1. Introduction
Salmonella enterica is a foodborne pathogen that causes an estimated 1.2 million human infections annually in the United States [
1]. There are more than 2600
Salmonella enterica serotypes which can infect a wide range of vertebrate species, but fewer than 100 serotypes cause the majority of human infections [
2,
3]. Infected poultry may be asymptomatic of clinical disease but continue to shed zoonotic subspecies into the environment causing gastroenteritis and systemic infections in humans [
4,
5].
Salmonella enterica Typhimurium (ST), Kentucky (SK), and Enteritidis (SE) are in the top 5 common isolates found in contaminated chicken [
4,
6,
7]. Reducing
Salmonella colonization of poultry would benefit agricultural and public health sectors by decreasing medical costs and lowering cases of human infections.
In 2014, the US Department of Agriculture (USDA) Food Safety and Inspection Service determined that SK, SE, and ST were isolated from 60.8%, 13.6%, and 7.7% of young chicken carcasses, respectively [
3].
Salmonella enterica serotype Kentucky (SK) is the most prevalent serotype isolated from contaminated poultry carcasses; however, only 0.14% of human clinical disease was reported with this bacterium [
8]. In 2016, SE and ST were confirmed from 16.8% and 9.8% human Salmonellosis cases, respectively [
8]. These
Salmonella are prevalent in poultry; however, only SE and ST are considered a major food safety concern when compared to SK [
3].
Neonatal chicks are more susceptible to foreign bacteria, such as
Salmonella, due to their lack of mature intestinal microbiota [
9]. Initial invasion in the gastrointestinal tract leads to increased expression of chemokines, cytokines and an influx of heterophils and macrophages [
10,
11,
12].
Salmonella Typhimurium infection significantly decreased jejunum villus height from the host’s inflammatory response due to the influx of heterophils in one-day-old chicks [
13,
14].
Salmonella can hide, multiply and survive in macrophages leading to persistence [
15]. Disease tolerance occurs as
Salmonella is then able to persist in the gut of chickens without severe clinical signs [
16].
Intestinal direct colonization resistance is the inability of foreign ingested bacteria to colonize due to host bacteria [
17]. Mechanisms can include nutrient competition or active antagonism, such as bacteriocins [
18,
19]. Rantala and Nurmi used mature chicken intestinal bacteria to reduce colonization of
Salmonella Infantis in chicks [
20]. Oral administration of attenuated ST given to one-day-old chicks has been shown to competitively exclude future colonization of intestinal ST when it is again administered 24 h later [
21]. Methner and colleagues challenged chicks with various
Salmonella serotypes and found greater inhibition occurred between isogenic strains [
22]. Yang and colleagues determined intra-genus competitive exclusion occurs between ST and SE when administered 24 h apart in neonatal broilers [
23]. All are examples of ways colonization resistance can be used to exploit competitive exclusion in order to reduce
Salmonella colonization in chicks.
Indirect colonization resistance occurs through microbiota-stimulated host immunity and immune cell interactions. These include an enhanced mucosal barrier by production of mucus, short-chain fatty acids, such as butyrate or acetate, and host antimicrobial peptides from resident Paneth cells [
24]. Characterization of a broiler’s immune response during a
Salmonella infection can be measured by cytokine gene expression. An early localized inflammatory response includes an influx of heterophils and macrophages [
25]. Interleukin-1β (IL-1β) is a pro-inflammatory mediator and expression is increased in response to bacterial, viral, and parasitic infections [
26]. The expression of other pro-inflammatory cytokines, such as IL-6 enhance protection during ST infection by inducing acute phase protein synthesis and are important in further stimulating a T
H1 host immune response [
27,
28]. Interleukin-18 is a member of the IL-1 family and is produced by macrophages in response to lipopolysaccharide [
29,
30]. When IL-18 is in the presence of IL-12, gamma interferon (IFN-γ) production is upregulated in T
H1 and NK cells [
30]. Chicken IFN-γ is a macrophage-activating factor and is crucial in response to intracellular pathogens by inducing a cell-mediated T
H1 response [
31,
32,
33]. The function of IL-10 is to induce immunoregulatory effects, such as downregulating the production of pro-inflammatory cytokines [
34]. The cecal tonsils are composed of lymphoid follicles that contain multiple immune cells [
35]. The liver is important for immune function, containing Kupffer and other immune cells, synthesizing cytokines, chemokines, and acute-phase proteins in response to infection, trauma or stress [
36]. Acute phase proteins, such as mannan binding lectins, can activate phagocytes and modulate cytokine expression [
37]. Therefore, cytokine mRNA expression can be measured in chicken cecal tonsils and livers to measure the host immune interaction with
Salmonella as a local or systemic infection [
38]. The expression of cytokines can correlate with the presence of pathogens in the gut; however, mRNA expression may not always indicate actual protein synthesis [
38].
Salmonella Kentucky (SK) is the number one isolated serotype in poultry but causes a low incidence (0.14% cases) of clinical disease in humans [
8]. Fricke and colleagues screened
Salmonella enterica strains for avian pathogenic plasmid uptake from
Escherichia coli to screen for virulence evolution and host adaptations [
39]. Evidence of these plasmids was predominantly found in SK but not in other strains of
Salmonella, which could give an advantage to SK to cope with stress factors and competition [
39]. Cheng and colleagues showed SK persisted until the end of their challenge on day 36 compared to ST colonization which had fallen below the level of detection by day 15 [
40]. Prevalence of SK in the ceca, over other serotypes, could be further explained at the molecular level, such as increased transcription of regulatory protein, RNA polymerase (Rpos) [
40]. A sigma factor, such as Rpos, initiates transcription in stress response genes, so if it were elevated it would allow for greater proliferation of bacteria [
41]. Since SK is the predominantly isolated serotype, we believe this serotype may be used to exclude and reduce other
Salmonella as a live vaccine candidate.
We hypothesized that an initial infection by SK can reduce intra-genus serotypes SE and ST by competitive exclusion. Incidence of SK in the poultry industry is increasing [
39]. Understanding how these serotypes interact is necessary, but there is little information available. Furthermore, we attempted to characterize the host immune response by measuring cytokine gene expression in cecal tonsils and livers during a co-infection.
3. Results and Discussion
Colonization resistance is the inability of potentially pathogenic or foreign bacteria to expand due to host microbiota under homeostatic conditions [
55]. In trials 1 and 2, competitive exclusion between SK and ST was measured.
Salmonella was not detected in any of the negative control cultures in trials 1 and 2. As shown in
Table 4, ST was not recovered from the cecal contents in birds when SK was administered 1 day before in trial 1.
Salmonella Kentucky was significantly reduced when ST was administered prior (
Table 4).
Salmonella Typhimurium was significantly reduced when
Salmonella Kentucky was orally administered 24 h first and in combination with SK in trials 1 and 2 (
Table 4 and
Table 5).
Salmonella Kentucky was recovered from every bird challenged (
Table 4 and
Table 5).
Competitive exclusion of
Salmonella Enteritidis, another prevalent foodborne strain isolated from humans and poultry, and
Salmonella Kentucky was also compared in trials 3 and 4.
Salmonella was not detected in any of the negative control cultures in trials 1 and 2.
Salmonella Enteritidis was significantly reduced when
Salmonella Kentucky was administered first and when administered in combination with SK in trials 3 and 4 (
Table 6 and
Table 7).
Salmonella Kentucky was significantly reduced when challenged by SE 24 h later and when SE was administered first in Trial 3 (
Table 6).
Salmonella Kentucky and Enteritidis were recovered from all respective cecal enrichments from birds challenged (
Table 6 and
Table 7).
Research has shown that strains of
Salmonella can be used to exclude one another through competitive exclusion along with host defense colonization resistance mechanisms. When ST was administered to day-old chicks, it inhibited other
Salmonella strains from colonizing the alimentary tract [
56]. Yang and colleagues measured competitive exclusion in chicks using SE and ST and presented similar results [
23]. Similar to these findings, gnotobiotic pigs were protected from pathogenic ST when administered avirulent
Salmonella Infantis 24 h beforehand [
57]. As seen in this study, oral administration of
Salmonella reduces subsequent colonization of an isogenic isolate in neonatal broilers.
Heterophil populations increase in the chick’s cecal lamina propria from D2 to D4 post-infection with SE and ST [
13,
58]. An influx of avian host defense peptides includes gallinacins, cathelicidins and liver expressed antimicrobial peptides, which are upregulated during infection, with SE or ST [
59,
60,
61].
Salmonella enterica are facultative intracellular bacteria, which means they can persist in macrophages, travel through the bloodstream, and spread systemically [
13,
62,
63]. The incidence of SK in liver/spleen macerations was significantly higher in the SK + ST group compared to other treatments in trial 1 (
Table 4). A significantly higher incidence of SK and ST in liver/spleen macerations was measured in the ST → SK group compared to other treatments (
Table 5). In trial 3, there was a higher incidence of SK in the livers/spleens compared to the other treatments (
Table 6). Newly hatched chicks are more susceptible to systemic translocation of
Salmonella from the gut barrier due to an underdeveloped immune system, immature microflora, and a relatively sterile gut [
64,
65,
66,
67].
Interleukin-1β is a key mediator during inflammation and induces the production of chemokines, such as IL-8, to attract specific immune cells [
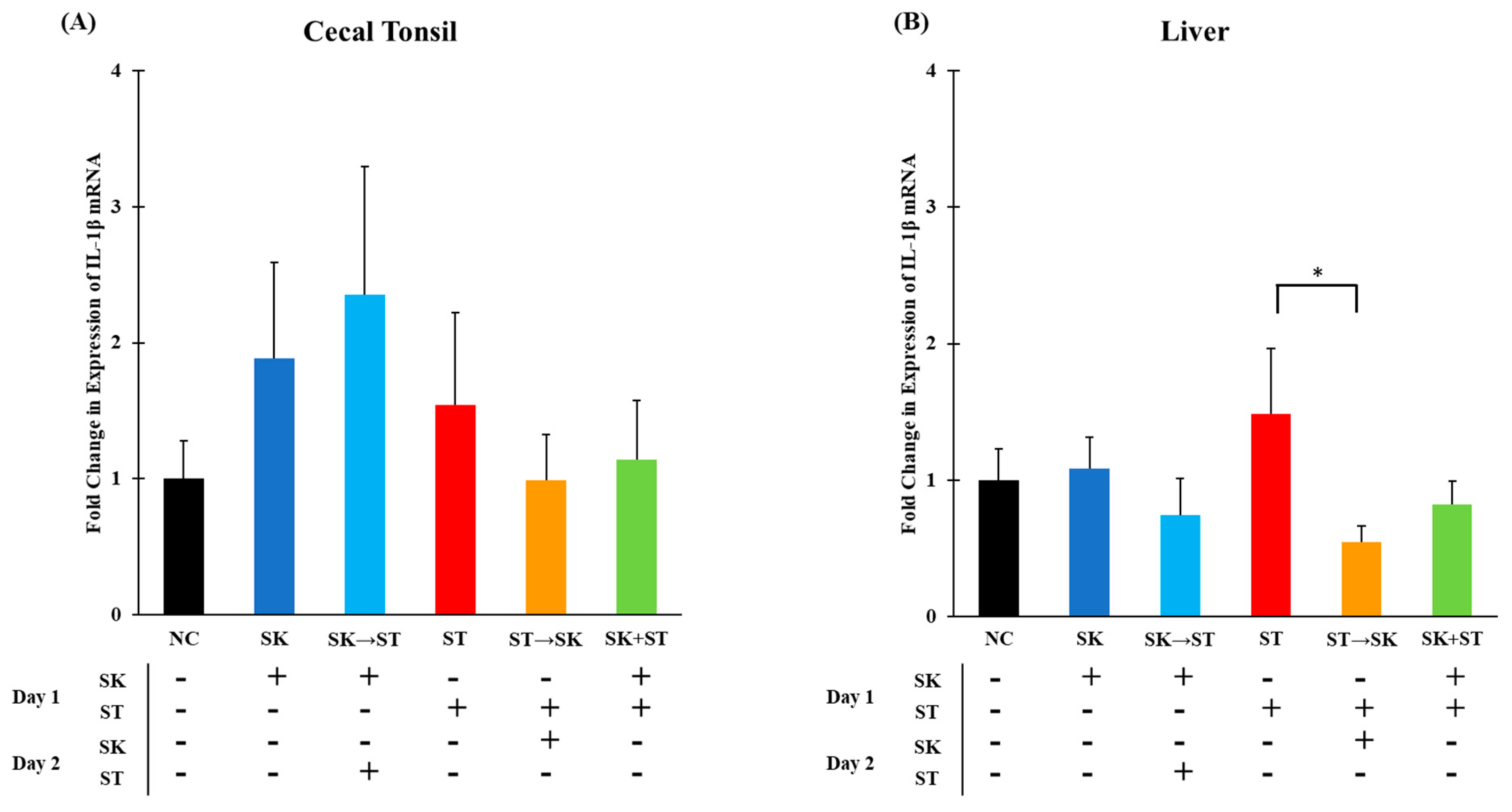
26]. In trial 1, the mRNA levels of IL-1β were not different in the cecal tonsils (
Figure 1A). The liver mRNA IL-1β levels were significantly higher in the ST treatment compared to the group given ST followed by SK 24 h later (
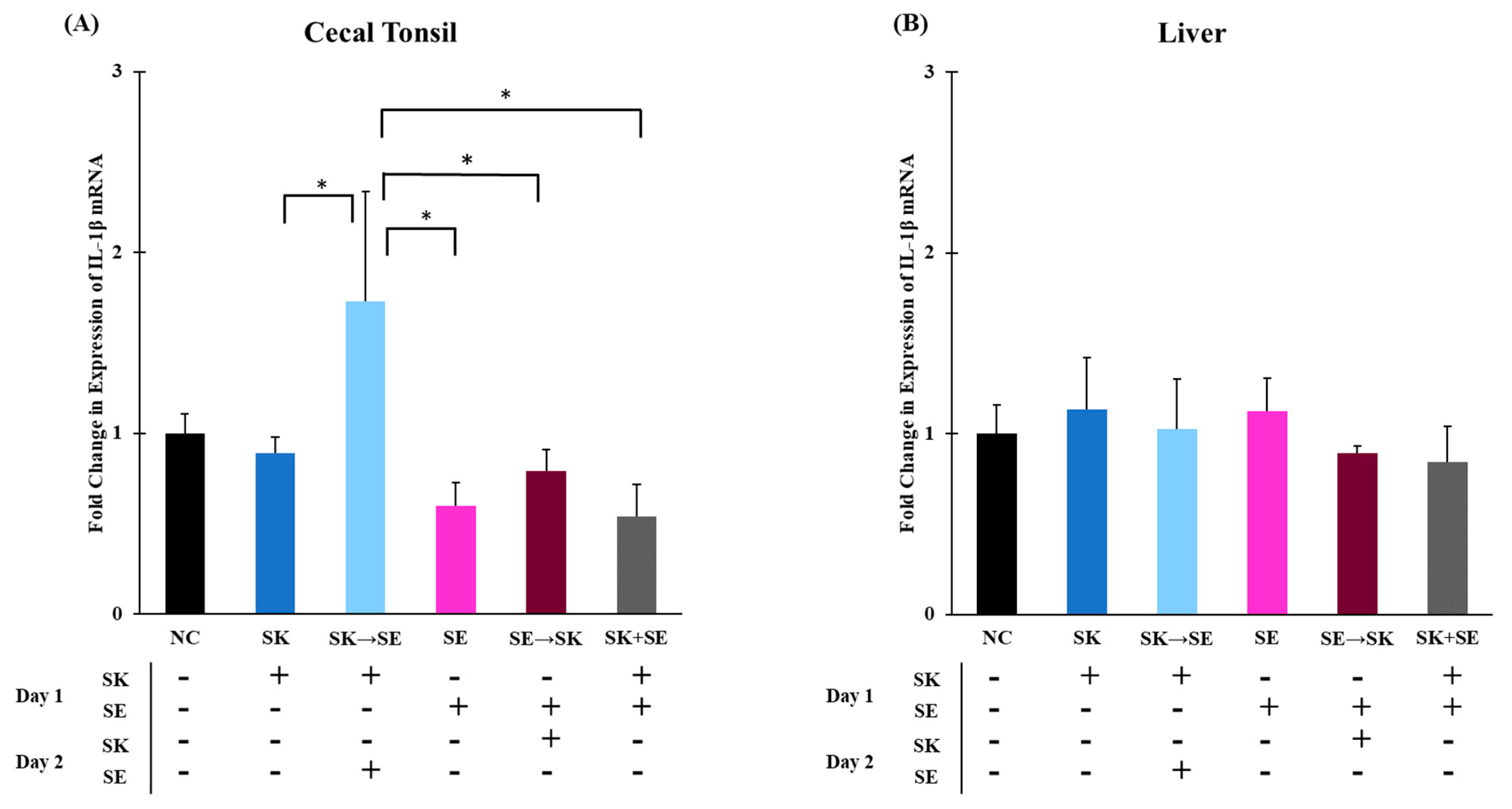
Figure 1B). In trial 3, the mRNA levels of IL-1β were significantly higher when SK was administered 24 h prior to SE than the treatments of SK, SE, SE followed by SK, and SK and SE combination in the cecal tonsils (
Figure 2A). The elevated expression could be due to the consecutive challenge. There were no changes in the liver (
Figure 2B). Withanage and colleagues measured a significant increase of IL-1β levels in cecal tonsils of day-old chicks 6–48 h post-infection when challenged with 1.0 × 10
8 CFU of
Salmonella Typhimurium [
38]. Fasina and colleagues found a significant upregulation in IL-1β mRNA expression in ceca of 5 and 10 days post challenged 4-day-old broilers that were gavaged with 7.8 × 10
6 CFU/mL of ST [
68]. Chranova and colleagues also measured an increase in IL-1β mRNA levels of day-old layer chick ceca when gavaged with 1.0 × 10
6 CFU of SE [
69]. However, changes in IL-1β mRNA levels were not found in the ceca of week-old chicks challenged with 10
8 CFU of ST in Withanage and colleagues’ earlier studies but was found in the liver 24 h post-infection [
11]. Different time point measurements and challenge amounts can affect mRNA expression levels which could be the reason why some of our levels contrast with the literature.
Another initial pro-inflammatory cytokine, IL-6, induces the synthesis of acute phase proteins, such as mannan binding lectin, from hepatic cells to initiate an innate immune response [
28,
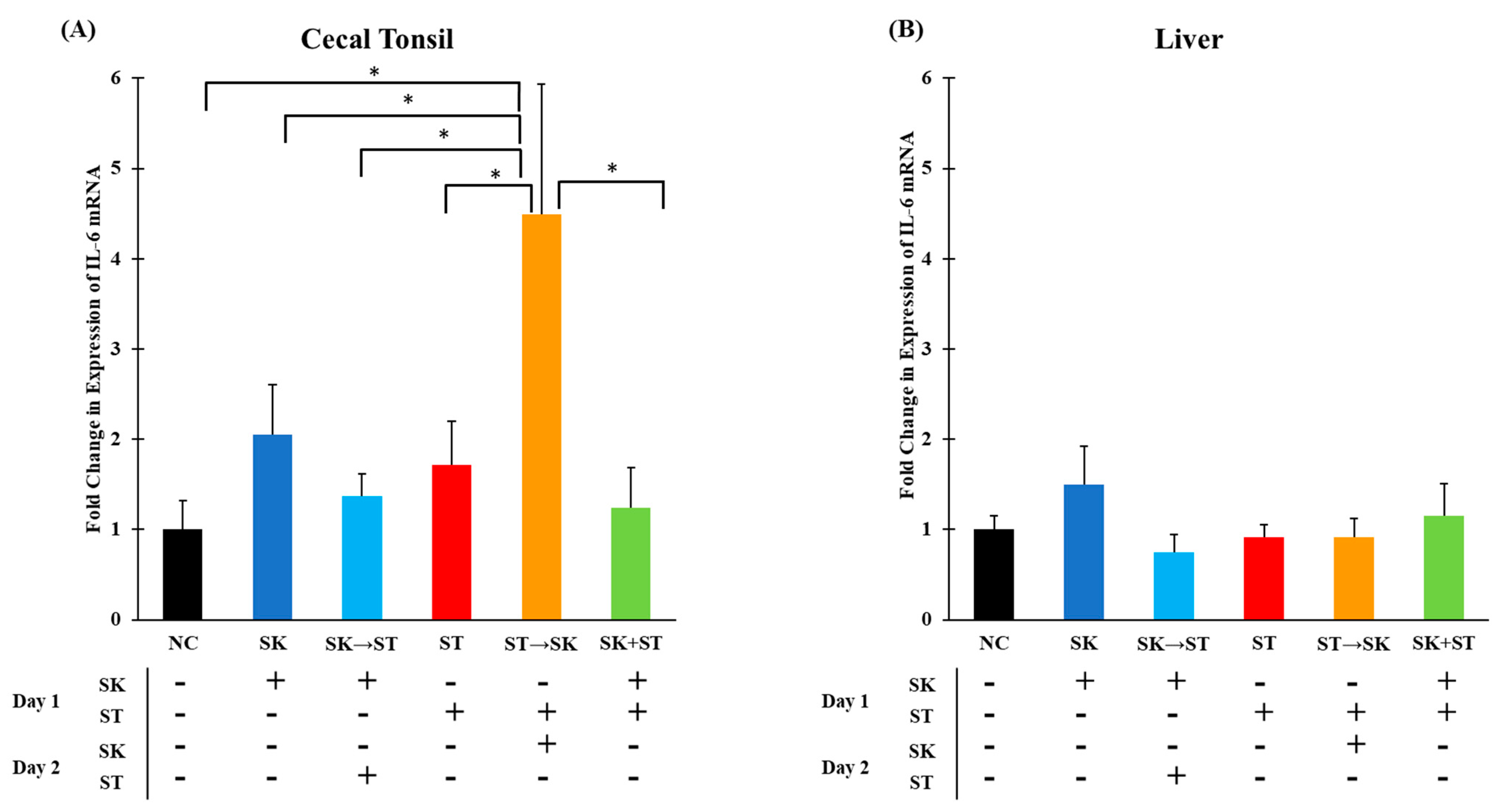
70]. The mRNA levels of IL-6 were significantly higher in the ST followed by the SK group compared to other treatments in cecal tonsils (
Figure 3A). An increase in levels could be explained by the consecutive challenge of these two serotypes. There were no significant differences in IL-6 mRNA expression in the liver among the treatment groups (
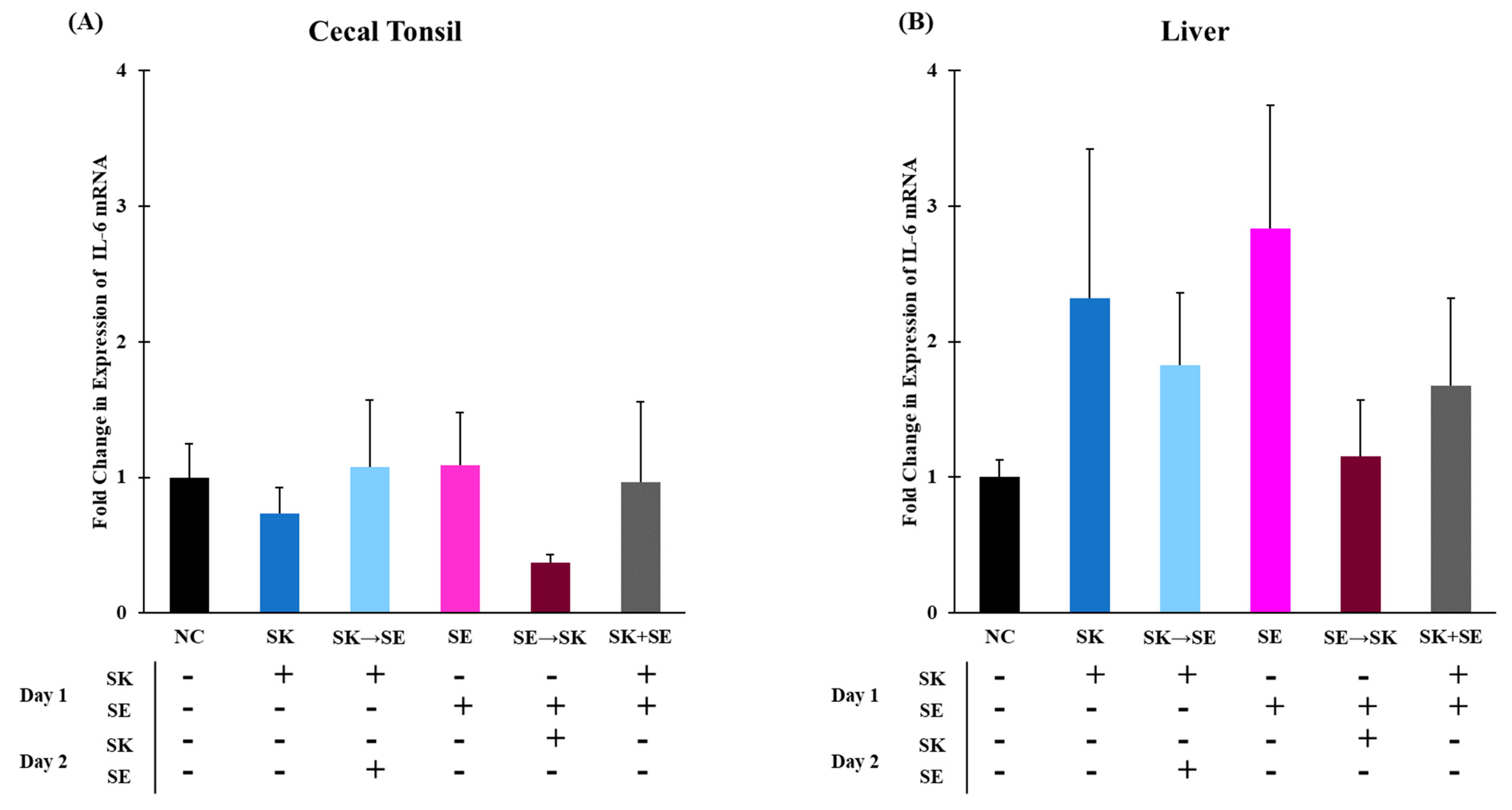
Figure 3B). There were no significant changes in IL-6 expression in the cecal tonsils and liver in trial 3 (
Figure 4A,B). Changes in IL-6 mRNA levels were not found in the ceca of day-old chicks challenged with 10
8 CFU of ST 6–48 h post-infection [
38]. Setta and colleagues also measured no changes in IL-6 mRNA levels in the ceca of one-day-old broilers challenged with 10
9 CFU of SE [
12]. In week-old chicks challenged with 10
8 cfu of ST, IL-6 was not significantly increased until 21 and 28 days post-infection in the ceca and liver [
11]. Millet and colleagues measured an acute phase response to ST lipopolysaccharide in whole blood [
71]. We did not observe significantly elevated levels of IL-6 expression in the liver; however, expression does not always correlate to protein function.
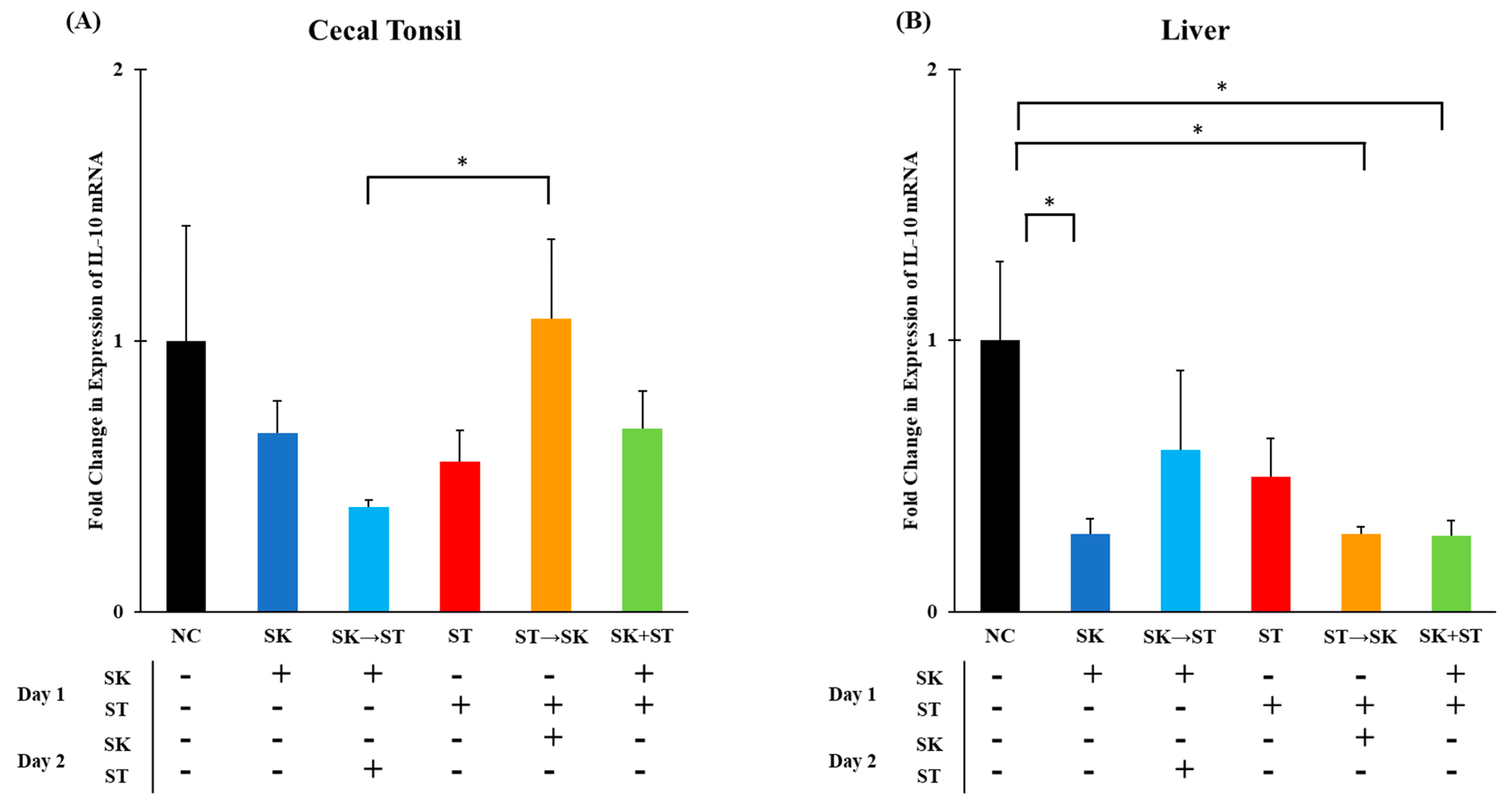
Regulatory cytokines, such as IL-10, inhibit the production and secretion of pro-inflammatory cytokines therefore suppressing a T
H1 response [
72,
73]. The mRNA levels of IL-10 were significantly higher in the ST followed by SK treatment than in the SK followed by ST treatment in the cecal tonsils (
Figure 5A). The levels of IL-10 were higher in the negative control group than the SK, ST followed by SK, and SK and ST combination treatments in the liver (
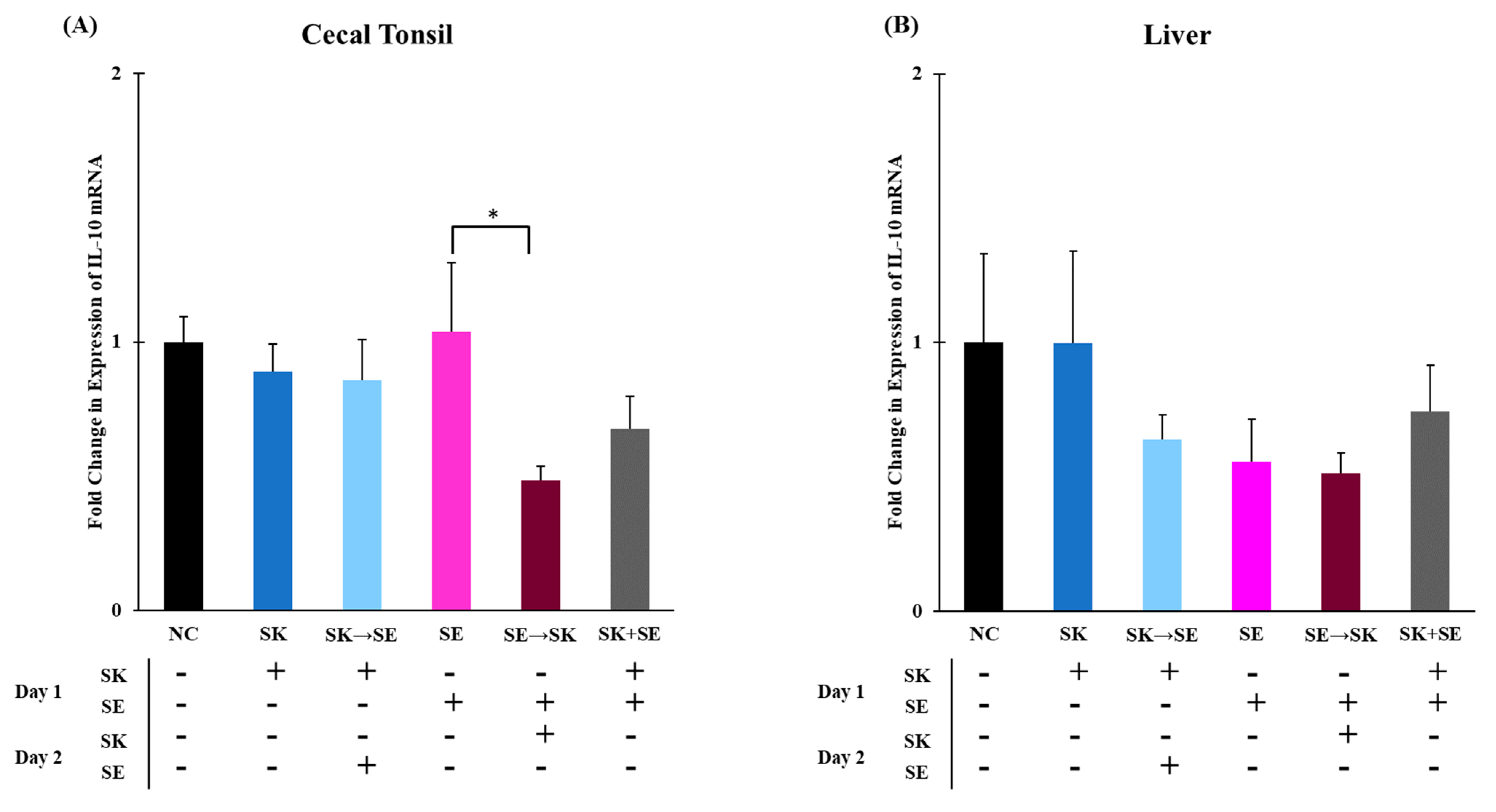
Figure 5B). The mRNA levels of IL-10 were significantly higher in the SE only treatment than the SE followed by SK treatment in the cecal tonsils (
Figure 6A). There were no changes in the liver (
Figure 6B). The IL-10 levels were downregulated 5 days post ST challenge in the ceca of broiler chicks in Fasina’s previously described experiment [
68]. Chranova and colleagues reported mRNA levels for IL-10 were significantly lower in the ceca of the previously mentioned experiment [
69].
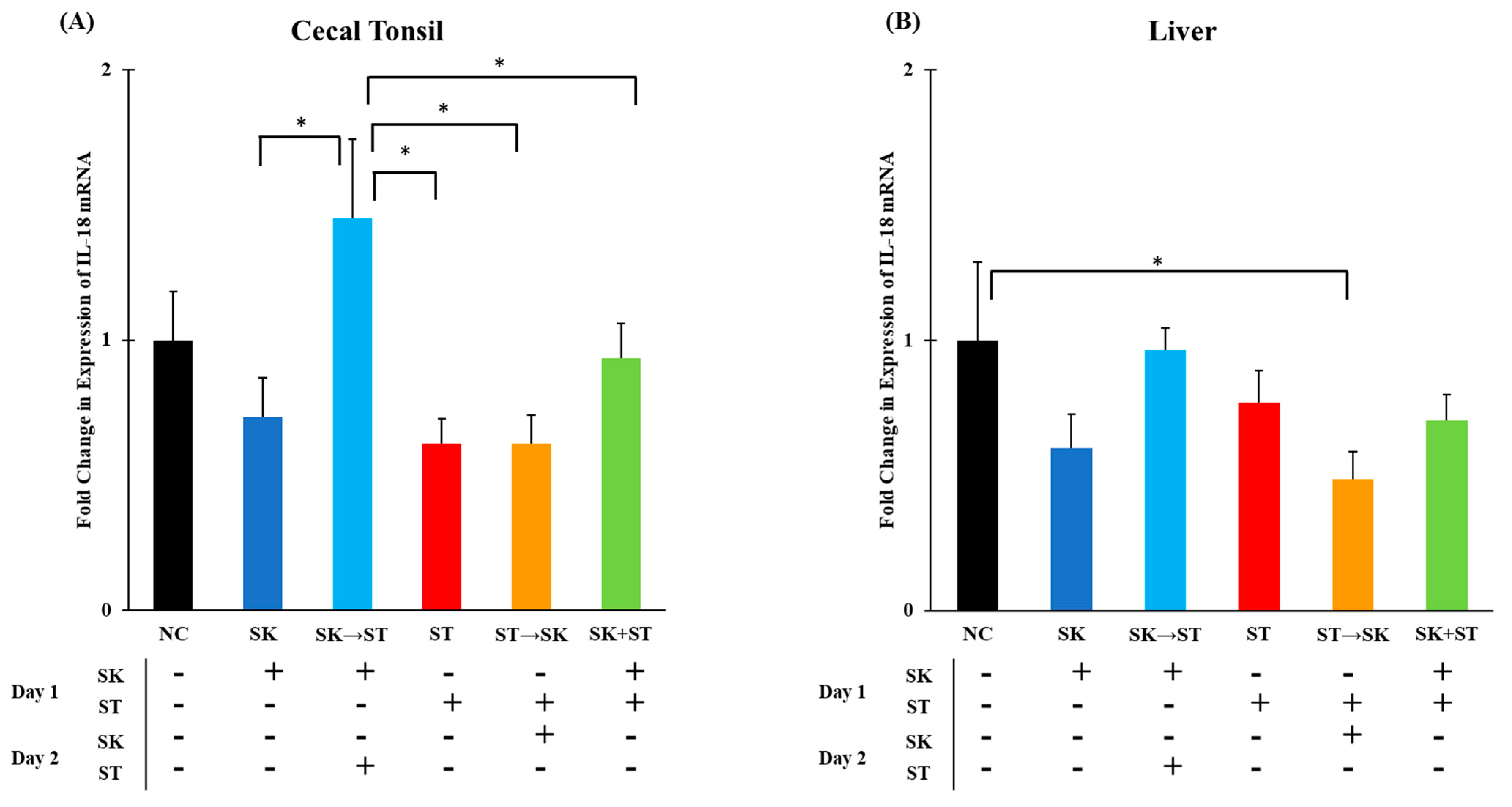
Chicken IL-18 is produced by macrophages and induces production of IFN-γ which further mediates T
H1 cell development [
30]. The mRNA levels of IL-18 were significantly higher in the SK followed by ST treatment than in the SK, ST, ST followed by SK, and SK and ST combination treatments in the cecal tonsils (
Figure 7A). Levels of IL-18 were significantly higher in the negative control group compared to the ST followed by SK treatment in the liver (
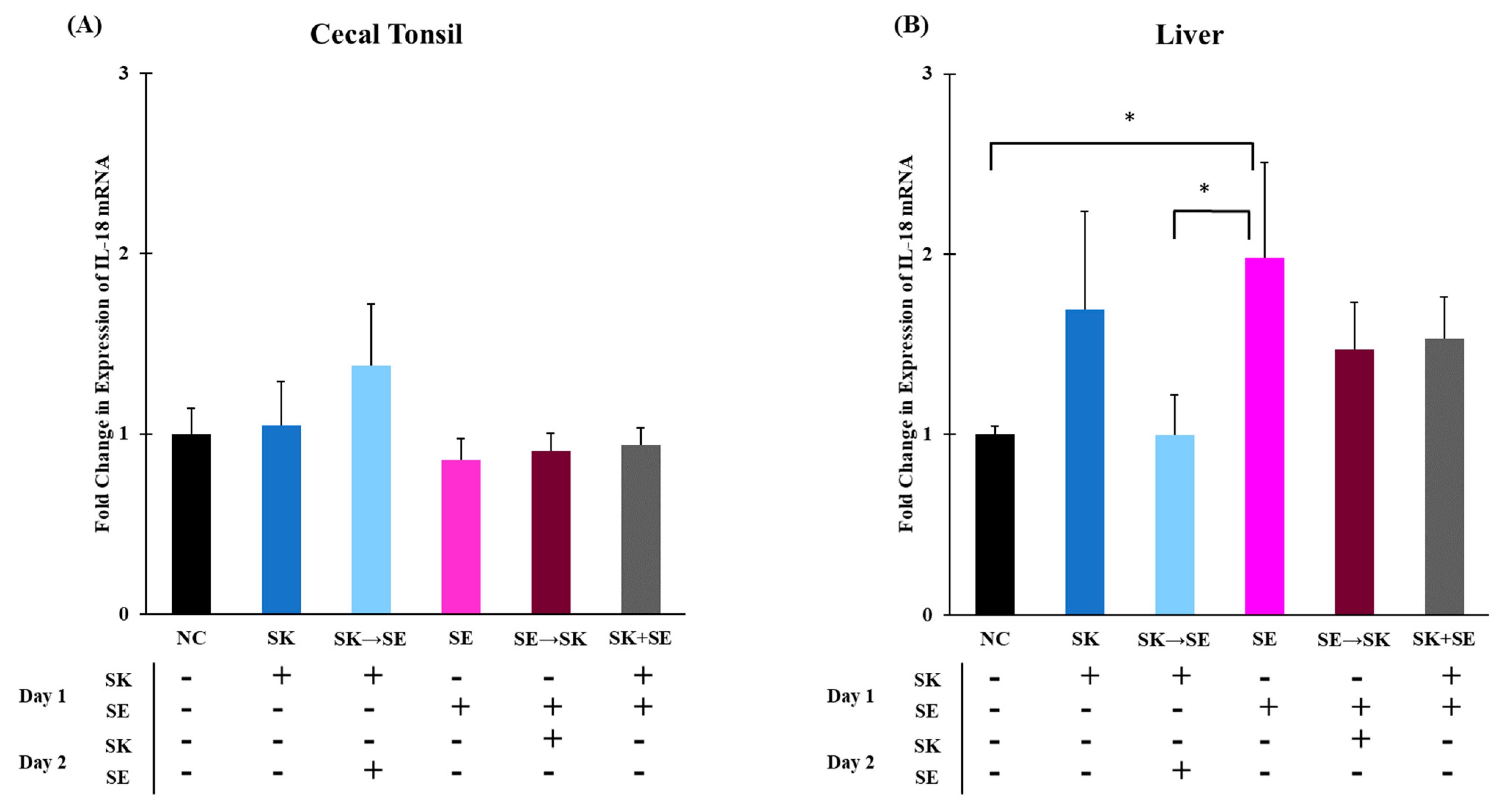
Figure 7B). The mRNA levels of IL-18 were significantly higher in the SK followed by SE treatment than the SK only, SE only, SE followed by SK, and SK and SE combination treatments in the cecal tonsils (
Figure 8A). There were no changes in the liver (
Figure 8B). Berndt and colleagues measured a significant increase in IL-18 mRNA levels in the ceca of chicks gavaged with 1.0 × 10
7 CFU of SE or ST at a peak 2 and 4 days post-infection [
74]. Secretion of IL-18 is important for a later adaptive immune response to produce IFN-γ [
30].
Chicken IFN-γ is primarily produced by T
H1 lymphocytes and natural killer cells and is driven by the production of IL-12 and IL-18 for a later immune response [
74,
75]. Expression of IFN-γ is critical to the host immune response to intracellular pathogens because it activates macrophages, which increases their ability to kill [
76]. There were no significant changes in IFN-γ expression in the cecal tonsils and liver in trial 1 (
Figure 9A,B). The mRNA levels of IFN-γ were not different in the cecal tonsils (
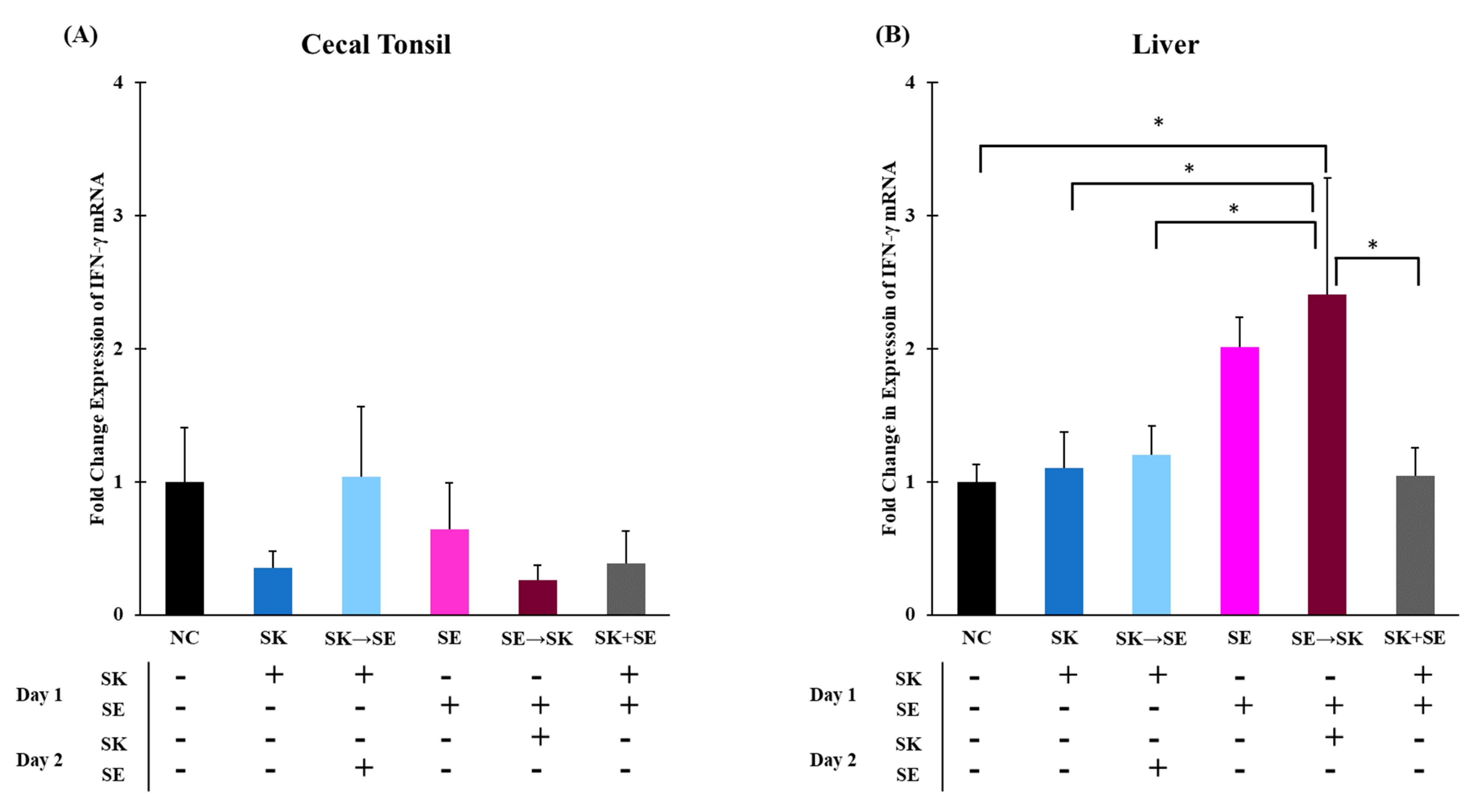
Figure 10A). Levels of IFN-γ were significantly higher in the SE → SK group compared to the NC, SK, SK → SE, SE, and SK+SE groups in the liver (
Figure 10B). Gamma interferon expression is increased by signals from pro-inflammatory or T
H1 cytokines such as IL-6 and IL-18 in an adaptive immune response [
74,
77]. Berndt and colleagues measured a significant increase IFN-γ mRNA levels in the ceca up to 4 days post-infection [
76].
Cytokine mRNA expression measured by (qRT)-PCR does not always correlate with protein levels, but it is a sensitive method [
78]. An increase in cytokine expression has been found as early as 12 h post-infection [
35]. Here we measured cytokine mRNA expression 24–48 h post-infection and the responses could have had an earlier or later expression pattern. Broiler chick ceca are colonized by
Salmonella quicker than the spleen and liver which could explain the variable results of our mRNA levels [
55]. The cecal tonsils are a part of the gut-associated lymphoid tissues and are a local site for immune responses against enteric bacteria [
79]. The liver produces acute phase proteins in response to pro-inflammatory cytokines [
80]. There was a lower incidence of
Salmonella in the liver/spleen macerations, therefore, it may be that not all the livers were affected like the ceca. However, soluble factors from the gastrointestinal tract can affect the liver, so that could explain the results seen [
37]. Cheeseman and colleagues measured an increase of IFN-γ in the spleen compared to the ceca in birds challenged with SE, indicating differences in immune gene expression across organs [
81]. Withanage and colleagues found initial cytokine detection was greater in the liver before the spleen indicating a more rapid response [
38]. The differences in cytokine gene expression suggest differences in
Salmonella subspecies’ interactions and pathogenesis [
82].
The current research focused on the characterization of an immune response in the cecal tonsils and liver during a concurrent infection. Intestinal cytokine responses are stimulated by foreign or pathogenic bacteria [
83].
Salmonella may not be pathogenic to the avian host, but it can persist and colonize the cecal lumen of chickens, which would allow shedding into the environment [
84]. As previously mentioned,
Salmonella Kentucky was isolated from fewer human cases, than ST and SE [
8]. Yet, SK was isolated from more chicken carcasses than ST and SE combined [
3]. In conclusion, the data presented show that the oral administration of
Salmonella Kentucky reduced subsequent colonization of Enteritidis and Typhimurium in neonatal broilers. The current study shows expression of cytokines were affected by consecutive challenges indicating immune function could be altered during competitive exclusion. A subunit vaccine exploiting SK’s mechanisms to colonize and persist in chickens could benefit public health and agricultural sectors.


 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}