
Characterization of the Habitat- and Season-Independent Increase in Fungal Biomass Induced by the Invasive Giant Goldenrod and Its Impact on the Fungivorous Nematode Community

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Sites
2.2. Soil Sampling
2.3. Nematode Extraction and Community Analysis
2.4. Abiotic Soil Characteristics
2.5. Fungal and Bacterial Extraction and Community Analysis
2.6. Ergosterol Measurements
2.7. PCR Amplification and Sequencing of Fungal 16S rDNA
2.8. Combined Analysis of Abiotic Characteristics and Quantitative Biotic Data
2.9. Bioinformatics Framework and Statistics
3. Results
3.1. Changes in Abiotic Soil Characteristics upon S. gigantea Invasion
3.2. Invasive S. Gigantea Increase Fungal Biomass, but Not the Total Fungal DNA
3.3. Two Fungivorous Nematode Families Benefited from S. gigantea-Induced Increase in Fungal Biomass
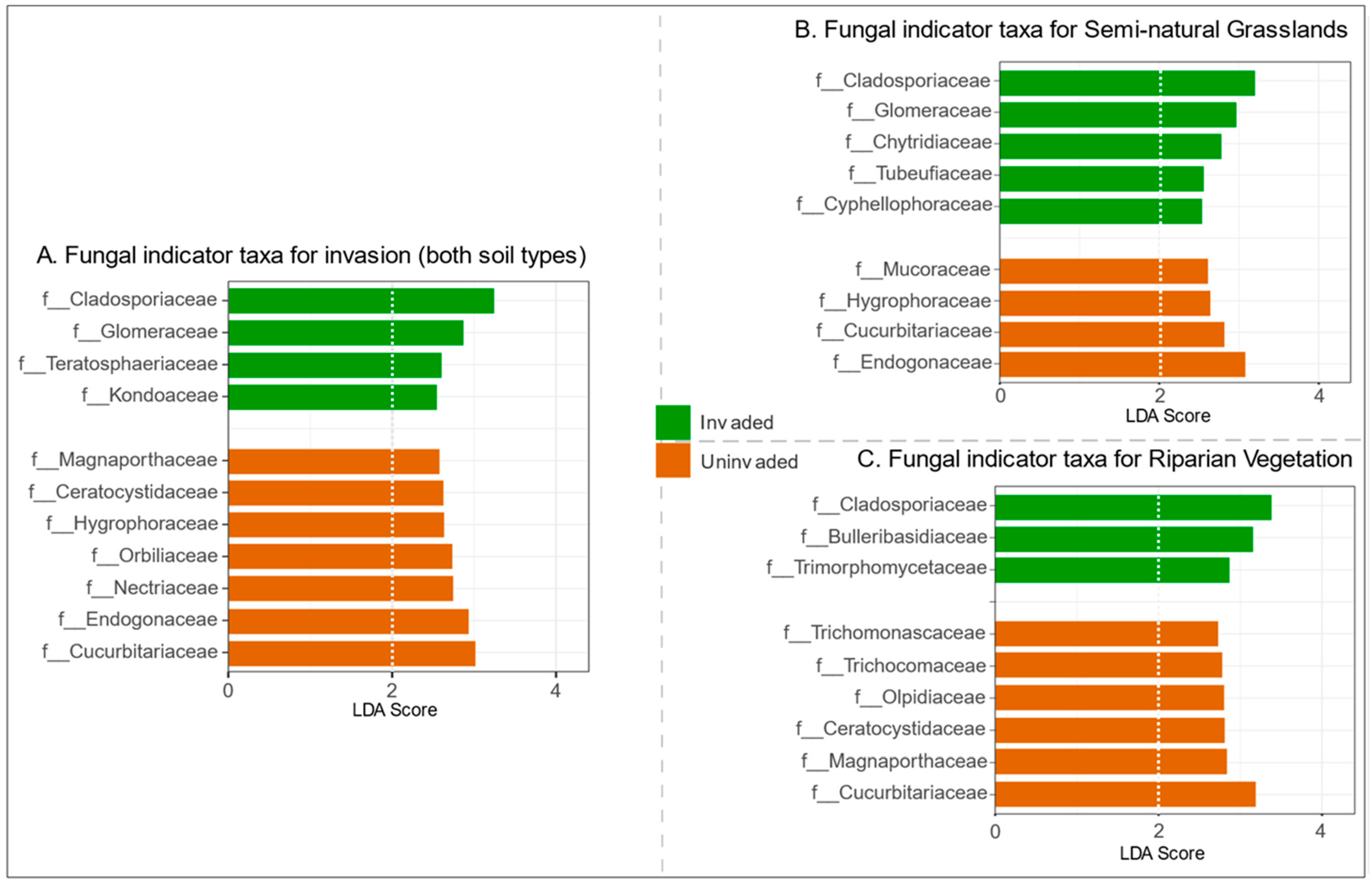
3.4. Fungal Indicator Taxa Associated with Invasive S. gigantea
4. Discussion
4.1. Apparent Discrepancy between Results from Independent Fungal Biomass Markers
4.2. The Habitat (In)Dependent Impact of S. gigantea on Fungivorous Nematode Lineages
4.3. Effect of Habitat-Characteristic Abiotic Differences between Habitat-Type
4.4. Fungal Indicator Taxa Related to Invasion with S. gigantea
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Pakpour, S.; Klironomos, J. The invasive plant, Brassica nigra, degrades local mycorrhizas across a wide geographical landscape. R. Soc. Open Sci. 2015, 2, 150300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, B.E.; Klironomos, J.N. Breaking new ground: Soil communities and exotic plant invasion. Bioscience 2005, 55, 477–487. [Google Scholar] [CrossRef]
- Dickie, I.A.; Bufford, J.L.; Cobb, R.C.; Desprez-Loustau, M.L.; Grelet, G.; Hulme, P.E.; Thrall, P.H. The emerging science of linked plant–fungal invasions. New Phytol. 2017, 215, 1314–1332. [Google Scholar] [CrossRef] [Green Version]
- Scharfy, D.; Güsewell, S.; Gessner, M.O.; Venterink, H.O. Invasion of Solidago gigantea in contrasting experimental plant communities: Effects on soil microbes, nutrients and plant–soil feedbacks. J. Ecol. 2010, 98, 1379–1388. [Google Scholar] [CrossRef]
- Weber, E.; Jakobs, G. Biological flora of central Europe: Solidago gigantea Aiton. Flora Morphol. Distrib. Funct. Ecol. Plants 2005, 200, 109–118. [Google Scholar] [CrossRef]
- Pal, R.W.; Chen, S.; Nagy, D.U.; Callaway, R.M. Impacts of Solidago gigantea on other species at home and away. Biol. Invasions 2015, 17, 3317–3325. [Google Scholar] [CrossRef]
- Vanderhoeven, S.; Dassonville, N.; Chapuis-Lardy, L.; Hayez, M.; Meerts, P. Impact of the invasive alien plant Solidago gigantea on primary productivity, plant nutrient content and soil mineral nutrient concentrations. Plant Soil 2006, 286, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Zubek, S.; Majewska, M.L.; Błaszkowski, J.; Stefanowicz, A.M.; Nobis, M.; Kapusta, P. Invasive plants affect arbuscular mycorrhizal fungi abundance and species richness as well as the performance of native plants grown in invaded soils. Biol. Fertil. Soils 2016, 52, 879–893. [Google Scholar] [CrossRef] [Green Version]
- Awaydul, A.; Zhu, W.; Yuan, Y.; Xiao, J.; Hu, H.; Chen, X.; Cheng, L. Common mycorrhizal networks influence the distribution of mineral nutrients between an invasive plant, Solidago canadensis, and a native plant, Kummerowa striata. Mycorrhiza 2019, 29, 29–38. [Google Scholar] [CrossRef]
- Hodge, A.; Campbell, C.D.; Fitter, A.H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atul-Nayyar, A.; Hamel, C.; Hanson, K.; Germida, J. The arbuscular mycorrhizal symbiosis links N mineralization to plant demand. Mycorrhiza 2009, 19, 239–246. [Google Scholar] [CrossRef]
- Cheng, L.; Booker, F.L.; Tu, C.; Burkey, K.O.; Zhou, L.; Shew, H.D.; Hu, S. Arbuscular mycorrhizal fungi increase organic carbon decomposition under elevated CO2. Science 2012, 337, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Nuccio, E.E.; Hodge, A.; Pett-Ridge, J.; Herman, D.J.; Weber, P.K.; Firestone, M.K. An arbuscular mycorrhizal fungus significantly modifies the soil bacterial community and nitrogen cycling during litter decomposition. Environ. Microbiol. 2013, 15, 1870–1881. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.A.; Reshi, Z.A.; Khasa, D.P. Arbuscular mycorrhizas: Drivers or passengers of alien plant invasion. Bot. Rev. 2009, 75, 397–417. [Google Scholar] [CrossRef]
- Yuan, Y.G.; Tang, J.J.; Leng, D.; Hu, S.J.; Yong, J.W.H.; Chen, X. An invasive plant promotes its arbuscular mycorrhizal symbioses and competitiveness through its secondary metabolites: Indirect evidence from activated carbon. PLoS ONE 2014, 9, e97163. [Google Scholar] [CrossRef] [PubMed]
- Quist, C.W.; Vervoort, M.T.W.; van Megen, H.; Gort, G.; Bakker, J.; van der Putten, W.H.; Helder, J. Selective alteration of soil food web components by invasive giant goldenrod Solidago gigantea in two distinct habitat types. Oikos 2014, 123, 837–845. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Stanek, M.; Nobis, M.; Zubek, S. Species-specific effects of plant invasions on activity, biomass, and composition of soil microbial communities. Biol. Fertil. Soils 2016, 52, 841–852. [Google Scholar] [CrossRef] [Green Version]
- Weete, J.D.; Abril, M.; Blackwell, M. Phylogenetic distribution of fungal sterols. PLoS ONE 2010, 5, e10899. [Google Scholar] [CrossRef] [Green Version]
- Holterman, M.H.M.; Karegar, A.; Mooijman, P.J.W.; van Megen, H.; van den Elsen, S.J.J.; Vervoort, M.T.W.; Quist, C.W.; Karssen, G.; Decraemer, W.; Opperman, C.H.; et al. Disparate gain and loss of parasitic abilities among nematode lineages. PLoS ONE 2017, 12, e185445. [Google Scholar] [CrossRef] [PubMed]
- Baynes, M.A.; Russell, D.M.; Newcombe, G.; Carta, L.K.; Rossman, A.Y.; Ismaiel, A. A mutualistic interaction between a fungivorous nematode and a fungus within the endophytic community of Bromus tectorum. Fungal Ecol. 2012, 5, 610–623. [Google Scholar] [CrossRef]
- Okada, H.; Kadota, I. Host status of 10 fungal isolates for two nematode species, Filenchus misellus and Aphelenchus avenae. Soil Biol. Biochem. 2003, 35, 1601–1607. [Google Scholar] [CrossRef]
- Barkman, J.J.; Doing, H.; Segal, S. Kritische bemerkungen und vorschläge zur quantitativen Vegetationsanalyse. Acta Bot. Neerl. 1964, 13, 394–419. [Google Scholar] [CrossRef]
- Leps, J.; Hadincova, V. How reliable are our vegetation analyses. J. Veg. Sci. 1992, 3, 119–124. [Google Scholar] [CrossRef]
- Oostenbrink, M. Estimating nematode populations by some selected methods. Nematology 1960, 6, 85–102. [Google Scholar]
- Vervoort, M.T.W.; Vonk, J.A.; Mooijman, P.J.W.; van den Elsen, S.J.J.; van Megen, H.H.B.; Veenhuizen, P.; Landeweert, R.; Bakker, J.; Mulder, C.; Helder, J. SSU ribosomal DNA-based monitoring of nematode assemblages reveals distinct seasonal fluctuations within evolutionary heterogeneous feeding guilds. PLoS ONE 2012, 7, e47555. [Google Scholar] [CrossRef]
- Harkes, P.; Verhoeven, A.; Sterken, M.G.; Snoek, L.B.; van den Elsen, S.J.J.; Mooijman, P.J.W.; Quist, C.W.; Vervoort, M.T.W.; Helder, J. The differential impact of a native and a non-native ragwort species (Senecioneae) on the first and second trophic level of the rhizosphere food web. Oikos 2017, 126, 1790–1803. [Google Scholar] [CrossRef]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal identification using molecular tools: A primer for the natural products research community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef]
- Bååth, E. Estimation of fungal growth rates in soil using C-14-acetate incorporation into ergosterol. Soil Biol. Biochem. 2001, 33, 2011–2018. [Google Scholar] [CrossRef]
- de Ridder-Duine, A.S.; Smant, W.; van der Wal, A.; van Veen, J.A.; de Boer, W. Evaluation of a simple, non-alkaline extraction protocol to quantify soil ergosterol. Pedobiologia 2006, 50, 293–300. [Google Scholar] [CrossRef]
- Harkes, P.; Suleiman, A.K.A.; van den Elsen, S.J.J.; de Haan, J.J.; Holterman, M.H.M.; Kuramae, E.E.; Helder, J. Conventional and organic soil management as divergent drivers of resident and active fractions of major soil food web constituents. Sci. Rep. 2019, 9, 13521. [Google Scholar] [CrossRef]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Schabenberger, O. SAS for Mixed Models, 2nd ed.; SAS Institute: Cary, NC, USA, 2006. [Google Scholar]
- Bushnell, B. BBMap: Short Read Aligner and Other Bioinformatic Tools V36.99. 2018. Available online: https://sourceforge.net/projects/bbmap/ (accessed on 17 February 2021).
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.T.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. Microbiome Analyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Barille-Boyer, A.L.; Barille, L.; Massé, H.; Razet, D.; Heral, M. Correction for particulate organic matter as estimated by loss on ignition in estuarine ecosystems. Estuar. Coast. Shelf Sci. 2003, 58, 147–153. [Google Scholar] [CrossRef]
- Lofgren, L.A.; Uehling, J.K.; Branco, S.; Bruns, T.D.; Martin, F.; Kennedy, P.G. Genome-based estimates of fungal rDNA copy number variation across phylogenetic scales and ecological lifestyles. Mol. Ecol. 2019, 28, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, H.J.; Monreal, C.M.; Young, J.C.; Seifert, K.A. Determination of soil fungal biomass from soil ergosterol analyses. Soil Biol. Biochem. 2000, 32, 1207–1217. [Google Scholar] [CrossRef]
- Hannula, S.E.; Morrien, E.; de Hollander, M.; van der Putten, W.H.; van Veen, J.A.; de Boer, W. Shifts in rhizosphere fungal community during secondary succession following abandonment from agriculture. ISME J. 2017, 11, 2294–2304. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Díaz-Ravinã, M.; Ritz, K.; McNicol, J.W.; Ebblewhite, N.; Bååth, E. Community DNA hybridisation and %G+C profiles of microbial communities from heavy metal polluted soils. FEMS Microbiol. Ecol. 1997, 24, 103–112. [Google Scholar] [CrossRef]
- Roper, M.; Ellison, C.; Taylor, J.W.; Glass, N.L. Nuclear and genome dynamics in multinucleate ascomycete fungi. Curr. Biol. 2011, 21, R786–R793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čerevková, A.; Miklisová, D.; Bobuľská, L.; Renčo, M. Impact of the invasive plant Solidago gigantea on soil nematodes in a semi-natural grassland and a temperate broadleaved mixed forest. J. Helminthol. 2019, 94, e51. [Google Scholar] [CrossRef]
- Li, Y.; Tsay, T.; Chen, P. Using three fungivorous nematodes to control lettuce damping-off disease caused by Rhizoctonia solani (AG4). Plant Pathol. Bull. 2014, 23, 43–53. [Google Scholar]
- Ruess, L.; Zapata, E.J.G.; Dighton, J. Food preferences of a fungal-feeding Aphelenchoides species. Nematology 2000, 2, 223–230. [Google Scholar] [CrossRef]
- Rousk, J.; Baath, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, N.F.; Liu, H.Y.; Zhang, Y.Q.; Yu, L.Y. Soil pH is a key determinant of soil fungal community composition in the Ny-Ålesund Region, Svalbard (High Arctic). Front. Microbiol. 2016, 7, 227. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Denis, M.; Gene, J.; Sutton, D.A.; Wiederhold, N.P.; Cano-Lira, J.F.; Guarro, J. New species of Cladosporium associated with human and animal infections. Persoonia 2016, 36, 281–298. [Google Scholar] [CrossRef] [Green Version]
- Hamayun, M.; Khan, S.A.; Khan, A.L.; Rehman, G.; Kim, Y.H.; Iqbal, I.; Hussain, J.; Sohn, E.-Y.; Lee, I.-J. Gibberellin production and plant growth promotion from pure cultures of Cladosporium sp MH-6 isolated from cucumber (Cucumis sativus L.). Mycologia 2010, 102, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Paul, D.; Park, K.S. Identification of volatiles produced by Cladosporium cladosporioides CL-1, a fungal biocontrol agent that promotes plant growth. Sensors 2013, 13, 13969–13977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirozynski, K.A.; Dalpe, Y. Geological history of the Glomaceae with particular reference to mycorrhizal symbiosis. Symbiosis 1989, 7, 1–36. [Google Scholar]
- Vallino, M.; Massa, N.; Lumini, E.; Bianciotto, V.; Berta, G.; Bonfante, P. Assessment of arbuscular mycorrhizal fungal diversity in roots of Solidago gigantea8 growing in a polluted soil in Northern Italy. Environ. Microbiol. 2006, 8, 971–983. [Google Scholar] [CrossRef]
- Treseder, K.K.; Allen, M.F. Mycorrhizal fungi have a potential role in soil carbon storage under elevated CO2 and nitrogen deposition. New Phytol. 2000, 147, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.G.; Miller, R.M. Carbon cycling by arbuscular mycorrhizal fungi in soil—Plant systems. Trends Plant Sci. 2003, 8, 407–409. [Google Scholar] [CrossRef]
- Zhou, J.; Zang, H.; Loeppmann, S.; Gube, M.; Kuzyakov, Y.; Pausch, J. Arbuscular mycorrhiza enhances rhizodeposition and reduces the rhizosphere priming effect on the decomposition of soil organic matter. Soil Biol. Biochem. 2020, 140, 107641. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Checa, J.; Blanco, M.N.; Olariaga, I.; Tello, S.; Voglmayr, H. A preliminary account of the Cucurbitariaceae. Stud. Mycol. 2018, 90, 71–118. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Habitat Type | Study Site | Soil Type | Coordinates | Year of S. gigantea Introduction | Number of Plots Pairs |
|---|---|---|---|---|---|
| Riparian zone | Millingerwaard | River clay | 51°51′58.11″ N 6°00′35.47″ E | ~1950 | 8 |
| Ewijkse plaat | River clay | 51°52′47.36″ N 5°44′52.17″ E | ~1950 | 8 | |
| Blauwe Kamer | |||||
| West | River clay | 51°56′40.22″ N 5°36′19.90″ E | after 1950 | 4 | |
| East | River clay and sand | 51°56′32.56″ N 5°37′09.54″ E | after 1950 | 4 | |
| Semi-natural grassland | Dennenkamp | Pleistocene sand | 52°01′45.64″ N 5°47′53.50″ E | 1982 | 8 |
| Plantage Willem III | Pleistocene sand | 51°58′48.62″ N 5°31′08.47″ E | 1995 | 8 | |
| Hollandseweg | Pleistocene sand | 51°58′49.89″ N 5°40′59.84″ E | before 2005 | 4 | |
| Scheidingslaan | Pleistocene sand | 51°58′28.60″ N 5°41′55.40″ E | unknown | 4 | |
| Reijerscamp | Pleistocene sand | 52°00′47.49″ N 5°46′08.64″ E | 2006 | 4 |
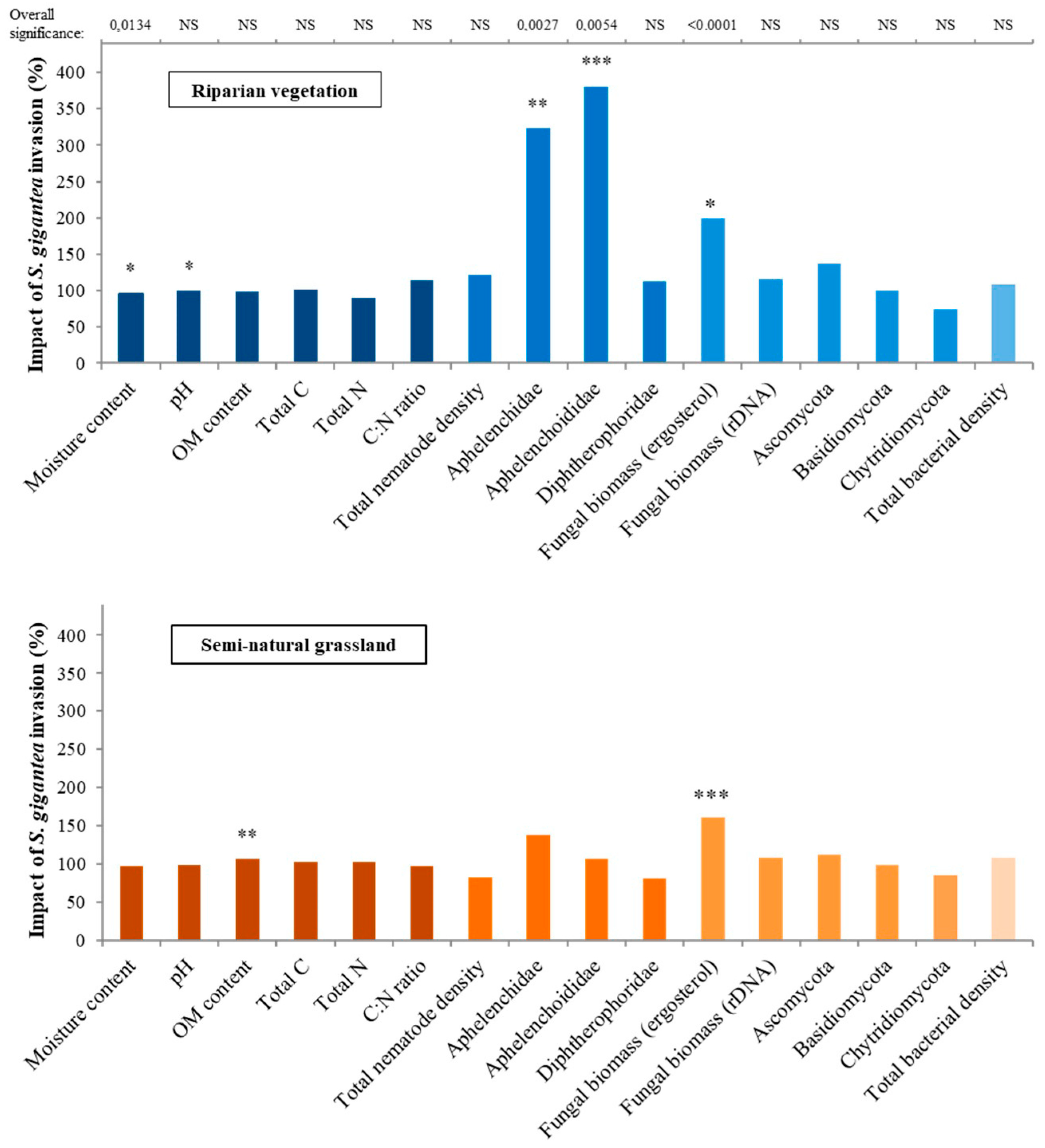
| Habitat Type | Invasion | Habitat Type * Invasion | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Abiotic Variables | Fdf | p-Value | Fdf | p-Value | Fdf | p-Value | |||
| Moisture content (%) | F1,6 | 2.02 | 0.2052 | F1,50 | 6.58 | 0.0134 | F1,50 | 0.93 | 0.3391 |
| pH | F1,6 | 92.22 | <0.0001 | F1,50 | 2.48 | 0.1213 | F1,50 | 0.19 | 0.6661 |
| Organic matter (OM) content (%) | F1,6 | 0.13 | 0.7343 | F1,50 | 2.87 | 0.0965 | F1,50 | 4.74 | 0.0341 |
| Total C (g/kg) | F1,6 | 1.38 | 0.2848 | F1,6 | 0.05 | 0.8281 | F1,6 | 0.00 | 0.9562 |
| Total N (g/kg) | F1,6 | 0.57 | 0.4803 | F1,6 | 0.15 | 0.7090 | F1,6 | 0.60 | 0.4692 |
| C:N ratio | F1,6 | 0.73 | 0.4267 | F1,6 | 0.43 | 0.5378 | F1,6 | 1.00 | 0.3568 |
| Biotic variables | |||||||||
| Total nematode density (#) | F1,6 | 26.69 | 0.0021 | F1,50 | 0.00 | 0.9758 | F1,50 | 3.24 | 0.0780 |
| Aphelenchidae (#) | F1,6 | 0.02 | 0.8962 | F1,50 | 9.96 | 0.0027 | F1,50 | 3.16 | 0.0814 |
| Aphelenchoididae (#) | F1,6 | 0.65 | 0.4507 | F1,50 | 8.44 | 0.0054 | F1,50 | 6.92 | 0.0113 |
| Diphtherophora (#) | F1,4 | 0.08 | 0.7930 | F1,42 | 0.02 | 0.9007 | F1,42 | 0.29 | 0.5945 |
| Fungal biomass (mg erg/kg) | F1,6 | 3.63 | 0.1055 | F1,48 | 21.97 | <0.0001 | F1,48 | 0.72 | 0.3990 |
| Fungal DNA (μg) | F1,6 | 0.75 | 0.4184 | F1,50 | 2.50 | 0.1203 | F1,50 | 0.17 | 0.6846 |
| Ascomycota (μg) | F1,6 | 0.06 | 0.8187 | F1,50 | 3.34 | 0.0738 | F1,50 | 0.71 | 0.4027 |
| Basidiomycota (μg) | F1,6 | 10.83 | 0.0166 | F1,50 | 0.00 | 0.9637 | F1,50 | 0.01 | 0.9370 |
| Chytridiomycota (μg) | F1,6 | 1.07 | 0.3405 | F1,50 | 1.25 | 0.2693 | F1,50 | 0.12 | 0.7340 |
| Total bacterial density (μg) | F1,6 | 0.05 | 0.8360 | F1,50 | 3.29 | 0.0759 | F1,50 | 0.02 | 0.8826 |
| Riparian Vegetation (n = 24 Plot-Pairs) | Semi-Natural Grassland (n = 28 Plot-Pairs) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Invaded (n = 24) | Uninvaded (n = 24) | Invaded (n = 28) | Uninvaded (n = 28) | |||||||||||
| Lower | Est. mean | Upper | Lower | Est. mean | Upper | p-Value | Lower | Est. mean | Upper | Lower | Est. mean | Upper | p-Value | |
| Abiotic variables | ||||||||||||||
| Moisture content (%) | 15.9 | 20.7 | 25.6 | 16.7 | 21.5 | 26.4 | 0.0199 | 12.8 | 16.6 | 20.4 | 13.2 | 17.0 | 20.7 | 0.2445 |
| pH | 7.33 | 7.46 | 7.59 | 7.37 | 7.50 | 7.63 | 0.0197 | 5.21 | 5.58 | 5.96 | 5.28 | 5.65 | 6.02 | 0.3059 |
| OM content (%) | 4.2 | 5.9 | 8.3 | 4.2 | 5.9 | 8.4 | 0.7429 | 4.3 | 5.7 | 7.4 | 4.0 | 5.3 | 6.9 | 0.0063 |
| Total C (g/kg) | 17.0 | 31.2 | 57.0 | 16.8 | 30.6 | 56.0 | 0.9181 | 13.9 | 22.1 | 35.3 | 13.5 | 21.5 | 34.4 | 0.8243 |
| Total N (g/kg) | 0.9 | 1.7 | 3.1 | 1.0 | 1.9 | 3.4 | 0.4895 | 0.9 | 1.5 | 2.3 | 0.9 | 1.4 | 2.2 | 0.7663 |
| C:N ratio | 12.9 | 19.1 | 25.2 | 10.6 | 16.8 | 22.9 | 0.3366 | 10.5 | 15.2 | 20.0 | 10.9 | 15.7 | 20.5 | 0.7879 |
| Biotic variables | ||||||||||||||
| Aphelenchidae (#) | 0.4 | 1.6 | 6.0 | 0.1 | 0.5 | 2.0 | 0.0015 | 0.4 | 1.2 | 3.6 | 0.3 | 0.9 | 2.7 | 0.3156 |
| Aphelenchoididae (#) | 1.4 | 4.3 | 13.0 | 0.3 | 1.1 | 3.6 | 0.0004 | 1.6 | 4.0 | 9.8 | 1.5 | 3.7 | 9.2 | 0.8404 |
| Diphtherophora (#) | 0.4 | 0.9 | 1.9 | 0.4 | 0.8 | 1.7 | 0.7621 | 0.4 | 0.9 | 2.0 | 0.5 | 1.1 | 2.4 | 0.6564 |
| Fungal biomass (mg erg/kg) | 1.07 | 1.75 | 2.85 | 0.54 | 0.88 | 1.43 | 0.0199 | 1.74 | 2.66 | 3.97 | 1.22 | 1.65 | 2.76 | <0.0001 |
| Fungal DNA (μg) | 220 | 299 | 405 | 191 | 259 | 351 | 0.1814 | 264 | 340 | 438 | 243 | 313 | 403 | 0.3925 |
| Ascomycota (μg) | 26.1 | 40.2 | 61.9 | 19.1 | 29.3 | 45.2 | 0.0748 | 26.9 | 38.7 | 55.6 | 24.0 | 34.5 | 49.5 | 0.4730 |
| Basidiomycota (μg) | 1.6 | 2.8 | 5.1 | 1.6 | 2.8 | 5.0 | 0.9818 | 5.4 | 8.8 | 14.3 | 5.5 | 9.0 | 14.5 | 0.9270 |
| Chytridiomycota (μg) | 0.4 | 0.9 | 1.8 | 0.6 | 1.2 | 2.4 | 0.3250 | 0.3 | 0.6 | 1.2 | 0.4 | 0.7 | 1.4 | 0.5709 |
| Total bacterial density (μg) | 3598 | 4929 | 6754 | 3303 | 4526 | 6201 | 0.1875 | 3654 | 4696 | 6034 | 3399 | 4367 | 5612 | 0.2263 |
| Main Effect | Riparian Vegetation | Semi-Natural Grassland | ||||
|---|---|---|---|---|---|---|
| R2 | p | R2 | p | R2 | p | |
| Habitat Type | 0.274 | 0.0001 | ||||
| Study Site | 0.227 | 0.0001 | 0.257 | 0.001 | 0.356 | 0.001 |
| Invasion | 0.017 | 0.005 | 0.037 | 0.010 | 0.033 | 0.001 |
| Study Site * Invasion | 0.030 | 0.463 | 0.030 | 0.532 | 0.051 | 0.327 |
| Habitat * Invasion | 0.009 | 0.082 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harkes, P.; van Heumen, L.J.M.; van den Elsen, S.J.J.; Mooijman, P.J.W.; Vervoort, M.T.W.; Gort, G.; Holterman, M.H.M.; van Steenbrugge, J.J.M.; Quist, C.W.; Helder, J. Characterization of the Habitat- and Season-Independent Increase in Fungal Biomass Induced by the Invasive Giant Goldenrod and Its Impact on the Fungivorous Nematode Community. Microorganisms 2021, 9, 437. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020437
Harkes P, van Heumen LJM, van den Elsen SJJ, Mooijman PJW, Vervoort MTW, Gort G, Holterman MHM, van Steenbrugge JJM, Quist CW, Helder J. Characterization of the Habitat- and Season-Independent Increase in Fungal Biomass Induced by the Invasive Giant Goldenrod and Its Impact on the Fungivorous Nematode Community. Microorganisms. 2021; 9(2):437. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020437
Chicago/Turabian StyleHarkes, Paula, Lisa J. M. van Heumen, Sven J. J. van den Elsen, Paul J. W. Mooijman, Mariëtte T. W. Vervoort, Gerrit Gort, Martijn H. M. Holterman, Joris J. M. van Steenbrugge, Casper W. Quist, and Johannes Helder. 2021. "Characterization of the Habitat- and Season-Independent Increase in Fungal Biomass Induced by the Invasive Giant Goldenrod and Its Impact on the Fungivorous Nematode Community" Microorganisms 9, no. 2: 437. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020437