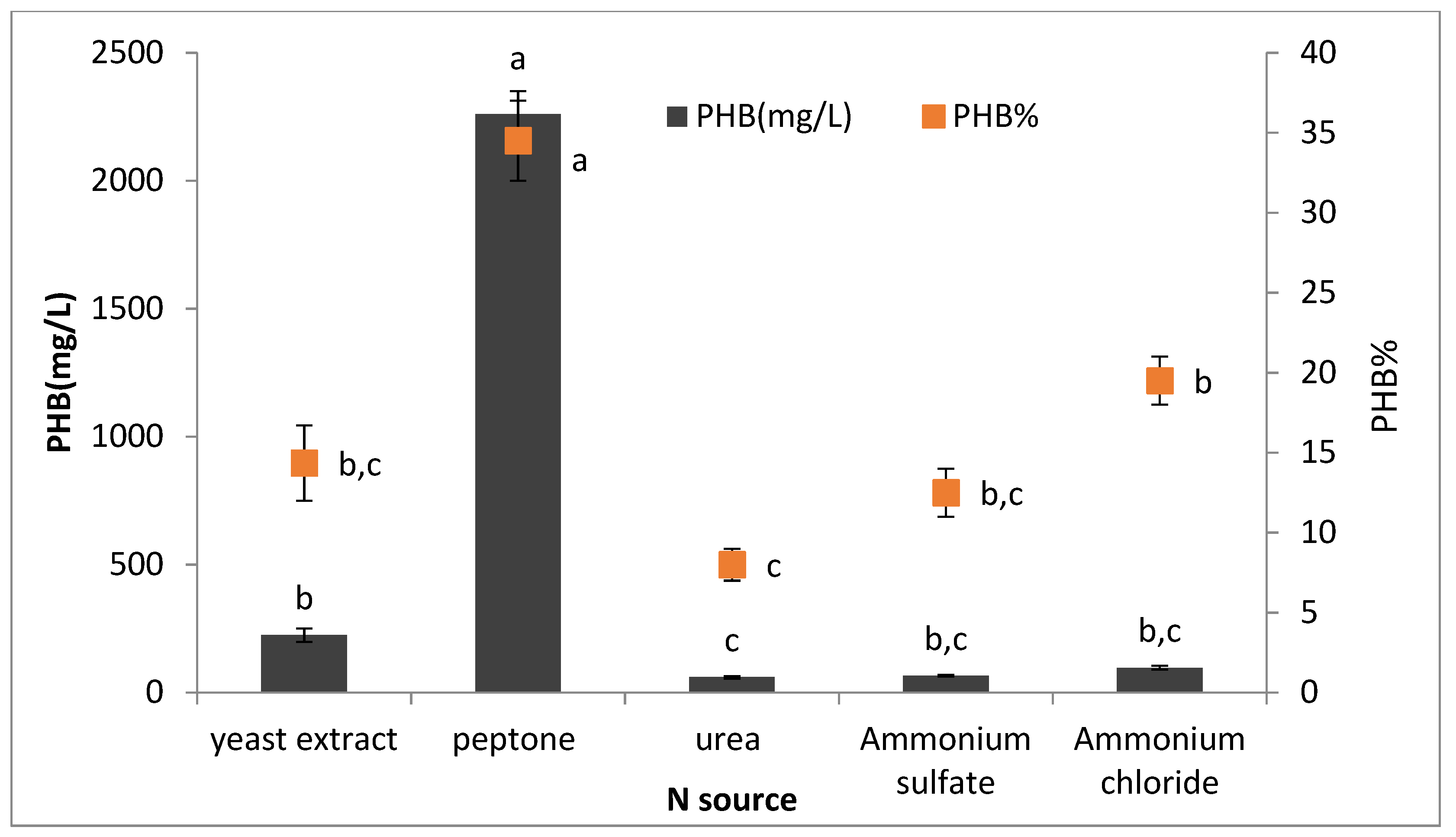
2.8.4. Effect of Different Nitrogen Sources on PHB Production
To detect the best nitrogen source for PHB production, 50 mL of sterile production media was prepared in different conical flasks supplemented with different nitrogen sources (peptone, yeast extract, urea, ammonium sulfate, and ammonium chloride) at 0.4% concentration. All flasks were inoculated with 5% of inoculum and incubated for 72 h at 35 °C after sterilization. The PHB production was estimated as described previously.
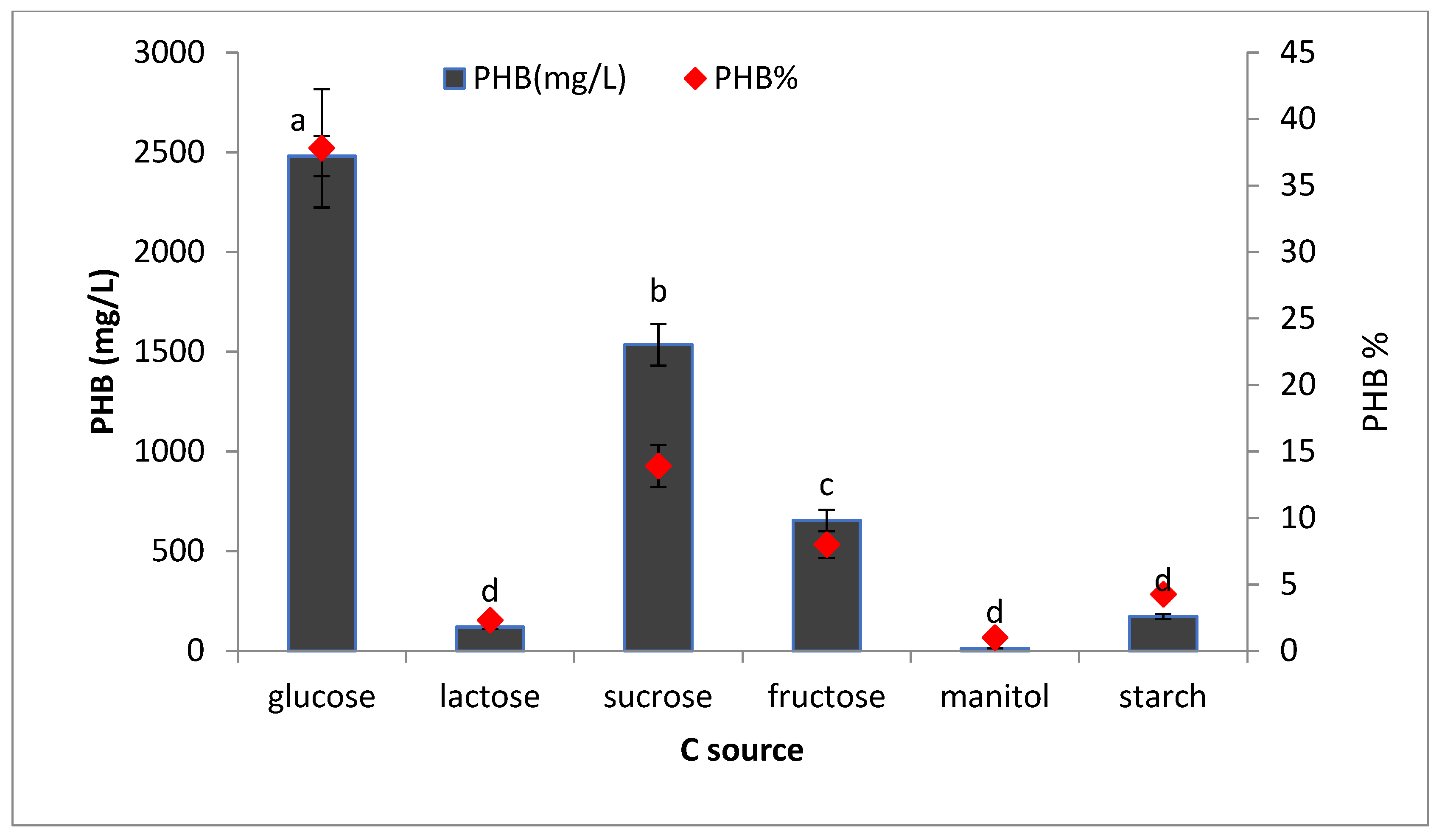
2.8.5. Effect of Different Carbon Sources on PHB Production
To find out the best carbon source for PHB production, 50 mL of sterile production media containing the best nitrogen source was prepared in different conical flasks and supplemented with 2% of different carbon sources (glucose, lactose, sucrose, fructose, maltose, and starch). All flasks were inoculated with 5% of inoculum and incubated for 72 h at 35 °C after sterilization. The PHB production was estimated as described previously.
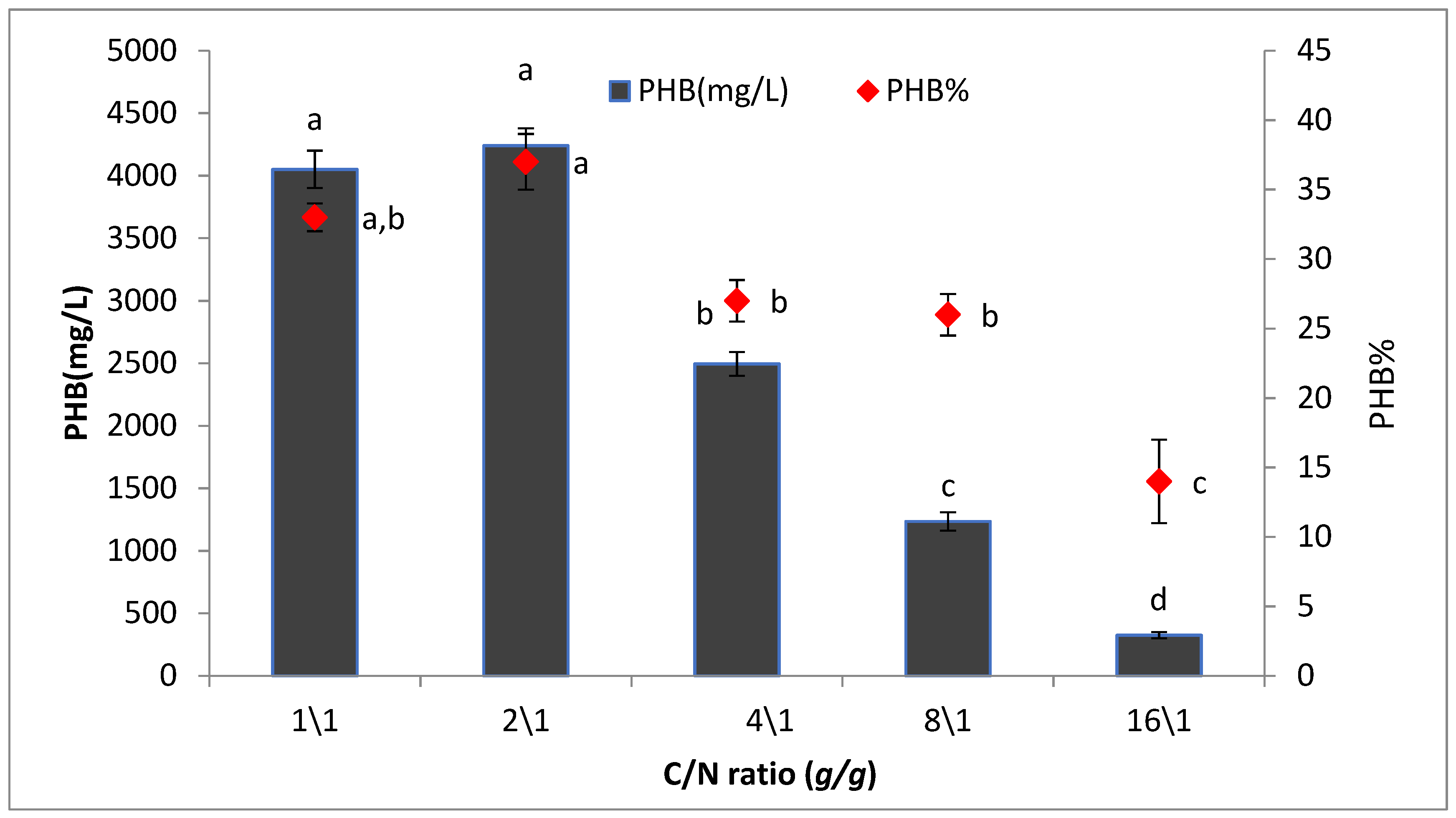
2.8.6. Effect of Different Carbon to Nitrogen Ratios on PHB Production (C/N Ratio)
To detect the best C: N ratios for PHB production, the strain was inoculated in production media supplemented with different ratios of concentrations of the best C and N sources (C/N ratio as 1:1, 2:1, 4:1, 8:1, 10:1, and 20:1). The culture was incubated at 35 °C for 72 h. After incubation, PHB content was quantified according to the yields, based on which the most favorable C/N ratio was determined.
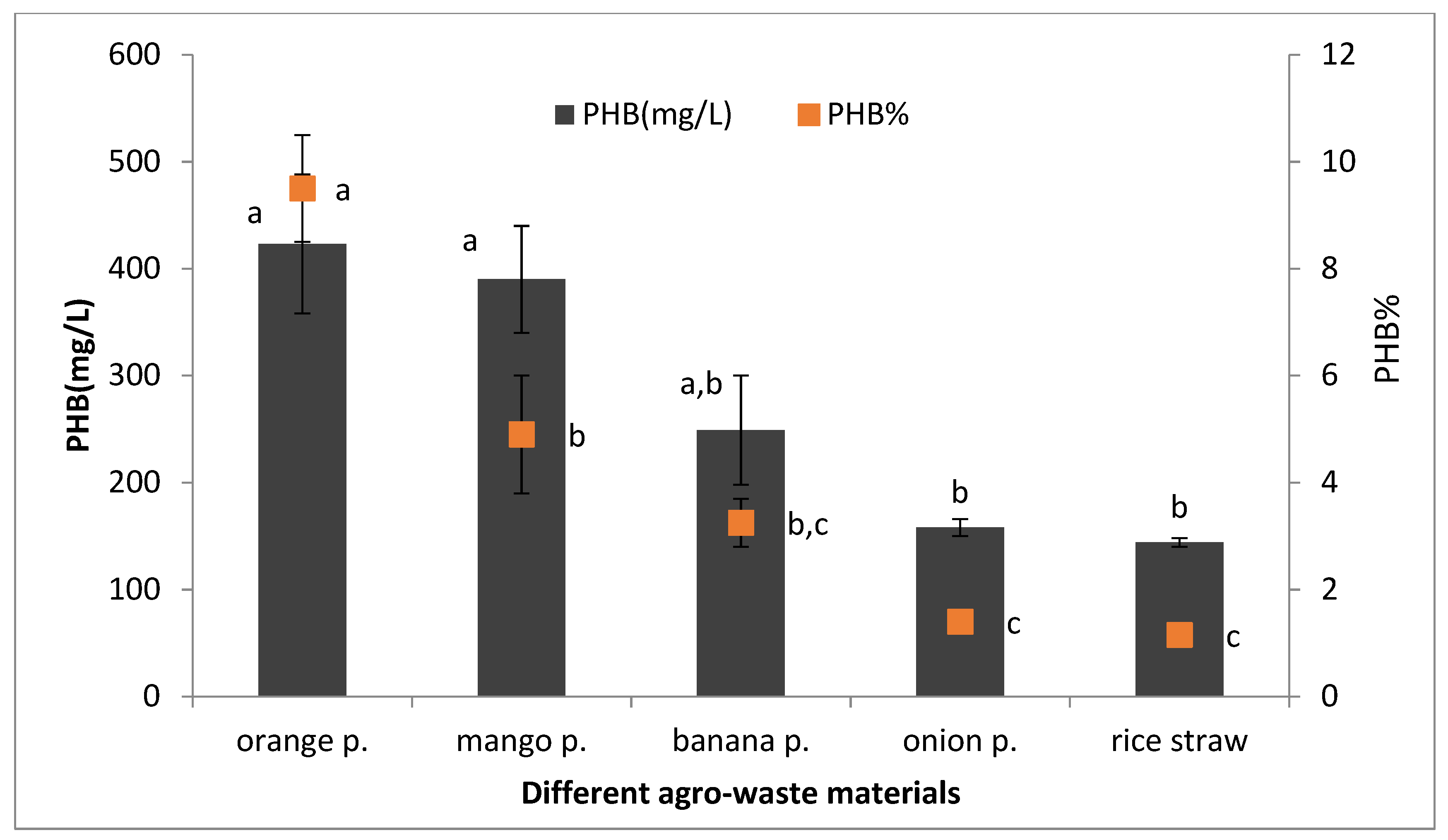
2.9. Development of Low-Cost Production Media Such as Fruit Peel Waste
Agricultural waste materials such as orange peel, mango peel, banana peel, onion peels, and rice straw were collected and dried for 5 to 7 days. Then, they were powdered and used for the preparation of the extract. Acid pre-treatment for agro-wastes was carried out by hydrolyzing of wastes using (0.5–5%) sulphuric acid and autoclaving wastes at 121 °C for 30 min. The extract was filtered, and the supernatant was neutralized by sodium hydroxide. The hydrolysates extract at 4% concentration, supplemented with the media component except for carbon source, were used as production media for PHB production by selected bacteria [
12]. The total carbohydrate content of waste material was estimated using phenol sulphuric acid method; 0.2 mL of extract was mixed with 1.8 mL distilled water in boiling tubes. A combination of 1 mL of 5% phenol and 5 mL of 96% sulphuric acid were added into all tubes one by one and shook well so that the phenol and sulphuric acid were mixed thoroughly with working standard. After 10 min, all the tubes were placed in water bath at 25–30 °C for 15 min. Blank was set with 1 mL of distilled water, and absorbance was measured at 490 nm using a double beam spectrophotometer (Thermo Scientific, Evolution 160, Waltham, MA, USA) [
19]. In addition, protein content for each type of waste was estimated according to the Lowry method using the following reagents: Reagent A, 2 g of Na
2CO
3 dissolved in 100 mL of 0.1 N sodium hydroxide; Reagent B, 0.5 g of CuSO
4. H
2O was dissolved in 1% sodium potassium tartrate. Alkaline reagent solution was freshly prepared by mixing 50 mL of reagent A with 1 mL of reagent B. Five milliliters of the alkaline reagent solution was added to 1 mL of the extract in a clean tube. Both were mixed thoroughly and allowed to stand at room temperature for at least 10 min. Then, 0.5 mL of the diluted folin reagent 1:1(
v/
v) was added to the above mixture and immediately mixed. After 30 min, absorbance against a blank (devoid of proteins) was measured at 570 nm with a double beam spectrophotometer (Thermo Scientific, Evolution 160, Waltham, MA, USA). A calibration curve was constructed using egg albumin [
20].
2.10. Statistical Analysis
Statistical analysis of the data was conducted using ANOVA one-way test (analysis of variance) by SPSS program version 21 (IBM Corp., Armonk, NY, USA), and Duncan values were determined at 0.05 levels.
2.11. GenBank Accession Number
The nucleotide sequence of 16S rRNA gene sequences of isolated strain AS-02 reported in this study has been deposited in the DDBJ, EMBL, and GenBank nucleotide sequence databases under the name Bacillus wiedmanni and the accession number OK576278.


 ,
, 














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}