
Bacterial Interactions with Aspergillus fumigatus in the Immunocompromised Lung

Department of Biology, Maynooth University, Manooth W23VP22, Kildare, Ireland
*
Author to whom correspondence should be addressed.
Microorganisms 2021, 9(2), 435; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020435
Submission received: 1 February 2021
/
Revised: 18 February 2021
/
Accepted: 18 February 2021
/
Published: 19 February 2021
(This article belongs to the Special Issue Aspergillus and Health 1.0)
Abstract
:The immunocompromised airways are susceptible to infections caused by a range of pathogens which increases the opportunity for polymicrobial interactions to occur. Pseudomonas aeruginosa and Staphylococcus aureus are the predominant causes of pulmonary infection for individuals with respiratory disorders such as cystic fibrosis (CF). The spore-forming fungus Aspergillus fumigatus, is most frequently isolated with P. aeruginosa, and co-infection results in poor outcomes for patients. It is therefore clinically important to understand how these pathogens interact with each other and how such interactions may contribute to disease progression so that appropriate therapeutic strategies may be developed. Despite its persistence in the airways throughout the life of a patient, A. fumigatus rarely becomes the dominant pathogen. In vitro interaction studies have revealed remarkable insights into the molecular mechanisms that drive agonistic and antagonistic interactions that occur between A. fumigatus and pulmonary bacterial pathogens such as P. aeruginosa. Crucially, these studies demonstrate that although bacteria may predominate in a competitive environment, A. fumigatus has the capacity to persist and contribute to disease.
{kind=link}
{kind=link}
{kind=link}
1. Introduction
By definition, polymicrobial communities are a collection of microbial species that co-exist in a particular habitat [1]. Arising from co-habitation, interspecies interactions occur, and these interactions shape the landscape of the environment in which the microbial communities reside [2]. When that environment exists in humans, such interactions can influence the health status of an individual. In the context of infectious disease, polymicrobial interactions can be synergistic, whereby the combined effect of multiple microbial species is worse than that where individual species act alone [2]. On the other hand, antagonistic interactions arise due to competition, and occur when one species suppresses the other [3]. The mechanisms employed by microbes as a consequence of these interactions is often detrimental to host health. Inter-species interactions may influence microbial pathogenesis by altering microbial virulence factors and disease progression [1]. Thus, an understanding of how polymicrobial communities, interact with each other and with the host is important when deciding appropriate therapeutic strategies [1,4].
Individuals with chronic pulmonary disease such as cystic fibrosis (CF) or chronic obstructive pulmonary disease (COPD) are susceptible to respiratory infections caused by a multitude of microbial species; the filamentous fungus Aspergillus fumigatus is the dominant cause of fungal infections in the immunocompromised airways, while the bacterium Pseudomonas aeruginosa is the chief cause of bacterial infections [5,6]. Importantly, these pathogens do not act in isolation, rather their development in the airways is governed by their environment and mutual interactions. Co-infection of the lungs by these pathogens is associated with poor prognostic outcomes for the patient, thus, understanding how these interactions affect disease progression can be key to identifying enhanced control mechanisms.
By their very nature, polymicrobial interactions are difficult to dissect and the results are largely dependent upon the in vitro model systems used to analyze these interactions. Nonetheless, the findings arising from such studies increase our understanding of these relationships and may provide a pathway to design new therapeutic targets. This review will highlight some of the studies concerning the interactions that occur between A. fumigatus and other common pulmonary pathogens, with a focus on P. aeruginosa, and will consider how the findings may influence the pathogenesis of pulmonary diseases caused by the co-existence of these pathogens in the airways.
2. Aspergillus fumigatus
A. fumigatus is an opportunistic fungal pathogen and the most pathogenic member of its genus [7]. Although its natural ecological niche is the soil, A. fumigatus is ubiquitous, existing indoors and outdoors [8]. Because of this, inhalation of conidia is a daily occurrence. A. fumigatus is a versatile microorganism that is equipped to survive and propagate in a variety of environments [9]. The fungus possesses a number of features that make it an excellent human pathogen, including the ability to grow at high temperatures and varying pH. A. fumigatus can sustain growth above 42 °C, which in the context of human infection is beneficial for maintaining infection during a febrile state [10]. Additionally, it can adapt to the changing pH of the mammalian host by activating a set of pH-responsive genes regulated by the PacC transcription factor [11,12].
Like many pathogens, A. fumigatus can form a biofilm which enables persistence in the host. A. fumigatus biofilms are formed when conidia and hyphae become embedded in a self-made hydrophobic extracellular matrix composed of glucans, galactomannans, monosaccharides, hydrophobins, and major antigen proteins [13,14]. The emergence of hyphae within biofilm coincides with the production of secondary metabolites (e.g., gliotoxin, fumagillin), antigenic surface molecules (β(1,3) glucans), and antigens (e.g., aspergillopepsin), which the host responds to by inducing a proinflammatory response. Biofilm formation is dependent upon fungal cell density [15], thus, the ability to reach this density threshold may play a factor in the ability of A. fumigatus to establish biofilms in the host. Biofilms enable fungal persistence in the pulmonary cavity by providing protection against cells of the immune system and antifungal drugs. Furthermore, in vitro studies have shown that while hyphae contained in biofilms may be inhibited by competing microbes, they are difficult to kill [16,17,18].
The physical size and hydrophobic nature enable A. fumigatus conidia to enter the respiratory tract through inhalation, bypass mucociliary clearance, and reach the alveoli. A. fumigatus conidia may evade initial host-cell recognition by masking β(1,3)-glucan residues on the condial cell wall with a thin proteinaceous hydrophobic layer called RodA hydrophobin [19]. As conidia germinate, the RodA layer is shed and β(1,3)-glucan residues are revealed, allowing for recognition by cells of the innate immune system [20]. The shedding of RodA also reveals dihydroxynapthalene (DHN)-melanin, a secondary metabolite found in the conidial cell wall. In the environment, DHN-melanin confers resistance against desiccation and damage from UV radiation, and in the host it plays an important role in virulence by scavenging reactive oxygen species (ROS) and protecting conidia against phagocytosis by host cells [21,22,23].
Where the immune system is compromised, conidia germinate and hyphae may form [20]. This may lead to the manifestation of a disease called aspergillosis, the severity of which is determined by the immune status of the host. There are three forms of aspergillosis; allergic aspergillosis, the most common form of which is known as allergic bronchopulmonary aspergillosis (ABPA) is characterized by the induction of an immune response triggered by the secretion of toxins and allergens from the developing fungus. Saprophytic aspergillosis is characterized by the development of aspergilloma (fungal ball) in chronic lung cavities of the pulmonary tissue, such as those caused by tuberculosis [24]. Invasive aspergillosis (IA) is the most devastating form of aspergillosis and is characterized by the dissemination of fungal hyphae throughout the tissues of the affected area. This occurs in the lungs in more than 90% of cases and is called invasive pulmonary aspergillosis (IPA) [25]. IA targets severely immunocompromised individuals including individuals with neutropenia, organ transplant recipients, and chemotherapy patients [7].
A. fumigatus is the causative agent of allergic bronchopulmonary aspergillosis (ABPA). It is estimated that 1–2% of asthma patients and 1–15% of CF patients are affected by ABPA [26]. Clinical manifestations of ABPA include wheezing and bronchospasms and for individuals with CF, decline in lung function may occur [27]. For non-CF patients, ABPA diagnostic criteria include asthma, elevated serum levels of Aspergillus-specific IgG antibodies, elevated serum levels of IgE and eosinophilia [28,29]. Several of the diagnostic criteria for ABPA are common manifestations of CF, for example, elevated IgG and IgE anti-A. fumigatus antibodies are not uncommon in CF serum due to sensitization to A. fumigatus in CF [30]. For this reason diagnosis of ABPA in a CF patient may present certain challenges [29]. Nonetheless, in the context of ABPA diagnosis, A. fumigatus-specific IgE levels are recognized as the most useful diagnostic tool [30,31].
ABPA is described as a hypersensitivity lung disease in response to bronchial colonization by A. fumigatus [32]. It occurs when conidia deposited in the airways begin to germinate and release metabolites such as gliotoxin, fumagillin, and allergens such as Asp f family of allergens [33,34]. These toxins disturb the epithelial barrier and impede mucociliary clearance [35,36]. An influx of pulmonary macrophages and neutrophils mediate a proinflammatory cytokine cascade that promote a Th2-type adaptive immune response involving the release of IL-4, IL-5, IL-9, and IL-13 [37]. IL-4 induces IgE production, which binds to, and sensitizes granulocytes including basophils and mast cells. IL-5 and IL-9 recruit eosinophils and mast cells to the infection site and IL-13 induces mucus hypersecretion, airway fibrosis, and eotaxin production, thus contributing to the eosinophilic inflammatory response [38,39]. These factors contribute to the chronic inflammation that feature heavily in the CF airways.
In the absence of ABPA, the role of A. fumigatus in CF is becoming better understood and more appreciated. Until recently, young children with CF were thought to be less affected by A. fumigatus than older patients, with a prevalence rate of 6%–25%, compared with up to 57% in adults [40,41]. Traditional culture methods, such as plate assays, likely underestimate the actual prevalence of A. fumigatus among this group of patients [42]. The inclusion of molecular methods (qPCR) as a diagnostic tool for A. fumigatus infections provide a more accurate scenario and indicate that A. fumigatus is more prevalent in juveniles than previously reported [42]. Recent longitudinal studies have provided evidence to support this and A. fumigatus infections in children are now recognized as a major contributing factor in lung function decline in this cohort of patients, affecting up to 68% of patients [42,43,44,45]. This is, in part, associated with aggressive antibiotic therapies targeting bacterial pathogens, which thereby provide fungal pathogens with an opportunity to colonize [44]. A. fumigatus infection during early childhood is correlated with structural damage to the lung and decline in lung function, and while co-infection with other pathogens exacerbates disease prognosis the long term effects of early exposure to A. fumigatus remain to be explored [43,45,46,47].
3. Pseudomonas aeruginosa
P. aeruginosa is a Gram-negative, rod-shaped bacterium that is ubiquitous in nature, particularly in aquatic and soil environments. Its ubiquity is due to the ability of P. aeruginosa to survive in environmental niches that are intolerable to other microorganisms and its nutritional versatility. The genome of P. aeruginosa is large (~6.3 kbp) [48], and approximately 8–10% of these genes are predicted to be regulators of gene expression [49]. This confers P. aeruginosa with an incredible capacity to adapt rapidly to environmental changes such as nutritional availability [49]. Additionally, P. aeruginosa possess several efflux pumps which can expel toxic compounds, such as antibiotics, from the cell faster than they can accumulate [49,50]. A classic feature of chronic infection caused by P. aeruginosa is the increased exopolysaccharide production and the emergence of biofilms. Biofilms confer a layer of protection against phagocytic cells such as neutrophils, and antibiotics. Although neutrophils migrate to biofilms, they become immobilized and surrounded by bacteria that escape from biofilms. Neutrophil degranulation is compromised as a result and oxygen consumption by both neutrophils and the biofilm is increased, thereby reducing oxygen availability on the airways [51].
P. aeruginosa biofilm formation is dependent upon quorum sensing (QS), the mechanism by which bacteria communicate [52]. QS occurs in a cell density-dependent manner and is necessary for the biosynthesis of secondary metabolites such as pyocyanin and rhamnolipids, which induce neutrophil apoptosis and necrosis, respectively [53,54]. Biofilms are unable to form in the absence of iron and under iron-limiting conditions, QS regulates iron acquisition systems by inducing the production of siderophores such as pyoverdin [55,56].
The switch from non-mucoid to the over-producing alginate mucoid strain is probably the most pronounced phenotypic change that P. aeruginosa adopts as it establishes chronic infection [57]. Alginate plays an important role in the maturation and structural stability of P. aeruginosa biofilm and increases bacterial evasion of host immune cells and antibiotics [58]. Several loss-of-function mutations that occur during adaptation in the CF lung are characteristic of the establishment of chronic infection, including loss of motility, repression of type three secretion systems and downregulation of QS regulatory genes, such as lasR [59,60,61].
4. The Microbial Environment of the Immunocompromised Airways
The microbial environment of the CF airways is an evolving ecosystem, and from infancy, the lungs of CF patients are subject to colonization by a diverse range of microbial species from various genera including Streptococcus, Prevotella, Rothia, Veillonella, and Actinomyces [62,63]. The CF airways are characterized by an age-related succession of microbial species; in children under the age of 16, Staphylococcus aureus, Haemophilus influenzae, and Stenotrophomonas maltophilia predominate [64,65]. The reduction of infection with H. influenzae and S. aureus is strongly correlated with increased colonization by P. aeruginosa and Burkholderia spp., and a decline in lung function [62,65,66]. It is estimated that chronic infection with P. aeruginosa affects up to 80% of adults with CF.
Despite the diverse nature of the microbial community that exists in the CF airways, P. aeruginosa is consistently identified as the most common pathogen isolated from the lungs of patients after their first decade of life [64,67,68]. Reflected in this observation, in vitro and in vivo interaction studies involving P. aeruginosa demonstrate a greater capacity of P. aeruginosa, to outcompete other CF-associated species such as S. aureus and H. influenzae [69,70]. For this reason, P. aeruginosa has become the focus for many studies investigating the role of polymicrobial interactions in the context of CF.
Individuals that live with chronic non-cystic fibrosis-related respiratory diseases such as COPD and bronchiectasis are susceptible to infection by pathogens from multiple taxa including Pasteurellaceae, Streptococcaceae, and Pseudomonadaceae [71]. Microbial diversity is associated with clinical status and is reduced where acute exacerbations occur [71,72]. Two of the most common pathogens detected from the airways of individuals with bronchiectasis are P. aeruginosa and H. influenzae, however, due to antagonistic interactions that occur between these pathogens, when one is detected, the other is absent [71,72].
The prevalence of P. aeruginosa in adults with COPD is estimated to be between 4–15% and higher for individuals with severe COPD and bronchiectasis as part of the diagnosis [73,74]. In contrast, the frequency of chronic P. aeruginosa infection for individuals with bronchiectasis as the primary condition is between 9–31% [75]. Compared to infection by other pathogens, P. aeruginosa is associated with disease progression, recurrent pulmonary exacerbations, and poorer clinical outcomes, including a higher rate of mortality in patients with bronchiectasis [76,77].
The immunocompromised airways are susceptible to infection by a range of fungal pathogens and several of these, including Candida spp., Cryptococcus spp., and Scedosporium aurantiacum have been studied in the context of co-infection with P. aeruginosa [78,79,80]. In all cases, P. aeruginosa inhibits fungal growth and/or biofilm formation. Perhaps one of the most fascinating of the fungal-bacterial relationships associated with pulmonary infections, are those arising from the interactions between Aspergillus fumigatus and P. aeruginosa. Aspergillus fumigatus is the most common fungal pathogen isolated from the CF airways. It is detected from early childhood and is persistent in the airways throughout the life of a CF patient [42,64,66]. Longitudinal studies have shown that colonization with A. fumigatus is associated with an increased risk of P. aeruginosa colonization in CF, and disease prognosis is poor when both pathogens are present [47,81,82,83]. The prevalence of co-colonization with P. aeruginosa and A. fumigatus in the CF airways is estimated to be between 3.1 and 15.8%, although this occurrence may be higher [47,81,82].
Co-infection with P. aeruginosa and A. fumigatus have been detected in severe cases of COPD, and the presence of P. aeruginosa in the airways is considered a risk factor for A. fumigatus infection [84]. A. fumigatus is frequently isolated from the airways of individuals with COPD and bronchiectasis, and infection with A. fumigatus is a known risk factor for the onset of bronchiectasis in COPD [85,86,87,88]. ABPA is employed as a diagnostic feature of bronchiectasis and can inform treatment programs [89].
Due to the frequency at which these pathogens co-exist in the airways, the interactions that occur between A. fumigatus and P. aeruginosa are of immense clinical importance in the area of pulmonology. What drives pulmonary exacerbation when the two pathogens are present remains to be fully elucidated [90,91]. The increased number of studies that have begun to investigate A. fumigatus–P. aeruginosa interactions in the past decade is a reflection on the clinical importance of co-colonization with these pathogens. In general, the results of these studies show that P. aeruginosa outcompetes A. fumigatus, a finding supported by the predominance of the bacteria in the CF lung.
5. The Interactions Between A. fumigatus and P. aeruginosa
A. fumigatus persists in the CF airways throughout childhood and into adulthood, yet despite this, P. aeruginosa eventually predominates [42,47,81]. This suggests that interactions with pathogens such as A. fumigatus may influence the pathogenicity of P. aeruginosa by altering its virulence and the host environment to pave the way for chronic P. aeruginosa infection [92].
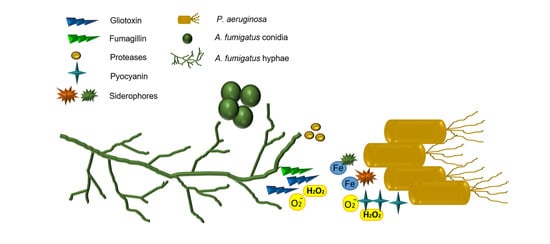
Analysis of the interactions between A. fumigatus and P. aeruginosa in vitro have revealed several antifungal mechanisms by which P. aeruginosa can outcompete A. fumigatus [16,93,94,95,96] (Figure 1). P. aeruginosa isolates taken from patients with cystic fibrosis have a greater antifungal capacity than non-cystic fibrosis isolates. Non-mucoid isolates are more inhibitory than mucoid isolates, which may explain why A. fumigatus is detected at higher levels in older cystic fibrosis patients where chronic (mucoid) P. aeruginosa infections are more common [91,97,98,99]. Many of these interaction studies have focused on the direct effects of P. aeruginosa on A. fumigatus-biofilm formation, on the effects of bacterial biosynthetic products (e.g., phenazines) on the fungal growth and development or, of fungal metabolites on P. aeruginosa [95,96,100,101,102].
P. aeruginosa secretes a range of compounds that inhibit A. fumigatus development and biofilm formation [16,94,95]. Phenazines (pyocyanin, phenazine-1-carboxamide, 1-HP and phenazine-1-carboxylic acid) are QS-regulated redox-active molecules that are important in bacterial respiration and energy production in oxygen-limiting environments such as the CF airways [103]. Phenazines are ROS producing compounds and in the host, changes in the redox balance caused by ROS result in host-cell damage and death [103]. The production of ROS by phenazines also has implications for A. fumigatus survival [95]. Phenazines can enter into swollen, but not resting, conidia and target the mitochondria, inducing ROS production [95]. The accumulation of ROS is thought to interfere with A. fumigatus growth and biofilms by inducing fungal apoptosis [95,96]. Exposure of A. fumigatus biofilms to culture supernatants from non-mucoid and mucoid P. aeruginosa CF isolates resulted in a greater increase of ROS in fungal biofilms exposed to the non-mucoid strain [96]. In the CF airways, mucoid strains are associated with the downregulation of QS-regulated molecules including phenazines [104]. This suggests that these antagonistic interactions may occur prior to the switch from non-mucoid to mucoid and the establishment of chronic infection in the CF lung.
The definitive role of phenazines as a fungicidal agent is uncertain, however, as phenazine-deficient mutants have also been shown to inhibit fungal growth, although the authors of this study acknowledge the possible anti-fungal role of an unknown molecule upregulated as a result of phenazine depletion [94]. The P. aeruginosa siderophores, pyoverdin and 1-hydroxyphenazine (1-HP), chelate iron in the environment, depriving A. fumigatus of a necessary nutrient, thereby suppressing fungal growth and biofilm formation [94,95]. Pyoverdin is thought to be the key component involved in outcompeting A. fumigatus, and mutants deficient in pyoverdin biosynthesis were unable to inhibit fungal growth [94].
Another class of P. aeruginosa QS-regulated molecules are dirhamnolipids. These biosurfactant molecules alter A. fumigatus cell-wall phenotype by interfering with the extracellular matrix, enabling enhanced bacterial binding to the fungus, increasing melanin production and inhibiting β 1,3-glucan synthase, causing the hyphal cell-wall to thicken, thereby suppressing fungal growth development [102]. In co-cultures, A. fumigatus stimulates P. aeruginosa elastase production, which inhibits the growth of fungus and is also cytotoxic to the alveolar epithelial cells, A549, in vitro [105].
These findings are of clinical relevance because although the arsenal of secondary metabolites secreted by P. aeruginosa in the presence of A. fumigatus may have anti-fungal properties, these bacterial compounds and the consequences arising from their interactions with A. fumigatus, may have negative implications for the host. For example, in vivo, melanin enables fungal evasion of phagocytic activity [106]. P. aeruginosa elastases can degrade host antimicrobial surfactant proteins SP-A and SP-D and disrupt tight junctions between epithelial cells [107,108,109]. Phenazines contribute to cytokine-mediated damage to host cells by induce proinflammatory cytokines and siderophores contribute to iron depletion in the host environment [110,111]
Despite the demonstrable ability of P. aeruginosa to subdue A. fumigatus growth (Figure 1) [16,17,94,95,102], several studies reported the capacity of A. fumigatus to compete with P. aeruginosa [100,112]. This supports the notion that A. fumigatus can persist in the CF airways, despite not being the dominant pathogen. For example, P. aeruginosa can inhibit the growth of A. fumigatus conidia, but not of preformed hyphae [17]. This may be attributed to the ability of hyphae, but not conidia to produce gliotoxin which has anti-Pseudomonas activity [100,113]. A. fumigatus produce hydroxamate-containing siderophores (ferricrocin, hydroxyferricrocin, fusarinine C, triacetylfusarinine C) in response to iron limitation. The production of these siderophores can mitigate the effect of P. aeruginosa pyoverdin and, in part, protect A. fumigatus biofilm, as shown in A. fumigatus siderophore-deficient mutants, which are more susceptible to the effects of pyoverdin than the wild-type [112].
A. fumigatus secretes a range of degradative enzymes that contribute to the ubiquity of the fungus in nature by supporting fungal growth on plant matter [114,115,116]. Many of these biological determinants also play a role in establishing disease in humans and are associated with virulence and pathogenesis [9,115,116,117,118,119]. How these enzymes directly or indirectly influence bacterial growth has not yet been investigated in detail, however, recent studies have shown that A. fumigatus alters the environmental conditions in vitro, by converting a nutrient-poor, nitrate-rich environment into one rich in amino acids. These conditions, known to exist in the CF airways, may enable P. aeruginosa to outcompete A. fumigatus by promoting a metabolic-driven increase in bacterial growth [113]. Analysis of the culture filtrates produced by A. fumigatus identified an abundance of degradative enzymes which are also involved in virulence, including alkaline protease 1, alkaline protease 2, aspergillopepsin-1, and major allergen Asp f 2 [115,117,118,119]. The increase in bacterial growth owing to the presence of A. fumigatus may affect the ability of host epithelial cells to efficiently internalize incoming pathogens and participate in microbial clearance [120]. This may be exacerbated by A. fumigatus-mediated inhibition of host cell apoptosis [121,122,123].
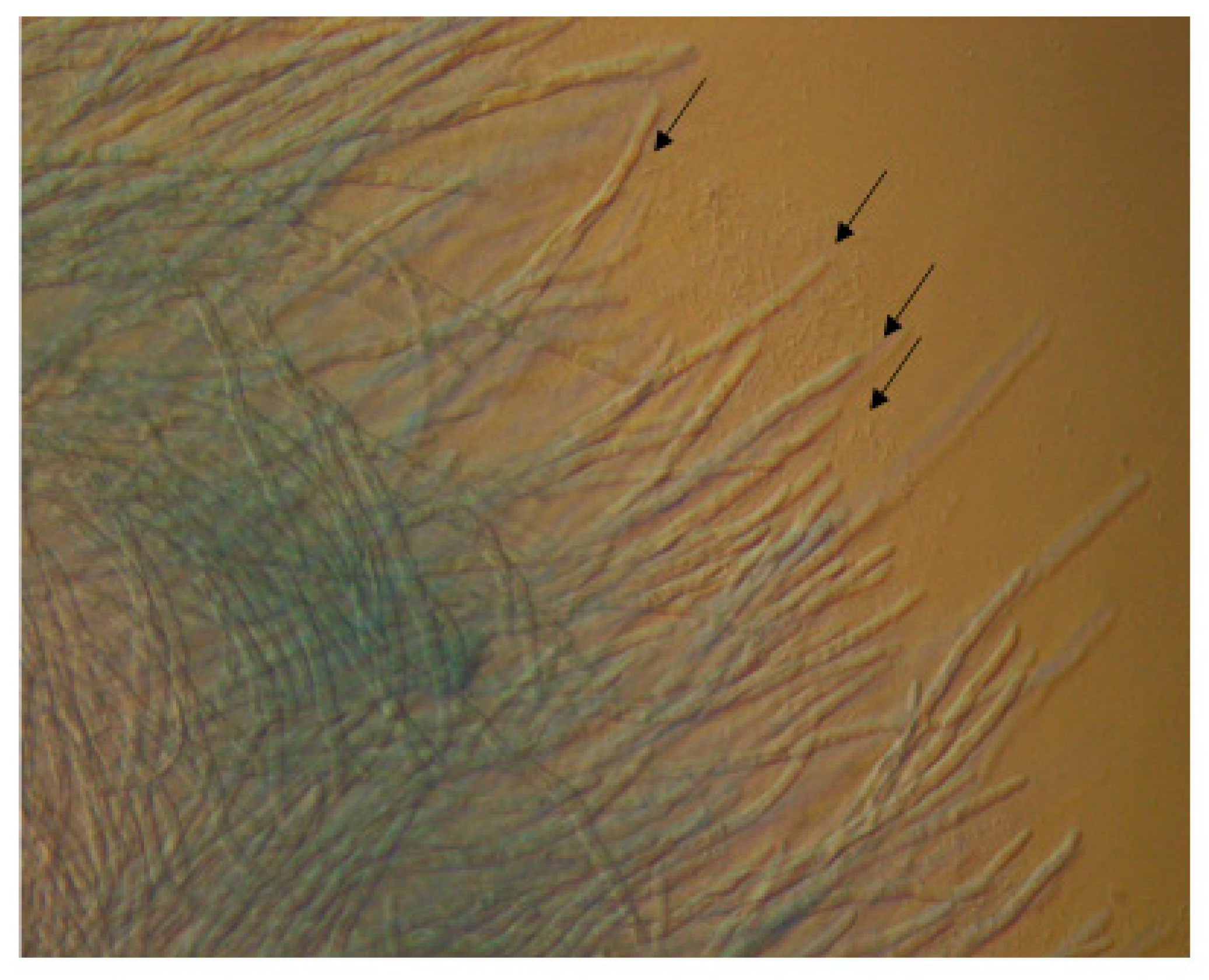
On semi-solid nutrient agar plates, P. aeruginosa cells can travel from one area of the plate toward the developing hyphae of A. fumigatus at another area, and form a cluster around the hyphal tips (Figure 2). This may be caused by an area of increased nutrient availability for the bacterial cells and indicates the ability of bacterial cells to interact directly with the fungus.
While the relationship between P. aeruginosa–A. fumigatus is antagonistic for the most part, there is increasing evidence to show that P. aeruginosa volatile organic compounds (VOC) stimulate the growth of A. fumigatus without the requirement for direct contact between the pathogens [101,124]. Volatile sulfur compounds (VSC) such as dimethyl sulfide (DMS) released by P. aeruginosa provide A. fumigatus with a sulfur source, which is necessary for fungal growth [124]. In the CF airways, P. aeruginosa releases VOCs [125], thus the VOC-mediated stimulation of fungal growth may facilitate the persistence of A. fumigatus in the lungs.
6. Interactions Between A. fumigatus and Other Pulmonary Pathogens
With the exception of P. aeruginosa, the interactions between A. fumigatus and other pulmonary pathogens remain relatively unexplored, although this is changing as the recognition for the impact of polymicrobial interactions involving this pathogen on disease progression begin to surface [6]. A better understanding of these dynamics may help predict the treatment regimens necessary to ameliorate pulmonary infections.
While bacteria such as S. aureus are associated with chronic colonization in juvenile CF patients, A. fumigatus persists throughout the lifetime of individuals with CF, but rarely establishes chronic infection [64,126,127]. Co-cultures of S. aureus and A. fumigatus conidia revealed antagonistic interactions resulting in the bacteria outcompeting the fungus [18]. In this study, S. aureus cells adhered to conidia and fungal-bound bacteria served as a chemoattractant for other bacterial cells. Fungal inhibition by S. aureus was most effective where bacteria adhered to the surface first. Bacteria induced lysis of the conidia and interfered with hyphal development [18].
The Gram-positive bacterium, Streptococcus pneumoniae, is the causative agent of pneumonia and sepsis in elderly people and children [128,129,130]. These bacteria are also detected in the airways of CF patients and associated with pulmonary exacerbations, particularly in children [131,132,133]. S. pneumoniae inhibit the development of A. fumigatus in vitro, and disassemble pre-formed fungal biofilm, the mechanism for which is regulated by pneumolysin and hydrogen peroxide, which bacteria produce as a byproduct of aerobic respiration [134].
Although Klebsiella pneumoniae is not typically associated with CF infections, it is nonetheless a common cause of pulmonary disease [135,136]. In vitro, in mixed biofilms, K. pneumoniae suppressed A. fumigatus conidial germination, hyphal development, and biofilm formation without killing the fungus [137]. On the contrary, K. pneumoniae biofilm increased in the presence of A. fumigatus. These effects were dependent upon direct contact between the fungal and bacterial pathogens in which K. pneumoniae induced oxidative stress and upregulation of cell wall synthesis genes in A. fumigatus [137].
Stenotrophomonas maltophilia is an emerging CF-associated pathogen [138]. Interactions between S. maltophilia and A. fumigatus were analyzed in mixed biofilms and, similar to that which was observed during studies using S. aureus and K. pneumoniae, the results showed that S. maltophilia interacted directly with fungal biofilm and in the presence of bacteria, A. fumigatus hyphal formation was delayed, conidiation was abrogated and biofilm formation was reduced. Moreover, the conidial cell wall was thicker in the presence of S. maltophilia.
These interaction studies indicate that while bacteria outcompete A. fumigatus in terms of growth, such encounters do not kill the fungus, but rather subdue its ability to become invasive. In the context of CF and asthma, this may be clinically relevant, because although A. fumigatus does not become invasive, it does persist and induce prolonged inflammation [139,140]. The propensity for the bacteria discussed here to disrupt fungal biofilm formation suggest that the pathogens do not co-exist in biofilms. However, these bacteria are frequently isolated with A. fumigatus from the immunocompromised airways, thereby indicating that although co-infections exist, the pathogens may be spatially segregated. The implications for this is the occurrence of colonization by multiple microbial species in different areas of the respiratory system, which may necessitate tailored therapeutic strategies.
7. Conclusions
For a long time, bacteria were thought to be the main drivers of disease in the immunocompromised airways [90]. However, advanced molecular techniques have identified fungal pathogens as a major contributing factor in the onset and development of pulmonary infections. In particular, A. fumigatus is recognized as one of the most prevalent pathogens associated with the lungs, rarely becoming invasive, but regularly inducing a hypersensitive response in the patient. The way in which A. fumigatus interacts with other members of the pulmonary ecosystem is fundamental to understanding how this pathogen competes with others to establish infection or facilitates the establishment of other pathogens. The consequences of synergistic and antagonistic interactions arising from co-infection with A. fumigatus and other bacteria may have serious implications for the respiratory health of patients and the importance of understanding how these pathogens interact is underpinned by the negative impact of co-infection between A. fumigatus and P. aeruginosa in CF patients. Thus, understanding the dynamics of the relationship between these pathogens is fundamental for the development of targeted therapeutics that may disturb these interactions and improve patient health.
Author Contributions
A.M. wrote the manuscript, J.C.C. and K.K. prepared it for submission. All authors have read and agreed to the published version of the manuscript.
Funding
A.M. is a recipient of an Irish Research Council Doctoral Scholarship.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are grateful for the assistance of Gemma Lynch in capturing the images of P. aeruginosa cells clustering around A. fumigatus hyphae.
Conflicts of Interest
The authors declared no conflict of interest.
References
- Peters, B.M.; Jabra-rizk, M.A.; Costerton, J.W.; Shirtliff, M.E. Polymicrobial Interactions: Impact on Pathogenesis and Human Disease. Clin. Microbiol Rev. 2012, 12, 193–213. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.L.; Connell, J.L.; Stacy, A.; Turner, K.H.; Whiteley, M. Mechanisms of synergy in polymicrobial infections. J. Microbiol. 2014, 52, 188–199. [Google Scholar] [CrossRef]
- Gabrilska, R.A.; Rumbaugh, K.P. Biofilm models of polymicrobial infection. Future Microbiol. 2015, 10, 1997–2015. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Hu, M.; Yu, K.; Zeng, X.; Liu, X. Mass spectrometry-based proteomic approaches to study pathogenic bacteria-host interactions. Protein Cell. 2015, 6, 265–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darch, S.E.; Ibberson, C.B.; Whiteley, M. Evolution of Bacterial “Frenemies”. MBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Filkins, L.M.; O’Toole, G.A. Cystic Fibrosis Lung Infections: Polymicrobial, Complex, and Hard to Treat. PLoS Pathog. 2015, 11, e1005258. [Google Scholar] [CrossRef] [Green Version]
- Kosmidis, C.; Denning, D.W. The clinical spectrum of pulmonary aspergillosis. Postgrad. Med. J. 2015, 91, 403–410. [Google Scholar] [CrossRef]
- Latgé, J.P. Aspergillus fumigatus and Aspergillosis. Clin. Microbiol. Rev. 1999, 12, 310–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulussen, C.; Hallsworth, J.E.; Álvarez-Pérez, S.; Nierman, W.C.; Hamill, P.G.; Blain, D.; Rediers, H.; Lievens, B. Ecology of aspergillosis: Insights into the pathogenic potency of Aspergillus fumigatus and some other Aspergillus species. Microb. Biotechnol. 2017, 10, 296–322. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.C.; Tsai, H.-F.; Karos, M.; Kwon-Chung, K.J. THTA, a thermotolerance gene of Aspergillus fumigatus. Fungal Genet. Biol. 2004, 41, 888–896. [Google Scholar] [CrossRef]
- Bignell, E.; Negrete-Urtasun, S.; Calcagno, A.M.; Haynes, K.; Arst, H.N., Jr.; Rogers, T. The Aspergillus pH-responsive transcription factor PacC regulates virulence. Mol. Microbiol. 2005, 55, 1072–1084. [Google Scholar] [CrossRef] [PubMed]
- Bertuzzi, M.; Schrettl, M.; Alcazar-Fuoli, L.; Cairns, T.C.; Munoz, A.; Walker, L.A.; Herbst, S.; Safari, M.; Cheverton, A.M.; Chen, D.; et al. The pH-responsive PacC transcription factor of Aspergillus fumigatus governs epithelial entry and tissue invasion during pulmonary aspergillosis. PLoS Pathog. 2014, 10, e1004413. [Google Scholar] [CrossRef] [Green Version]
- Beauvais, A.; Schmidt, C.; Guadagnini, S.; Roux, P.; Perret, E.; Henry, C.; Paris, S.; Mallet, A.; Prévost, M.-C.; Latgé, J. An extracellular matrix glues together the aerial-grown hyphae of Aspergillus fumigatus. Cell. Microbiol. 2007, 9, 1588–1600. [Google Scholar] [CrossRef]
- Müller, F.-M.C.; Seidler, M.; Beauvais, A. Aspergillus fumigatus biofilms in the clinical setting. Med. Mycol. 2011, 49, S96–S100. [Google Scholar] [CrossRef] [Green Version]
- Mowat, E.; Williams, C.; Jones, B.; McChlery, S.; Ramage, G. The characteristics of Aspergillus fumigatus mycetoma development: Is this a biofilm? Med. Mycol. 2009, 47, S120–S126. [Google Scholar] [CrossRef] [Green Version]
- Mowat, E.; Rajendran, R.; Williams, C.; McCulloch, E.; Jones, B.; Lang, S.; Ramage, G. Pseudomonas aeruginosa and their small diffusible extracellular molecules inhibit Aspergillus fumigatus biofilm formation. FEMS Microbiol. Lett. 2010, 313, 96–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manavathu, E.K.; Vager, D.L.; Vazquez, J.A. Development and antimicrobial susceptibility studies of in vitro monomicrobial and polymicrobial biofilm models with Aspergillus fumigatus and Pseudomonas aeruginosa. BMC Microbiol. 2014, 14, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez Granillo, A.; Canales, M.G.M.; Espíndola, M.E.S.; Martínez Rivera, M.A.; de Lucio, V.M.B.; Tovar, A.V.R. Antibiosis interaction of Staphylococccus aureus on Aspergillus fumigatus assessed in vitro by mixed biofilm formation. BMC Microbiol. 2015, 15, 33. [Google Scholar] [CrossRef] [Green Version]
- De Jesus Carrion, S.; Leal, S.M., Jr.; Ghannoum, M.A.; Aimanianda, V.; Latgé, J.-P.; Pearlman, E. The RodA hydrophobin on Aspergillus fumigatus spores masks dectin-1- and dectin-2-dependent responses and enhances fungal survival in vivo. J. Immunol. 2013, 191, 2581–2588. [Google Scholar] [CrossRef] [Green Version]
- Margalit, A.; Kavanagh, K. The innate immune response to Aspergillus fumigatus at the alveolar surface. FEMS Microbiol. Rev. 2015, 39, 670–687. [Google Scholar] [CrossRef] [Green Version]
- Heinekamp, T.; Thywißen, A.; Macheleidt, J.; Keller, S.; Valiante, V.; Brakhage, A.A. Aspergillus fumigatus melanins: Interference with the host endocytosis pathway and impact on virulence. Front. Microbiol. 2013, 3, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahn, B.; Langfelder, K.; Schneider, U.; Schindel, C.; Brakhage, A.A. PKSP-dependent reduction of phagolysosome fusion and intracellular kill of Aspergillus fumigatus conidia by human monocyte-derived macrophages. Cell. Microbiol. 2002, 4, 793–803. [Google Scholar] [CrossRef]
- Pal, A.K.; Gajjar, D.U.; Vasavada, A.R. DOPA and DHN pathway orchestrate melanin synthesis in Aspergillus species. Med. Mycol. 2013, 52, 10–18. [Google Scholar]
- Chabi, M.L.; Goracci, A.; Roche, N.; Paugam, A.; Lupo, A.; Revel, M.P. Pulmonary aspergillosis. Diagn. Interv. Imaging. 2015, 96, 435–442. [Google Scholar] [CrossRef] [Green Version]
- Hope, W.W.; Walsh, T.J.; Denning, D.W. The invasive and saprophytic syndromes due to Aspergillus spp. Med. Mycol. 2005, 43, S207–S238. [Google Scholar] [CrossRef] [Green Version]
- Stevens, D.A.; Moss, R.B.; Kurup, V.P.; Knutsen, A.P.; Greenberger, P.; Judson, M.A.; Denning, D.W.; Crameri, R.; Brody, A.S.; Light, M.; et al. Allergic bronchopulmonary aspergillosis in cystic fibrosis—state of the art: Cystic Fibrosis Foundation Consensus Conference. Clin. Infect. Dis. 2003, 37, S225–S264. [Google Scholar] [CrossRef]
- Janahi, I.A.; Rehman, A.; Al-Naimi, A.R. Allergic bronchopulmonary aspergillosis in patients with cystic fibrosis. Ann. Thorac. Med. 2017, 12, 74–82. [Google Scholar] [CrossRef]
- Patterson, K.; Strek, M.E. Allergic Bronchopulmonary Aspergillosis. Proc. Am. Thorac. Soc. 2010, 7, 237–244. [Google Scholar] [CrossRef]
- Tanner, N.T.; Judson, M.A. Diagnosis and treatment of allergic bronchopulmonary aspergillosis. Curr. Fungal Infect. Rep. 2008, 2, 199. [Google Scholar] [CrossRef]
- Knutsen, A.P.; Hutcheson, P.S.; Slavin, R.G.; Kurup, V.P. IgE antibody to Aspergillus fumigatus recombinant allergens in cystic fibrosis patients with allergic bronchopulmonary aspergillosis. Allergy 2004, 59, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, R.; Maskey, D.; Aggarwal, A.N.; Saikia, B.; Garg, M.; Gupta, D.; Chakrabarti, A. Diagnostic Performance of Various Tests and Criteria Employed in Allergic Bronchopulmonary Aspergillosis: A Latent Class Analysis. PLoS ONE 2013, 8, e61105. [Google Scholar] [CrossRef] [Green Version]
- Knutsen, A.P.; Slavin, R.G. Allergic bronchopulmonary aspergillosis in asthma and cystic fibrosis. Clin. Dev. Immunol. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farnell, E.; Rousseau, K.; Thornton, D.J.; Bowyer, P.; Herrick, S.E. Expression and secretion of Aspergillus fumigatus proteases are regulated in response to different protein substrates. Fungal Biol. 2012, 116, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Daly, P.; Kavanagh, K. Pulmonary aspergillosis: Clinical presentation, diagnosis and therapy. Br. J. Biomed. Sci. 2001, 58, 197–205. [Google Scholar]
- Kogan, T.V.; Jadoun, J.; Mittelman, L.; Hirschberg, K.; Osherov, N. Involvement of Secreted Aspergillus fumigatus Proteases in Disruption of the Actin Fiber Cytoskeleton and Loss of Focal Adhesion Sites in Infected A549 Lung Pneumocytes. J. Infect. Dis. 2004, 189, 1965–1973. [Google Scholar] [CrossRef] [Green Version]
- Amitani, R.; Taylor, G.; Elezis, E.N.; Llewellyn-Jones, C.; Mitchell, J.; Kuze, F.; Cole, P.J.; Wilson, R. Purification and characterization of factors produced by Aspergillus fumigatus which affect human ciliated respiratory epithelium. Infect. Immun. 1995, 63, 3266–3271. [Google Scholar] [CrossRef] [Green Version]
- Caminati, M.; Le Pham, D.; Bagnasco, D.; Canonica, G.W. Type 2 immunity in asthma. World Allergy Organ. J. 2018, 11, 13. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Homer, R.J.; Wang, Z.; Chen, Q.; Geba, G.P.; Wang, J.; Zhang, Y.; Elias, J.A. Pulmonary expression of interleukin-13 causes inflammation, mucus hypersecretion, subepithelial fibrosis, physiologic abnormalities, and eotaxin production. J. Clin. Investig. 1999, 103, 779–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahy, J.V. Type 2 inflammation in asthma—present in most, absent in many. Nat. Rev. Immunol. 2015, 15, 57–65. [Google Scholar] [CrossRef]
- de Almeida, M.B.; Bussamra, M.H.F.; Rodrigues, J.C. Allergic bronchopulmonary aspergillosis in paediatric cystic fibrosis patients. Paediatr. Respir. Rev. 2006, 7, 67–72. [Google Scholar]
- Burgel, P.-R.; Paugam, A.; Hubert, D.; Martin, C. Aspergillus fumigatus in the cystic fibrosis lung: Pros and cons of azole therapy. Infect. Drug Resist. 2016, 9, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Reece, E.; McClean, S.; Greally, P.; Renwick, J. The prevalence of Aspergillus fumigatus in early cystic fibrosis disease is underestimated by culture-based diagnostic methods. J. Microbiol. Methods 2019, 164, 105683. [Google Scholar] [CrossRef]
- Breuer, O.; Schultz, A.; Garratt, L.W.; Turkovic, L.; Rosenow, T.; Murray, C.P.; Karpievitch, Y.V.; Akesson, L.; Dalton, S.; Sly, P.D.; et al. Aspergillus Infections and Progression of Structural Lung Disease in Children with Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 2019, 201, 688–696. [Google Scholar] [CrossRef]
- Breuer, O.; Schultz, A.; Turkovic, L.; de Klerk, N.; Keil, A.D.; Brennan, S.; Harrison, J.; Robertson, C.; Robinson, P.J.; Sly, P.D.; et al. Changing Prevalence of Lower Airway Infections in Young Children with Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 2019, 200, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Saunders, R.V.; Modha, D.E.; Claydon, A.; Gaillard, E.A. Chronic Aspergillus fumigatus colonization of the pediatric cystic fibrosis airway is common and may be associated with a more rapid decline in lung function. Med. Mycol. 2016, 54, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Harun, S.N.; Wainwright, C.E.; Grimwood, K.; Hennig, S. Aspergillus and progression of lung disease in children with cystic fibrosis. Thorax 2019, 74, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Reece, E.; Segurado, R.; Jackson, A.; McClean, S.; Renwick, J.; Greally, P. Co-colonisation with Aspergillus fumigatus and Pseudomonasa eruginosais associated with poorer health in cystic fibrosis patients: An Irish registry analysis. BMC Pulm. Med. 2017, 17, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.L.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, E.P. Pump up the versatility. Nature 2000, 406, 947–948. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Jesaitis, A.J.; Franklin, M.J.; Berglund, D.; Sasaki, M.; Lord, C.I.; Bleazard, J.B.; Duffy, J.E.; Beyenal, H.; Lewandowski, Z. Compromised host defense on Pseudomonasaeruginosa biofilms: Characterization of neutrophil and biofilm interactions. J. Immunol. 2003, 171, 4329–4339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsek, M.R.; Greenberg, E.P. Sociomicrobiology: The connections between quorum sensing and biofilms. Trends Microbiol. 2005, 13, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.Ø.; Bjarnsholt, T.; Phipps, R.; Rasmussen, T.B.; Calum, H.; Christoffersen, L.; Moser, C.; Williams, P.; Pressler, T.; Givskov, M.; et al. Rapid necrotic killing of polymorphonuclear leukocytes is caused by quorum-sensing-controlled production of rhamnolipid by Pseudomonas aeruginosa. Microbiology 2007, 153, 1329–1338. [Google Scholar] [CrossRef] [Green Version]
- Managò, A.; Becker, K.A.; Carpinteiro, A.; Wilker, B.; Soddemann, M.; Seitz, A.P.; Edwards, M.J.; Grassmé, H.; Szabò, I.; Gulbins, E. Pseudomonas aeruginosa Pyocyanin Induces Neutrophil Death via Mitochondrial Reactive Oxygen Species and Mitochondrial Acid Sphingomyelinase. Antioxid. Redox Signal. 2015, 22, 1097–1110. [Google Scholar]
- Singh, P.K.; Parsek, M.R.; Greenberg, E.P.; Welsh, M.J. A component of innate immunity prevents bacterial biofilm development. Nature 2002, 417, 552–555. [Google Scholar] [CrossRef]
- Stintzi, A.; Evans, K.; Meyer, J.M.; Poole, K. Quorum-sensing and siderophore biosynthesis in Pseudomonas aeruginosa: lasR/lasI mutants exhibit reduced pyoverdine biosynthesis. FEMS Microbiol. Lett. 1998, 166, 341–345. [Google Scholar] [CrossRef]
- Marvig, R.L.; Sommer, L.M.; Molin, S.; Johansen, H.K. Convergent evolution and adaptation of Pseudomonas aeruginosa within patients with cystic fibrosis. Nat. Genet. 2014, 47, 57. [Google Scholar] [CrossRef] [PubMed]
- Mauch, R.M.; Jensen, P.O.; Moser, C.; Levy, C.E.; Hoiby, N. Mechanisms of humoral immune response against Pseudomonas aeruginosa biofilm infection in cystic fibrosis. J. Cyst. Fibros. 2018, 17, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Wolfgang, M.C.; Jyot, J.; Goodman, A.L.; Ramphal, R.; Lory, S. Pseudomonas aeruginosa regulates flagellin expression as part of a global response to airway fluid from cystic fibrosis patients. Proc. Natl. Acad. Sci. USA 2004, 101, 6664–6668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, L.R.; Kulasekara, H.D.; Emerson, J.; Houston, L.S.; Burns, J.L.; Ramsey, B.W.; Miller, S.I. Pseudomonas aeruginosa lasR mutants are associated with cystic fibrosis lung disease progression. J. Cyst. Fibros. 2009, 8, 66–70. [Google Scholar] [CrossRef] [Green Version]
- Jain, M.; Ramirez, D.; Seshadri, R.; Cullina, J.F.; Powers, C.A.; Schulert, G.S.; Bar-Meir, M.; Sullivan, C.L.; McColley, S.A.; Hauser, A.R. Type III secretion phenotypes of Pseudomonas aeruginosa strains change during infection of individuals with cystic fibrosis. J. Clin. Microbiol. 2004, 42, 5229–5237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filkins, L.M.; Hampton, T.H.; Gifford, A.H.; Gross, M.J.; Hogan, D.A.; Sogin, M.L.; Morrison, H.G.; Paster, B.J.; O’Toole, G.A. Prevalence of streptococci and increased polymicrobial diversity associated with cystic fibrosis patient stability. J. Bacteriol. 2012, 194, 4709–4717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, F. Microbial ecology of the cystic fibrosis lung. Microbiology 2007, 153, 917–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Cystic Fibriosis Registry of Ireland. CF Registry of Ireland 2017 Annual Report; The Cystic Fibriosis Registry of Ireland: Dublin, Ireland, 2017; p. 47. [Google Scholar]
- Zemanick, E.T.; Wagner, B.D.; Robertson, C.E.; Ahrens, R.C.; Chmiel, J.F.; Clancy, J.P.; Gibson, R.L.; Harris, W.T.; Kurland, G.; Laguna, T.A.; et al. Airway microbiota across age and disease spectrum in cystic fibrosis. Eur. Respir. J. 2017, 50, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Coburn, B.; Wang, P.W.; Caballero, J.D.; Clark, S.T.; Brahma, V.; Donaldson, S.; Zhang, Y.; Surendra, A.; Gong, Y.; Tullis, D.E.; et al. Lung microbiota across age and disease stage in cystic fibrosis. Sci. Rep. 2015, 5, 10241. [Google Scholar] [CrossRef] [PubMed]
- Cystic Fibrosis Foundation Patient Registry. 2017 Annual Data Report; Cystic Fibrosis Foundation Patient Registry: Bethesda, MD, USA, 2018. [Google Scholar]
- Cystic Fibrosis Trust. UK Cystic Fibrosis Registry Annual Data Report; Cystic Fibrosis Trust: London, UK, 2017. [Google Scholar]
- Riley, T.V.; Hoffman, D.C. Interference with Haemophilus influenzae growth by other microorganisms. FEMS Microbiol. Lett. 1986, 33, 55–58. [Google Scholar] [CrossRef]
- Baldan, R.; Cigana, C.; Testa, F.; Bianconi, I.; De Simone, M.; Pellin, D.; Di Serio, C.; Bragonzi, A.; Cirillo, D.M. Adaptation of Pseudomonas aeruginosa in Cystic Fibrosis Airways Influences Virulence of Staphylococcus aureus in vitro and Murine Models of Co-Infection. PLoS ONE 2014, 9, e89614. [Google Scholar] [CrossRef]
- Purcell, P.; Jary, H.; Perry, A.; Perry, J.D.; Stewart, C.J.; Nelson, A.; Lanyon, C.; Smith, D.L.; Cummings, S.P.; De Soyza, A. Polymicrobial airway bacterial communities in adult bronchiectasis patients. BMC Microbiol. 2014, 14, 130. [Google Scholar] [CrossRef] [Green Version]
- Woo, T.E.; Lim, R.; Heirali, A.A.; Acosta, N.; Rabin, H.R.; Mody, C.H.; Somayaji, R.; Surette, M.G.; Sibley, C.D.; Storey, D.G.; et al. A longitudinal characterization of the Non-Cystic Fibrosis Bronchiectasis airway microbiome. Sci. Rep. 2019, 9, 6871. [Google Scholar] [CrossRef]
- Gallego, M.; Pomares, X.; Espasa, M.; Castañer, E.; Solé, M.; Suárez, D.; Monsó, E.; Montón, C. Pseudomonas aeruginosa isolates in severe chronic obstructive pulmonary disease: Characterization and risk factors. BMC Pulm. Med. 2014, 14, 103. [Google Scholar] [CrossRef] [Green Version]
- Murphy, T.F.; Brauer, A.L.; Eschberger, K.; Lobbins, P.; Grove, L.; Cai, X.; Sethi, S. Pseudomonas aeruginosa in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2008, 177, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Araújo, D.; Shteinberg, M.; Aliberti, S.; Goeminne, P.C.; Hill, A.T.; Fardon, T.C.; Obradovic, D.; Stone, G.; Trautmann, M.; Davis, A.; et al. The independent contribution of Pseudomonas aeruginosa infection to long-term clinical outcomes in bronchiectasis. Eur. Respir. J. 2018, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, Y.-H.; Xu, J.-F. How does Pseudomonas aeruginosa affect the progression of bronchiectasis? Clin. Microbiol. Infect. 2020, 26, 313–318. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, M.J.; Jary, H.R.; Perry, A.; MacFarlane, J.G.; Hester, K.L.M.; Small, T.; Molyneux, C.; Perry, J.D.; Walton, K.E.; De Soyza, A. Non cystic fibrosis bronchiectasis: A longitudinal retrospective observational cohort study of Pseudomonas persistence and resistance. Respir. Med. 2015, 109, 716–726. [Google Scholar] [CrossRef] [Green Version]
- Kaur, J.; Pethani, B.P.; Kumar, S.; Kim, M.; Sunna, A.; Kautto, L.; Penesyan, A.; Paulsen, I.T.; Nevalainen, H. Pseudomonas aeruginosa inhibits the growth of Scedosporium aurantiacum, an opportunistic fungal pathogen isolated from the lungs of cystic fibrosis patients. Front. Microbiol. 2015, 6, 866. [Google Scholar] [CrossRef] [Green Version]
- Bandara, H.M.H.N.; Yau, J.Y.Y.; Watt, R.M.; Jin, L.J.; Samaranayake, L.P. Pseudomonas aeruginosa inhibits in vitro Candida biofilm development. BMC Microbiol. 2010, 10, 125. [Google Scholar] [CrossRef] [Green Version]
- Rella, A.; Yang, M.W.; Gruber, J.; Montagna, M.T.; Luberto, C.; Zhang, Y.-M.; Del Poeta, M. Pseudomonas aeruginosa inhibits the growth of Cryptococcus species. Mycopathologia 2012, 173, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Cheng, W.; He, X.; Liu, Y. The co-colonization prevalence of Pseudomonas aeruginosa and Aspergillus fumigatus in cystic fibrosis: A systematic review and meta-analysis. Microb. Pathog. 2018, 125, 122–128. [Google Scholar] [CrossRef]
- Paugam, A.; Baixench, M.-T.; Demazes-Dufeu, N.; Burgel, P.-R.; Sauter, E.; Kanaan, R.; Dusser, D.; Dupouy-Camet, J.; Hubert, D. Characteristics and consequences of airway colonization by filamentous fungi in 201 adult patients with cystic fibrosis in France. Med. Mycol. 2010, 48, S32–S36. [Google Scholar] [CrossRef] [Green Version]
- Hector, A.; Kirn, T.; Ralhan, A.; Graepler-Mainka, U.; Berenbrinker, S.; Riethmueller, J.; Hogardt, M.; Wagner, M.; Pfleger, A.; Autenrieth, I.; et al. Microbial colonization and lung function in adolescents with cystic fibrosis. J. Cyst. Fibros. 2016, 15, 340–349. [Google Scholar] [CrossRef] [Green Version]
- Huerta, A.; Soler, N.; Esperatti, M.; Guerrero, M.; Menendez, R.; Gimeno, A.; Zalacaín, R.; Mir, N.; Aguado, J.M.; Torres, A. Importance of Aspergillus spp. isolation in Acute exacerbations of severe COPD: Prevalence, factors and follow-up: The Fungi-COPD study. Respir. Res. 2014, 15, 17. [Google Scholar] [CrossRef] [Green Version]
- Everaerts, S.; Lagrou, K.; Vermeersch, K.; Dupont, L.J.; Vanaudenaerde, B.M.; Janssens, W. Aspergillus fumigatus Detection and Risk Factors in Patients with COPD-Bronchiectasis Overlap. Int. J. Mol. Sci. 2018, 19, 523. [Google Scholar] [CrossRef] [Green Version]
- Máiz, L.; Nieto, R.; Cantón, R.; Gómez de la Pedrosa, E.; Martinez-García, M.Á. Fungi in Bronchiectasis: A Concise Review. Int. J. Mol. Sci. 2018, 19, 142. [Google Scholar] [CrossRef] [Green Version]
- Moss, R.B. Fungi in cystic fibrosis and non-cystic fibrosis bronchiectasis. Semin. Respir. Crit. Care Med. 2015, 36, 207–216. [Google Scholar] [CrossRef]
- Everaerts, S.; Lagrou, K.; Dubbeldam, A.; Lorent, N.; Vermeersch, K.; Van Hoeyveld, E.; Bossuyt, X.; Dupont, L.J.; Vanaudenaerde, B.M.; Janssens, W. Sensitization to Aspergillus fumigatus as a risk factor for bronchiectasis in COPD. Int. J. Chron. Obstruct. Pulmon. Dis. 2017, 12, 2629–2638. [Google Scholar] [CrossRef] [Green Version]
- Bilton, D. Update on non-cystic fibrosis bronchiectasis. Curr. Opin. Pulm. Med. 2008, 14, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.; Ranjendran, R.; Ramage, G. Pathogenesis of Fungal Infections in Cystic Fibrosis. Curr. Fungal Infect. Rep. 2016, 10, 163–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briard, B.; Mislin, G.L.A.; Latgé, J.-P.; Beauvais, A. Interactions between Aspergillus fumigatus and Pulmonary Bacteria: Current State of the Field, New Data, and Future Perspective. J. Fungi (Basel, Switzerland) 2019, 5, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, S.; Fothergill, J.L. The role of multispecies social interactions in shaping Pseudomonas aeruginosa pathogenicity in the cystic fibrosis lung. FEMS Microbiol. Lett. 2017, 364, fnx128. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.E.; Buckley, D.G.; Wu, Z.; Saenphimmachak, C.; Hoffman, L.R.; D’Argenio, D.A.; Miller, S.I.; Ramsey, B.W.; Speert, D.P.; Moskowitz, S.M.; et al. Genetic adaptation by Pseudomonas aeruginosa to the airways of cystic fibrosis patients. Proc. Natl. Acad. Sci. USA 2006, 103, 8487–8492. [Google Scholar] [CrossRef] [Green Version]
- Sass, G.; Nazik, H.; Penner, J.; Shah, H.; Ansari, S.R.; Clemons, K.; Groleau, M.-C.; Dietl, A.-M.; Visca, P.; Haas, H.; et al. Studies of Pseudomonas aeruginosa Mutants Indicate Pyoverdine as the Central Factor in Inhibition of Aspergillus fumigatus Biofilm. J. Bacteriol. 2018, 200, 1–24. [Google Scholar]
- Briard, B.; Bomme, P.; Lechner, B.E.; Mislin, G.L.A.; Lair, V.; Prévost, M.C.; Latgé, J.P.; Haas, H.; Beauvais, A. Pseudomonas aeruginosa manipulates redox and iron homeostasis of its microbiota partner Aspergillus fumigatus via phenazines. Sci. Rep. 2015, 5, 8220. [Google Scholar] [CrossRef] [PubMed]
- Shirazi, F.; Ferreira, J.A.G.; Stevens, D.A.; Clemons, K.V.; Kontoyiannis, D.P. Biofilm filtrates of Pseudomonas aeruginosa strains isolated from cystic fibrosis patients inhibit preformed Aspergillus fumigatus biofilms via apoptosis. PLoS ONE 2016, 11, e0150155. [Google Scholar] [CrossRef]
- Ferreira, J.A.G.; Penner, J.C.; Moss, R.B.; Haagensen, J.A.J.; Clemons, K.V.; Spormann, A.M.; Nazik, H.; Cohen, K.; Banaei, N.; Carolino, E.; et al. Inhibition of Aspergillus fumigatus and Its Biofilm by Pseudomonas aeruginosa Is Dependent on the Source, Phenotype and Growth Conditions of the Bacterium. PLoS ONE 2015, 10, e0134692. [Google Scholar] [CrossRef] [Green Version]
- Bargon, J.; Dauletbaev, N.; Köhler, B.; Wolf, M.; Posselt, H.-G.; Wagner, T.O.F. Prophylactic antibiotic therapy is associated with an increased prevalence of Aspergillus colonization in adult cystic fibrosis patients. Respir. Med. 1999, 93, 835–838. [Google Scholar] [CrossRef] [Green Version]
- Aaron, S.D.; Vandemheen, K.L.; Freitag, A.; Pedder, L.; Cameron, W.; Lavoie, A.; Paterson, N.; Wilcox, P.; Rabin, H.; Tullis, E.; et al. Treatment of Aspergillus fumigatus in Patients with Cystic Fibrosis: A Randomized, Placebo-Controlled Pilot Study. PLoS ONE 2012, 7, e36077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reece, E.; Doyle, S.; Greally, P.; Renwick, J.; McClean, S. Aspergillus fumigatus inhibits Pseudomonas aeruginosa in co-culture: Implications of a mutually antagonistic relationship on virulence and inflammation in the CF airway. Front. Microbiol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briard, B.; Heddergott, C.; Latgé, J. Volatile compounds emitted by pseudomonas aeruginosa stimulate growth of the fungal pathogen Aspergillus fumigatus. MBio 2016, 7, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briard, B.; Rasoldier, V.; Bomme, P.; El Aouad, N.; Guerreiro, C.; Chassagne, P.; Muszkieta, L.; Latge, J.-P.; Mulard, L.; Beauvais, A. Dirhamnolipids secreted from Pseudomonas aeruginosa modify fungal susceptibility of Aspergillus fumigatus by inhibiting beta1,3 glucan synthase activity. ISME J. 2017, 11, 1578–1591. [Google Scholar] [CrossRef] [PubMed]
- Price-Whelan, A.; Dietrich, L.E.P.; Newman, D.K. Rethinking “secondary” metabolism: Physiological roles for phenazine antibiotics. Nat. Chem. Biol. 2006, 2, 71–78. [Google Scholar] [CrossRef]
- Price, C.E.; Brown, D.G.; Limoli, D.H.; Phelan, V.V.; O’Toole, G.A. Exogenous alginate protects Staphylococcus aureus from killing by Pseudomonas aeruginosa. J. Bacteriol 2020, 202, e00559-19. [Google Scholar] [CrossRef]
- Smith, K.; Rajendran, R.; Kerr, S.; Lappin, D.F.; Mackay, W.G.; Williams, C.; Ramage, G. Aspergillus fumigatus enhances elastase production in Pseudomonas aeruginosa co-cultures. Med. Mycol. 2015, 53, 645–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenman, H.C.; Casadevall, A. Synthesis and assembly of fungal melanin. Appl. Microbiol. Biotechnol. 2012, 93, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Kuang, Z.; Hao, Y.; Walling, B.E.; Jeffries, J.L.; Ohman, D.E.; Lau, G.W. Pseudomonas aeruginosa Elastase Provides an Escape from Phagocytosis by Degrading the Pulmonary Surfactant Protein-A. PLoS ONE 2011, 6, e27091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, K.; Obata, K.; Keira, T.; Miyata, R.; Hirakawa, S.; Takano, K.; Kohno, T.; Sawada, N.; Himi, T.; Kojima, T. Pseudomonas aeruginosa elastase causes transient disruption of tight junctions and downregulation of PAR-2 in human nasal epithelial cells. Respir. Res. 2014, 15, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariencheck, W.I.; Alcorn, J.F.; Palmer, S.M.; Wright, J.R. Pseudomonas aeruginosa elastase degrades surfactant proteins A and D. Am. J. Respir. Cell Mol. Biol. 2003, 28, 528–537. [Google Scholar] [CrossRef]
- Rada, B.; Leto, T.L. Pyocyanin effects on respiratory epithelium: Relevance in Pseudomonas aeruginosa airway infections. Trends Microbiol. 2013, 21, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Denning, G.M.; Iyer, S.S.; Reszka, K.J.; O’Malley, Y.; Rasmussen, G.T.; Britigan, B.E. Phenazine-1-carboxylic acid, a secondary metabolite of Pseudomonas aeruginosa, alters expression of immunomodulatory proteins by human airway epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 285, L584–L592. [Google Scholar] [CrossRef] [Green Version]
- Sass, G.; Ansari, S.R.; Dietl, A.M.; Déziel, E.; Haas, H.; Stevens, D.A. Intermicrobial interaction: Aspergillus fumigatus siderophores protect against competition by Pseudomonas aeruginosa. PLoS ONE 2019, 14, e0216085. [Google Scholar] [CrossRef] [Green Version]
- Margalit, A.; Carolan, J.C.; Sheehan, D.; Kavanagh, K. The Aspergillus fumigatus Secretome Alters the Proteome of Pseudomonas aeruginosa to Stimulate Bacterial Growth: Implications for Co-infection. Mol. Cell. Proteom. 2020, 19, 1346–1359. [Google Scholar] [CrossRef]
- de Vries, R.P.; Visser, J. Aspergillus Enzymes Involved in Degradation of Plant Cell Wall Polysaccharides. Microbiol. Mol. Biol. Rev. 2001, 65, 497–522. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhang, L.; Zou, H.; Wang, L. Secretome profiling reveals temperature-dependent growth of Aspergillus fumigatus. Sci. China Life Sci. 2018, 61, 578–592. [Google Scholar] [CrossRef]
- Tekaia, F.; Latge, J.-P. Aspergillus fumigatus: Saprophyte or pathogen? Curr. Opin. Microbiol. 2005, 8, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Vivek-Ananth, R.P.; Mohanraj, K.; Vandanashree, M.; Jhingran, A.; Craig, J.P.; Samal, A. Comparative systems analysis of the secretome of the opportunistic pathogen Aspergillus fumigatus and other Aspergillus species. Sci. Rep. 2018, 8, 6617. [Google Scholar] [CrossRef] [PubMed]
- Wartenberg, D.; Lapp, K.; Jacobsen, I.D.; Dahse, H.-M.; Kniemeyer, O.; Heinekamp, T.; Brakhage, A.A. Secretome analysis of Aspergillus fumigatus reveals Asp-hemolysin as a major secreted protein. Int. J. Med. Microbiol. 2011, 301, 602–611. [Google Scholar] [CrossRef]
- Behnsen, J.; Lessing, F.; Schindler, S.; Wartenberg, D.; Jacobsen, I.D.; Thoen, M.; Zipfel, P.F.; Brakhage, A.A. Secreted Aspergillus fumigatus protease Alp1 degrades human complement proteins C3, C4, and C5. Infect. Immun. 2010, 78, 3585–3594. [Google Scholar] [CrossRef] [Green Version]
- Margalit, A.; Kavanagh, K.; Carolan, J.C. Characterization of the Proteomic Response of A549 Cells Following Sequential Exposure to Aspergillus fumigatus and Pseudomonas aeruginosa. J. Proteome Res. 2020, 19, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Berkova, N.; Lair-Fulleringer, S.; Féménia, F.; Huet, D.; Wagner, M.C.; Gorna, K.; Tournier, F.; Ibrahim-Granet, O.; Guillot, J.; Chermette, R.; et al. Aspergillus fumigatus conidia inhibit tumour necrosis factor- or staurosporine-induced apoptosis in epithelial cells. Int. Immunol. 2006, 18, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Daly, P.; Verhaegen, S.; Clynes, M.; Kavanagh, K. Culture filtrates of Aspergillus fumigatus induce different modes of cell death in human cancer cell lines. Mycopathologia 1999, 146, 67–74. [Google Scholar] [CrossRef]
- Féménia, F.; Huet, D.; Lair-Fulleringer, S.; Wagner, M.C.; Sarfati, J.; Shingarova, L.; Guillot, J.; Boireau, P.; Chermette, R.; Berkova, N. Effects of Conidia of Various Aspergillus Species on Apoptosis of Human Pneumocytes and Bronchial Epithelial Cells. Mycopathologia 2009, 167, 249. [Google Scholar] [CrossRef]
- Scott, J.; Sueiro-Olivares, M.; Ahmed, W.; Heddergott, C.; Zhao, C.; Thomas, R.; Bromley, M.; Latgé, J.-P.; Krappmann, S.; Fowler, S.; et al. Pseudomonas aeruginosa-Derived Volatile Sulfur Compounds Promote Distal Aspergillus fumigatus Growth and a Synergistic Pathogen-Pathogen Interaction That Increases Pathogenicity in Co-infection. Front. Microbiol. 2019, 10, 2311. [Google Scholar] [CrossRef]
- Nasir, M.; Bean, H.D.; Smolinska, A.; Rees, C.A.; Zemanick, E.T.; Hill, J.E. Volatile molecules from bronchoalveolar lavage fluid can “rule-in” Pseudomonas aeruginosa and “rule-out” Staphylococcus aureus infections in cystic fibrosis patients. Sci. Rep. 2018, 8, 826. [Google Scholar] [CrossRef]
- Hurley, M.N. Staphylococcus aureus in cystic fibrosis: Problem bug or an innocent bystander? Breathe (Sheffield, England) 2018, 14, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Delfino, E.; Del Puente, F.; Briano, F.; Sepulcri, C.; Giacobbe, D.R. Respiratory Fungal Diseases in Adult Patients with Cystic Fibrosis. Clin. Med. Insights. Circ. Respir. Pulm. Med. 2019, 13, 1179548419849939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askim, Å.; Mehl, A.; Paulsen, J.; DeWan, A.T.; Vestrheim, D.F.; Åsvold, B.O.; Damås, J.K.; Solligård, E. Epidemiology and outcome of sepsis in adult patients with Streptococcus pneumoniae infection in a Norwegian county 1993–2011: An observational study. BMC Infect. Dis. 2016, 16, 223. [Google Scholar] [CrossRef] [Green Version]
- Ostapchuk, M.; Roberts, D.M.; Haddy, R. Community-acquired pneumonia in infants and children. Am. Fam. Physician 2004, 70, 899–908. [Google Scholar]
- Asner, S.A.; Agyeman, P.K.A.; Gradoux, E.; Posfay-Barbe, K.M.; Heininger, U.; Giannoni, E.; Crisinel, P.A.; Stocker, M.; Bernhard-Stirnemann, S.; Niederer-Loher, A.; et al. Burden of Streptococcus pneumoniae Sepsis in Children After Introduction of Pneumococcal Conjugate Vaccines: A Prospective Population-based Cohort Study. Clin. Infect. Dis. 2019, 69, 1574–1580. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, J.M. Treatment of pulmonary exacerbations in cystic fibrosis. Eur. Respir. Rev. 2013, 22, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Maeda, Y.; Elborn, J.S.; Parkins, M.D.; Reihill, J.; Goldsmith, C.E.; Coulter, W.A.; Mason, C.; Millar, B.C.; Dooley, J.S.G.; Lowery, C.J.; et al. Population structure and characterization of viridans group streptococci (VGS) including Streptococcus pneumoniae isolated from adult patients with cystic fibrosis (CF). J. Cyst. Fibros. 2011, 10, 133–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paganin, P.; Fiscarelli, E.V.; Tuccio, V.; Chiancianesi, M.; Bacci, G.; Morelli, P.; Dolce, D.; Dalmastri, C.; De Alessandri, A.; Lucidi, V.; et al. Changes in Cystic Fibrosis Airway Microbial Community Associated with a Severe Decline in Lung Function. PLoS ONE 2015, 10, e0124348. [Google Scholar] [CrossRef] [Green Version]
- Iwahashi, J.; Kamei, K.; Watanabe, H. Disruption of Aspergillus fumigatus biofilm by Streptococcus pneumoniae: Mycelial fragmentation by hydrogen peroxide. J. Infect. Chemother. 2020, 26, 831–837. [Google Scholar] [CrossRef]
- LiPuma, J.J. The Changing Microbial Epidemiology in Cystic Fibrosis. Clin. Microbiol. Rev. 2010, 23, 299–323. [Google Scholar] [CrossRef] [Green Version]
- Leão, R.S.; Pereira, R.H.V.; Folescu, T.W.; Albano, R.M.; Santos, E.A.; Junior, L.G.C.; Marques, E.A. KPC-2 Carbapenemase-producing Klebsiella pneumoniae isolates from patients with Cystic Fibrosis. J. Cyst. Fibros. 2011, 10, 140–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, M.F.; Pereira, L.; Jenull, S.; Kuchler, K.; Lion, T. Klebsiella pneumoniae prevents spore germination and hyphal development of Aspergillus species. Sci. Rep. 2019, 9, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, A.; Pompilio, A.; Bettua, C.; Crocetta, V.; Giacobazzi, E.; Fiscarelli, E.; Jousson, O.; Di Bonaventura, G. Evolution of Stenotrophomonas maltophilia in Cystic Fibrosis Lung over Chronic Infection: A Genomic and Phenotypic Population Study. Front. Microbiol. 2017, 8, 1590. [Google Scholar] [CrossRef] [PubMed]
- Hartl, D. Immunological mechanisms behind the cystic fibrosis-ABPA link. Med. Mycol. 2009, 47, S183–S191. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Hoselton, S.A.; Schuh, J.M. Allergic Inflammation in Aspergillus fumigatus-Induced Fungal Asthma. Curr. Allergy Asthma Rep. 2015, 15, 59. [Google Scholar] [CrossRef]
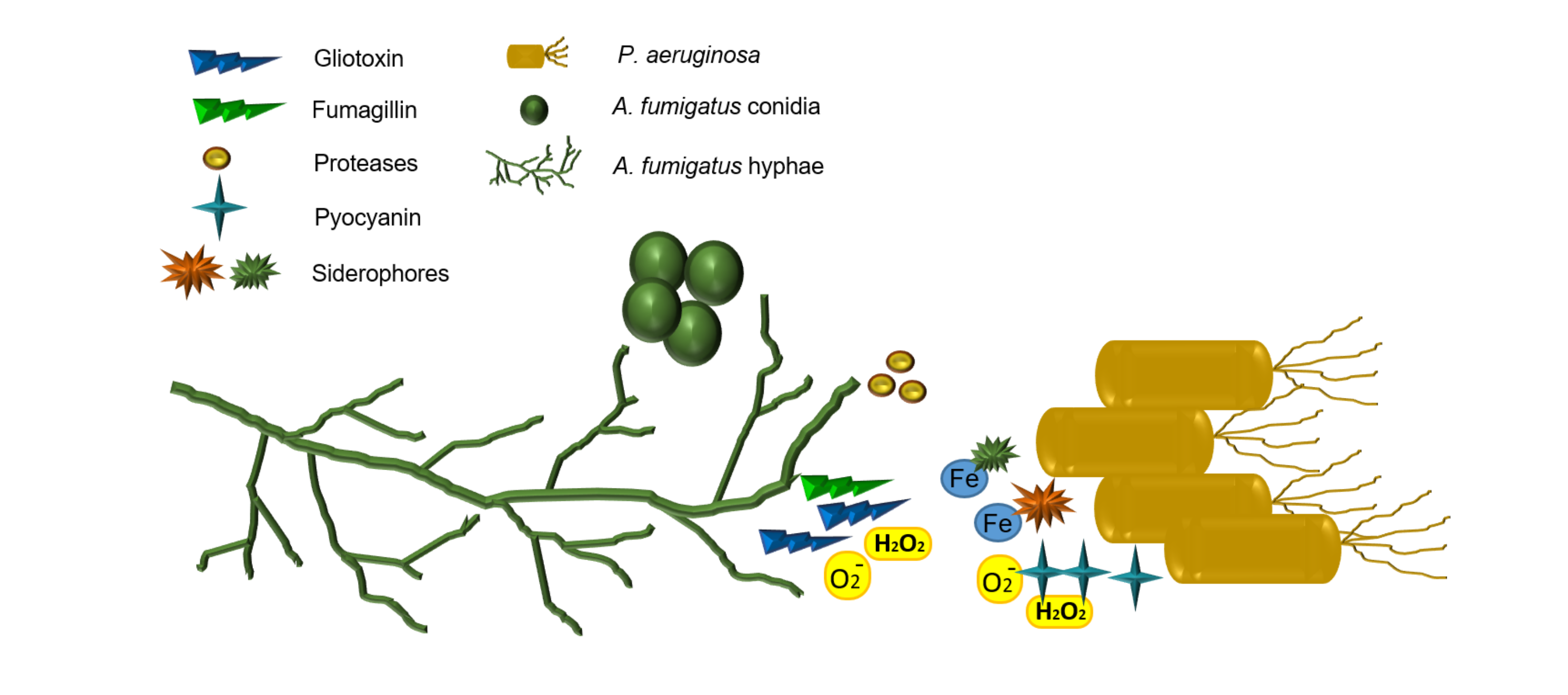
Figure 1.
(A) P. aeruginosa cells (right) grown alongside A. fumigatus conidia (left) on nutrient agar. P. aeruginosa inhibits growth of A. fumigatus as evidenced by reduced mycelial expansion on the side of bacterial growth. The green pigment produced by P. aeruginosa is pyocyanin. (B) A magnified image of (A) in which A. fumigatus growth is inhibited by P. aeruginosa; the expansion of A. fumigatus mycelia are inhibited by the close proximity to P. aeruginosa cells. By contrast, the absence of bacteria on the left hand side of the fungus allow mycelia to expand outward.
Figure 1.
(A) P. aeruginosa cells (right) grown alongside A. fumigatus conidia (left) on nutrient agar. P. aeruginosa inhibits growth of A. fumigatus as evidenced by reduced mycelial expansion on the side of bacterial growth. The green pigment produced by P. aeruginosa is pyocyanin. (B) A magnified image of (A) in which A. fumigatus growth is inhibited by P. aeruginosa; the expansion of A. fumigatus mycelia are inhibited by the close proximity to P. aeruginosa cells. By contrast, the absence of bacteria on the left hand side of the fungus allow mycelia to expand outward.

Figure 2.
P. aeruginosa cells (indicated with black arrows) travel toward A. fumigatus mycelia and cluster around the hyphal tips. Viewed through an Olympus BX61 fluorescent microscope (40X).
Figure 2.
P. aeruginosa cells (indicated with black arrows) travel toward A. fumigatus mycelia and cluster around the hyphal tips. Viewed through an Olympus BX61 fluorescent microscope (40X).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Margalit, A.; Carolan, J.C.; Kavanagh, K. Bacterial Interactions with Aspergillus fumigatus in the Immunocompromised Lung. Microorganisms 2021, 9, 435. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020435
AMA Style
Margalit A, Carolan JC, Kavanagh K. Bacterial Interactions with Aspergillus fumigatus in the Immunocompromised Lung. Microorganisms. 2021; 9(2):435. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020435
Chicago/Turabian StyleMargalit, Anatte, James C. Carolan, and Kevin Kavanagh. 2021. "Bacterial Interactions with Aspergillus fumigatus in the Immunocompromised Lung" Microorganisms 9, no. 2: 435. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020435
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.