
Potential of Acidithiobacillus ferrooxidans to Grow on and Bioleach Metals from Mars and Lunar Regolith Simulants under Simulated Microgravity Conditions

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Media and Growth Conditions
2.2. Composition of Lunar and Martian Regolith Simulants
2.3. Microgravity Simulation Experiment
2.4. Physicochemical Analysis
2.5. Cell Counting
2.6. Scanning Electron Microscopy (SEM)
2.7. Scanning Transmission Electron Microscopy (STEM)
2.8. Statistical Analysis
3. Results
3.1. Cell Growth
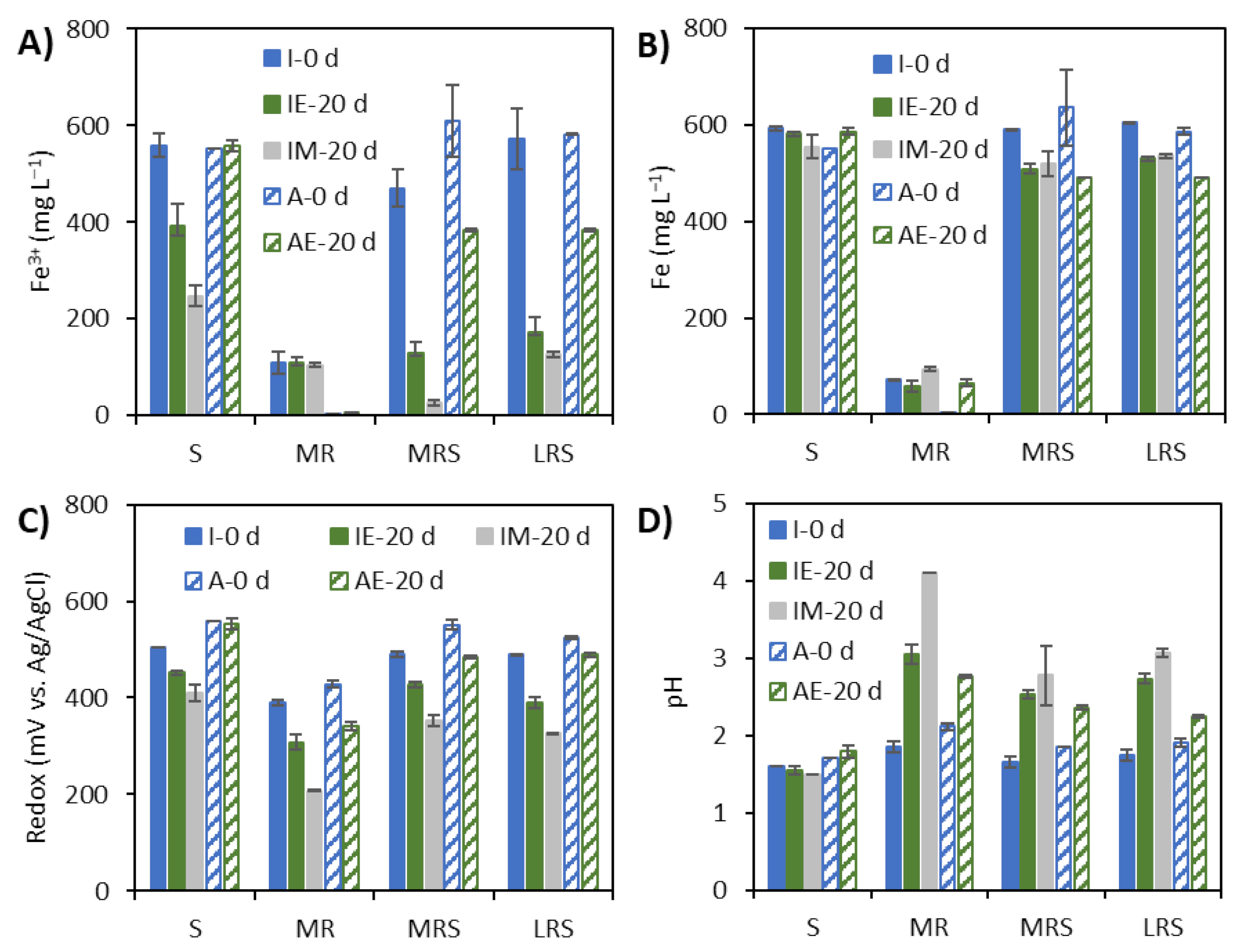
3.2. Changes in Ferric and Total Soluble Iron Concentration, Redox Potential and pH
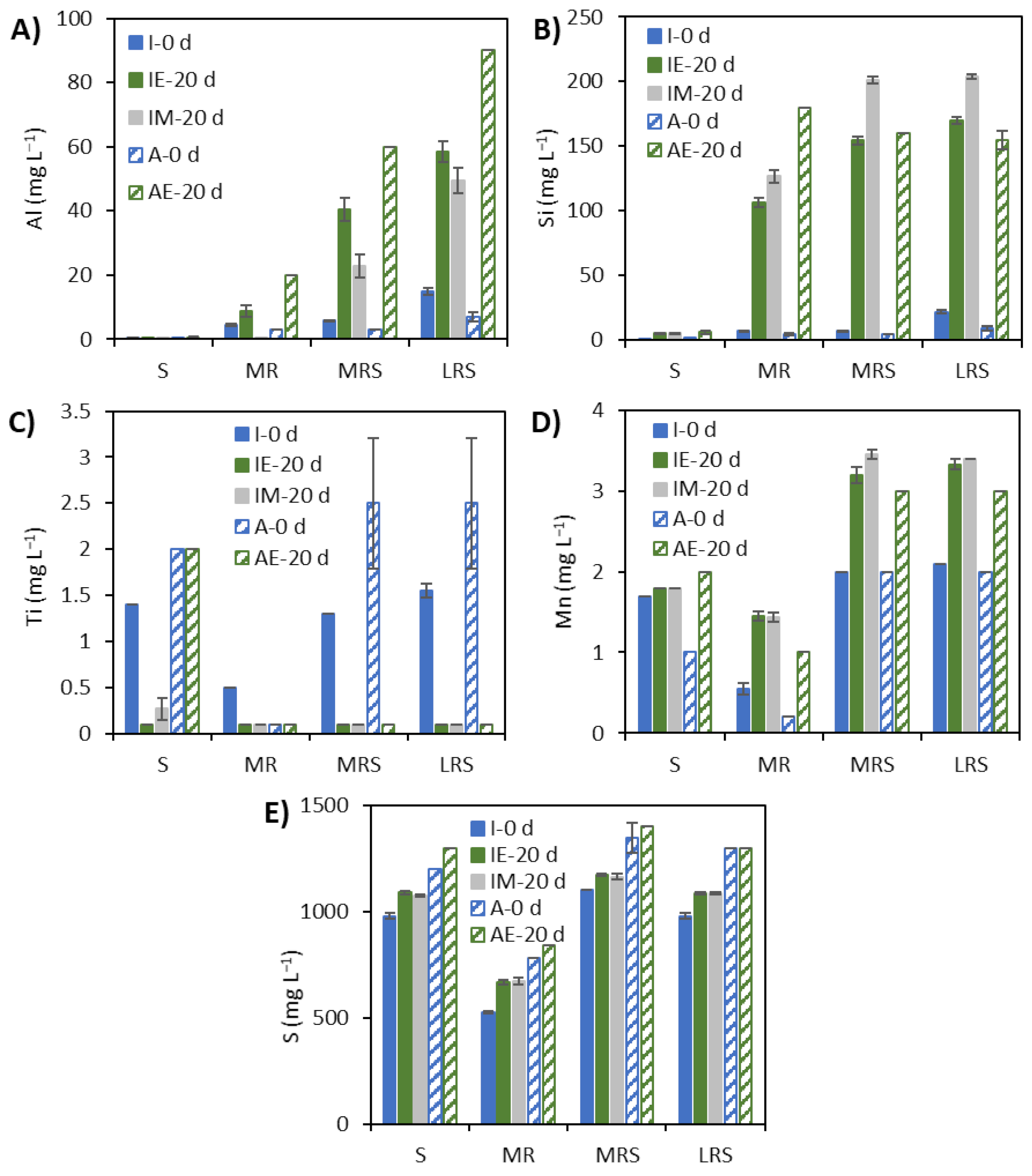
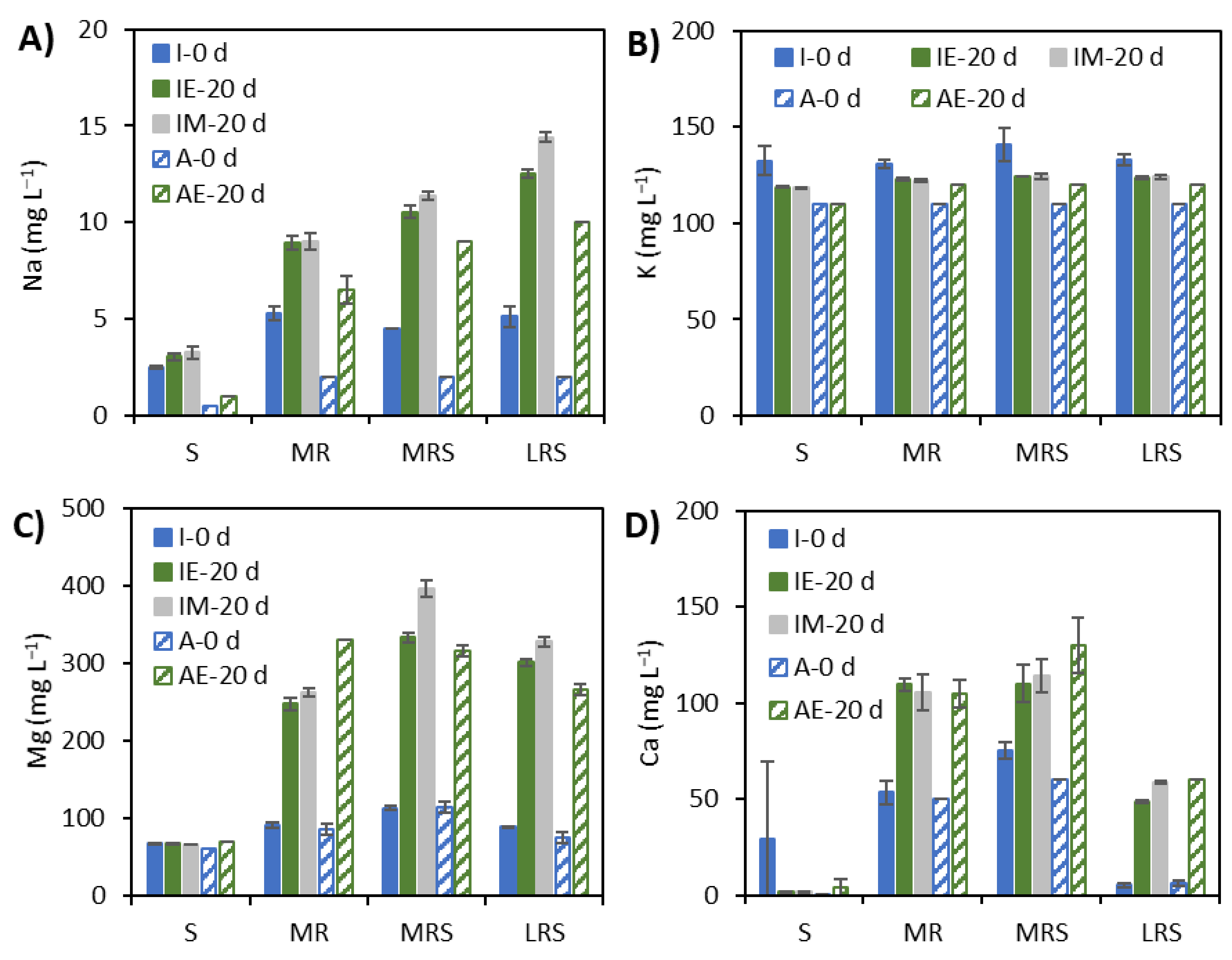
3.3. Solubilisation Elements under Earth Gravity and Simulated Microgravity Conditions
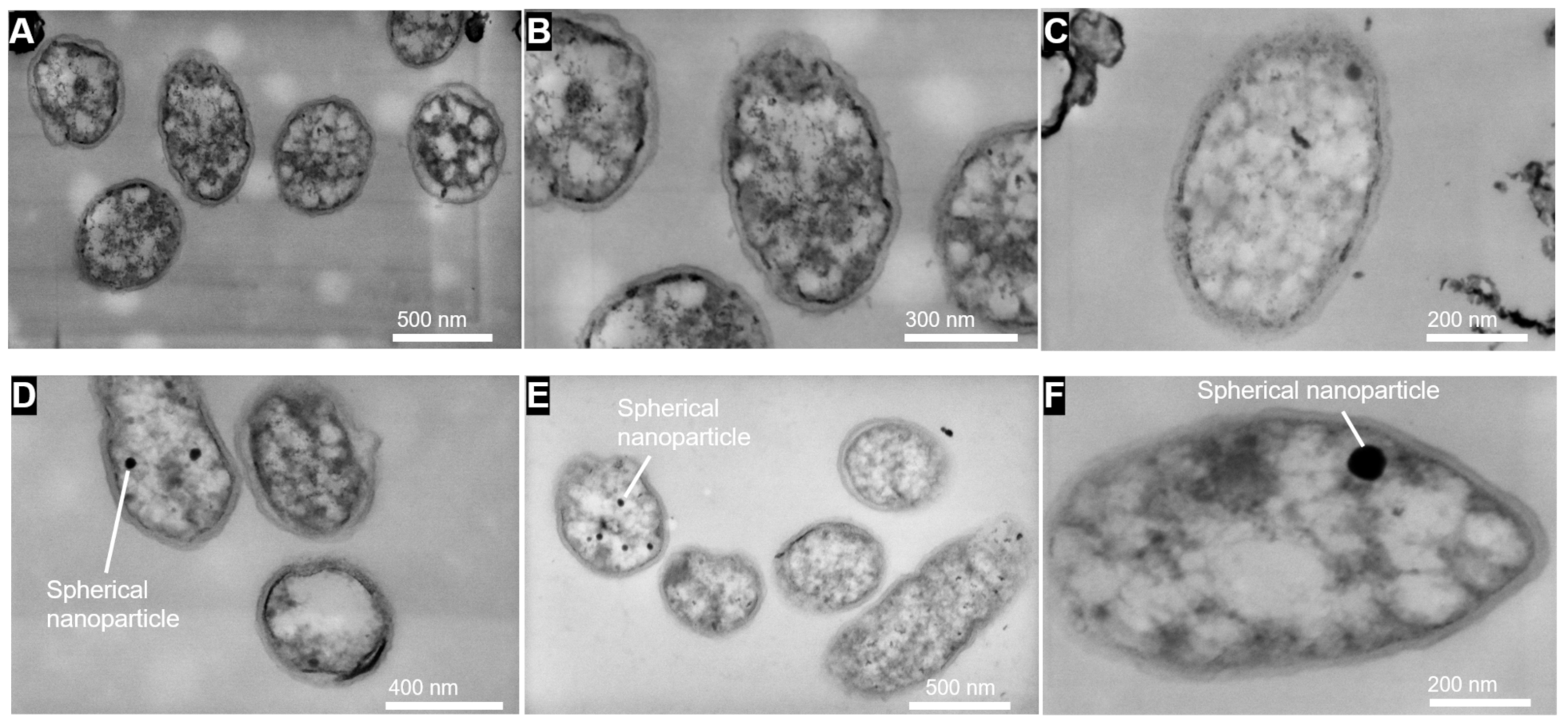
3.4. SEM and STEM Observation of Cell Morphology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Das, T.; Ayyappan, S.; Chaudhury, G.R. Factors affecting bioleaching kinetics of sulfide ores using acidophilic microorganisms. Biometals 1999, 12, 1–10. [Google Scholar] [CrossRef]
- Hong, Y.; Valix, M. Bioleaching of electronic waste using acidophilic sulfur oxidising bacteria. J. Clean. Prod. 2014, 65, 465–472. [Google Scholar] [CrossRef]
- Kaksonen, A.H.; Boxall, N.J.; Usher, K.M.; Ucar, D.; Sahinkaya, E. Biosolubilisation of metals and metalloids. In Sustainable Heavy Metal Remediation; Volume 1: Principles and Processes; Rene, E.R., Sahinkaya, E., Lewis, A., Lens, P.N.L., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 233–283. [Google Scholar]
- Menezes, A.A.; Montague, M.G.; Cumbers, J.; Hogan, J.A.; Arkin, A.P. Grand challenges in space synthetic biology. J. R. Soc. Interface 2015, 12, 20150803. [Google Scholar] [CrossRef]
- Dalton, B.; Roberto, F. Lunar Regolith Biomining: Workshop Report; NASA: Moffet Field, CA, USA, 2008.
- Klingelhöfer, G.; Morris, R.V.; Bernhardt, B.; Schröder, C.; Rodionov, D.S.; De Souza, P.A.; Yen, A.; Gellert, R.; Evlanov, E.N.; Zubkov, B.; et al. Jarosite and hematite at Meridiani Planum from opportunity’s Mössbauer spectrometer. Science 2004, 306, 1740–1745. [Google Scholar] [CrossRef]
- Squyres, S.W.; Grotzinger, J.P.; Arvidson, R.E.; Bell, J.F.; Calvin, W.; Christensen, P.R.; Clark, B.C.; Crisp, J.A.; Farrand, W.H.; Herkenhoff, K.E.; et al. In situ evidence for an ancient aqueous environment at Meridiani Planum, Mars. Science 2004, 306, 1709–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauermeister, A.; Rettberg, P.; Flemming, H.C. Growth of the acidophilic iron-sulfur bacterium Acidithiobacillus ferrooxidans under Mars-like geochemical conditions. Planet. Space Sci. 2014, 98, 205–215. [Google Scholar] [CrossRef]
- Gronstal, A.; Pearson, V.; Kappler, A.; Dooris, C.; Anand, M.; Poitrasson, F.; Kee, T.P.; Cockell, C.S. Laboratory experiments on the weathering of iron meteorites and carbonaceous chondrites by iron-oxidizing bacteria. Meteorit. Planet. Sci. 2009, 44, 233–247. [Google Scholar] [CrossRef]
- Gómez Felipe, F.; Mateo-Martí, E.; Prieto-Ballesteros, O.; Martín-Gago, J.; Amils, R. Protection of chemolithoautotrophic bacteria exposed to simulated Mars environmental conditions. Icarus 2010, 209, 482–487. [Google Scholar] [CrossRef] [Green Version]
- Loudon, C.M.; Nicholson, N.; Finster, K.; Leys, N.; Byloos, B.; Van Houdt, R.; Rettberg, P.; Moeller, R.; Fuchs, F.M.; Demets, R.; et al. BioRock: New experiments and hardware to investigate microbe-mineral interactions in space. Int. J. Astrobiol. 2018, 17, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Volger, R.; Pettersson, G.M.; Brouns, S.J.J.; Rothschild, L.J.; Cowley, A.; Lehner, B.A.E. Mining moon & mars with microbes: Biological approaches to extract iron from Lunar and Martian regolith. Planet. Space Sci. 2020, 184, 104850. [Google Scholar] [CrossRef]
- Nickerson, C.A.; Ott, C.M.; Wilson, J.W.; Ramamurthy, R.; Pierson, D.L. Microbial responses to microgravity and other low-shear environments. Microbiol. Mol. Biol. Rev. 2004, 68, 345–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenzweig, J.A.; Abogunde, O.; Thomas, K.; Lawal, A.; Nguyen, Y.U.; Sodipe, A.; Jejelowo, O. Spaceflight and modeled microgravity effects on microbial growth and virulence. Appl. Microbiol. Biotechnol. 2010, 85, 885–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarrete, J.U.; Cappelle, I.J.; Schnittker, K.; Borrok, D.M. Bioleaching of ilmenite and basalt in the presence of iron-oxidizing and iron-scavenging bacteria. Int. J. Astrobiol. 2013, 12, 123–134. [Google Scholar] [CrossRef]
- Kölbl, D.; Pignitter, M.; Somoza, V.; Schimak, M.P.; Strbak, O.; Blazevic, A.; Milojevic, T. Exploring fingerprints of the extreme thermoacidophile Metallosphaera sedula grown on synthetic martian regolith materials as the sole energy sources. Front. Microbiol. 2017, 8, 1918. [Google Scholar] [CrossRef] [Green Version]
- Dougherty, M. Results of the Micro-12 Flight Experiment: Effects of Microgravity on Shewanella Oneidensis MR-1; NASA: Moffet Field, CA, USA, 2019.
- Klaus, D.M. Clinostat and bioreactors. Gravit. Space Res. 2007, 14, 55–64. [Google Scholar]
- Zea, L.; Estante, F.; Rosengren, A.; Stodieck, L.; Klaus, L. Determining the appropriate clinostat rotational speed. In Proceedings of the 34th Annual Meeting of the American Society for Gravitational and Space Research (ASGSR), Washington, DC, USA, 31 October–3 November 2018. [Google Scholar]
- Govender, E.; Harrison, S.T.L.; Bryan, C.G. Modification of the ferric chloride assay for the spectrophotometric determination of ferric and total iron in acidic solutions containing high concentrations of copper. Miner. Eng. 2012, 35, 46–48. [Google Scholar] [CrossRef]
- Eckert, E.; Schaechterl, M. Bacterial growth under conditions of limited nutrition. Ann. N. Y. Acad. Sci. 1962, 102, 549–563. [Google Scholar] [CrossRef]
- Yan, L.; Zhang, S.; Chen, P.; Wang, W.; Wang, Y.; Li, H. Magnetic properties of Acidithiobacillus ferrooxidans. Mater. Sci. Eng. C 2013, 33, 4026–4031. [Google Scholar] [CrossRef]
- Sun, J.; Li, Y.; Liang, X.J.; Wang, P.C. Bacterial Magnetosome: A novel biogenetic magnetic targeted drug carrier with potential multifunction. J. Nanomater. 2011, 2011, 469031–469043. [Google Scholar] [CrossRef]
- Huang, B.; Li, D.G.; Huang, Y.; Liu, C.T. Effects of spaceflight and simulated microgravity on microbial growth and secondary metabolism. Mil. Med. Res. 2018, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Benoit, M.; Klaus, D.M. Microgravity, bacteria, and the influence of motility. Adv. Space Res. 2007, 39, 1225–1232. [Google Scholar] [CrossRef]
- Baker, P.; Leff, L.G. The effect of simulated microgravity on bacteria from the Mir space station. Microgravity Sci. Technol. 2004, 15, 35–41. [Google Scholar] [CrossRef]
- Li, Y.Q.; Wan, D.S.; Huang, S.S.; Leng, F.F.; Yan, L.; Ni, Y.Q.; Li, H.Y. Type IV pili of Acidithiobacillus ferrooxidans are necessary for sliding, twitching motility and adherence. Curr. Microbiol. 2010, 60, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milojevic, T.; Weckwerth, W. Molecular mechanisms of microbial survivability in outer space: A systems biology approach. Front. Microbiol. 2020, 11, 923. [Google Scholar] [CrossRef]
- Naser, M.Z. Extraterrestrial construction materials. Prog. Mater. Sci. 2019, 105, 100577. [Google Scholar] [CrossRef]
- Cockell, C.S.; Santomartino, R.; Finster, K.; Waajen, A.C.; Eades, L.J.; Moeller, R.; Rettberg, P.; Fuchs, F.M.; Van Houdt, R.; Leys, N.; et al. Space station biomining experiment demonstrates rare earth element extraction in microgravity and Mars gravity. Nat. Commun. 2020, 11, 5523. [Google Scholar] [CrossRef] [PubMed]
- Cockell, C.S.; Santomartino, R.; Finster, K.; Waajen, A.C.; Nicholson, N.; Loudon, C.M.; Eades, L.J.; Moeller, R.; Rettberg, P.; Fuchs, F.M.; et al. ’ Everroad, R.C.; Demets, R. Microbially enhanced vanadium mining and bioremediation under micro and mars gravity on the International Space Station. Front. Microbiol. 2021, 12, 641387. [Google Scholar] [CrossRef]
- Shively, J.M.; Cannon, G.C.; Heinhorst, S.; Fuerst, J.A.; Bryant, D.A.; Gantt, E.; Maupin-Furlow, J.A.; Schüler, D.; Pfeifer, F.; Docampo, R.; et al. Intracellular structures of prokaryotes: Inclusions, compartments and assemblages. In Encyclopedia of Microbiology, 3rd ed.; Schaechter, M., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2009; pp. 404–424. [Google Scholar]
- Olszweska-Widdrat, A.; Schiro, G.; Reichel, W.E.; Faivre, D. Reducing conditions favor magnetosome production in Magnetospirillum magneticum AMB-1. Front. Microbiol. 2019, 10, 582. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yan, L.; Li, H.; Liu, H. Optimal conditions for growth and magnetosome formation of Acidithiobacillus ferrooxidans. Afr. J. Microbiol. Res. 2012, 6, 6142–6151. [Google Scholar] [CrossRef]
- Urban, J.E. Adverse effects of microgravity on the magnetotactic bacterium Magnetospirillum magnetotacticum. Acta Astronaut. 2000, 47, 775–780. [Google Scholar] [CrossRef]
- Kuzajewska, D.; Wszolek, A.; Zwierello, W.; Kirczuk, L.; Maruszweska, A. Magnetotactic bacteria and magnetosomes as smart drug delivery systems: A new weapon on the battlefield with cancer? Biology 2020, 9, 102. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Major Chemical Composition | Mars Global Simulant (MGS-1)(%-wt) | Lunar Mare Simulant (LMS-1) (%-wt) |
|---|---|---|
| SiO2 | 45.57 | 42.81 |
| TiO2 | 0.30 | 4.62 |
| Al2O3 | 9.43 | 14.13 |
| Cr2O3 | 0.12 | 0.21 |
| FeO T | 16.85 | 7.87 |
| MgO | 16.50 | 18.89 |
| MnO | 0.10 | 0.15 |
| CaO | 4.03 | 5.94 |
| Na2O | 3.66 | 4.92 |
| K2O | 0.43 | 0.57 |
| P2O5 | 0.37 | - |
| SO3 | 2.63 | 0.11 |
| Mineral Component | Mars Global Simulant (MGS-1)(%-wt) | Lunar Mare Simulant (LMS-1) (%-wt) |
|---|---|---|
| Pyroxene | 20.3 | 32.8 |
| Plagioclase | 27.1 | 19.8 |
| Olivine | 13.7 | 11.1 |
| Basalt | - | 7.5 |
| Basaltic glass | 22.9 | - |
| Ilmenite | - | 4.3 |
| Mg-sulfate | 4.0 | - |
| Ferrihydrite | 3.5 | - |
| Hydrated silica | 3.0 | - |
| Magnetite | 1.9 | - |
| Anhydrite | 1.7 | - |
| Fe-carbonate | 1.4 | - |
| Hematite | 0.5 | - |
| Treatment | Regolith Simulant (1% w/v) | Electron Acceptor/Donor 1 | Culture (10% v/v) | Gravity |
|---|---|---|---|---|
| S-IM | - | Fe3+ + H2/CO2 | A. ferrooxidans | Simulated µg |
| MR-IM | Martian | H2/CO2 | A. ferrooxidans | Simulated µg |
| MRS-IM | Martian | Fe3+ + H2/CO2 | A. ferrooxidans | Simulated µg |
| LRS-IM | Lunar | Fe3+ + H2/CO2 | A. ferrooxidans | Simulated µg |
| S-IE | - | Fe3+ + H2/CO2 | A. ferrooxidans | Earth Gravity |
| MR-IE | Martian | H2/CO2 | A. ferrooxidans | Earth Gravity |
| MRS-IE | Martian | Fe3+ + H2/CO2 | A. ferrooxidans | Earth Gravity |
| LRS-IE | Lunar | Fe3+ + H2/CO2 | A. ferrooxidans | Earth Gravity |
| S-AE | - | Fe3+ + H2/CO2 | - | Earth Gravity |
| MR-AE | Martian | H2/CO2 | - | Earth Gravity |
| MRS-AE | Martian | Fe3+ + H2/CO2 | - | Earth Gravity |
| LRS-AE | Lunar | Fe3+ + H2/CO2 | - | Earth Gravity |
| Ferric Iron Consumption Rate (mg L−1 d−1) | ||
|---|---|---|
| Cultures | Earth Gravity | Simulated Microgravity |
| S-I | 8.3 ± 2.2 | 15.6 ± 1.1 |
| MR-I | NA | NA |
| MRS-I | 17.1 ± 1.1 | 22.2 ± 0.2 |
| LRS-I | 20.0 ± 1.5 | 22.3 ± 0.3 |
| S-A | 0 | ND |
| MR-A | NA | ND |
| MRS-A | 11.3 ± 3.8 | ND |
| LRS-A | 9.9 ± 0.3 | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaksonen, A.H.; Deng, X.; Morris, C.; Khaleque, H.N.; Zea, L.; Gumulya, Y. Potential of Acidithiobacillus ferrooxidans to Grow on and Bioleach Metals from Mars and Lunar Regolith Simulants under Simulated Microgravity Conditions. Microorganisms 2021, 9, 2416. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122416
Kaksonen AH, Deng X, Morris C, Khaleque HN, Zea L, Gumulya Y. Potential of Acidithiobacillus ferrooxidans to Grow on and Bioleach Metals from Mars and Lunar Regolith Simulants under Simulated Microgravity Conditions. Microorganisms. 2021; 9(12):2416. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122416
Chicago/Turabian StyleKaksonen, Anna H., Xiao Deng, Christina Morris, Himel Nahreen Khaleque, Luis Zea, and Yosephine Gumulya. 2021. "Potential of Acidithiobacillus ferrooxidans to Grow on and Bioleach Metals from Mars and Lunar Regolith Simulants under Simulated Microgravity Conditions" Microorganisms 9, no. 12: 2416. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122416