
Dysbiosis in Pediatrics Is Associated with Respiratory Infections: Is There a Place for Bacterial-Derived Products?



Abstract
:1. Introduction
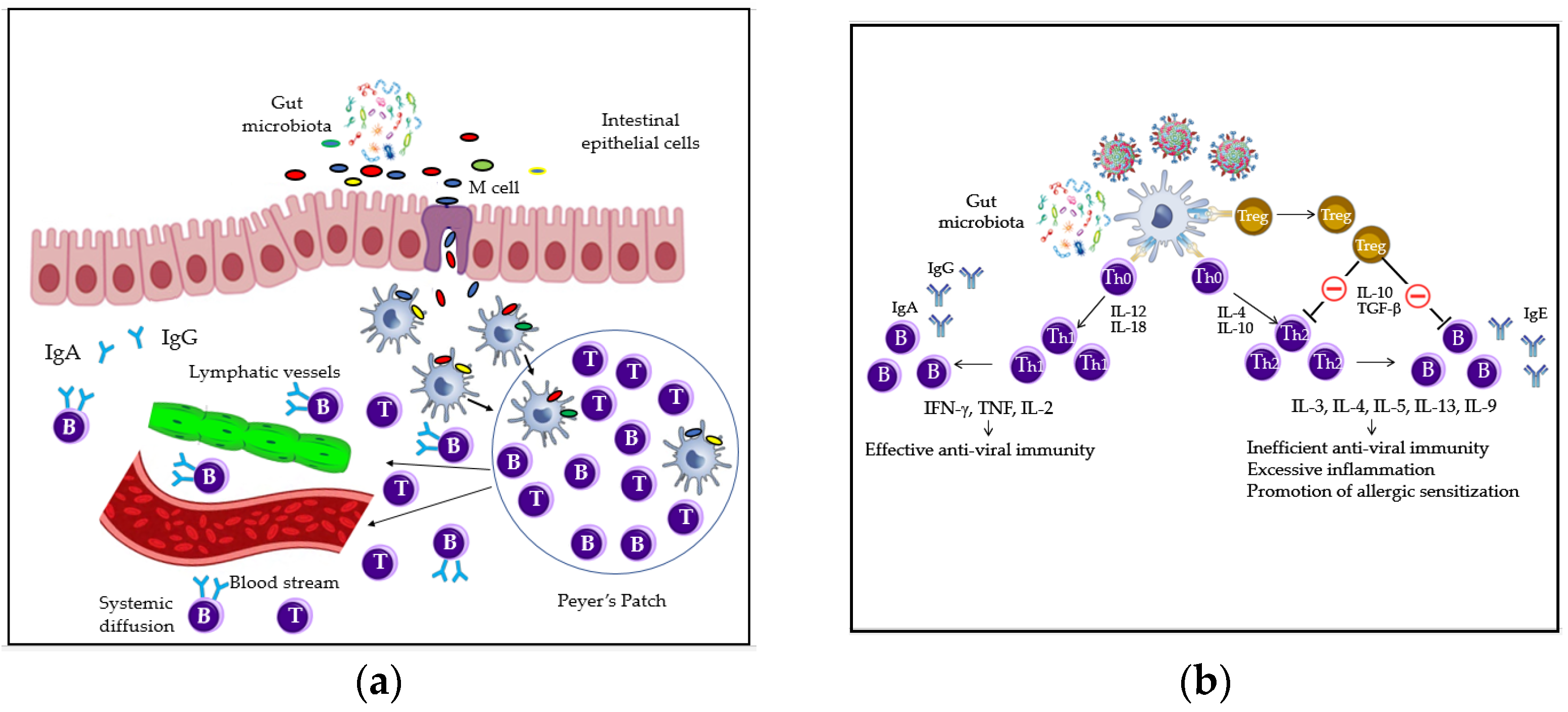
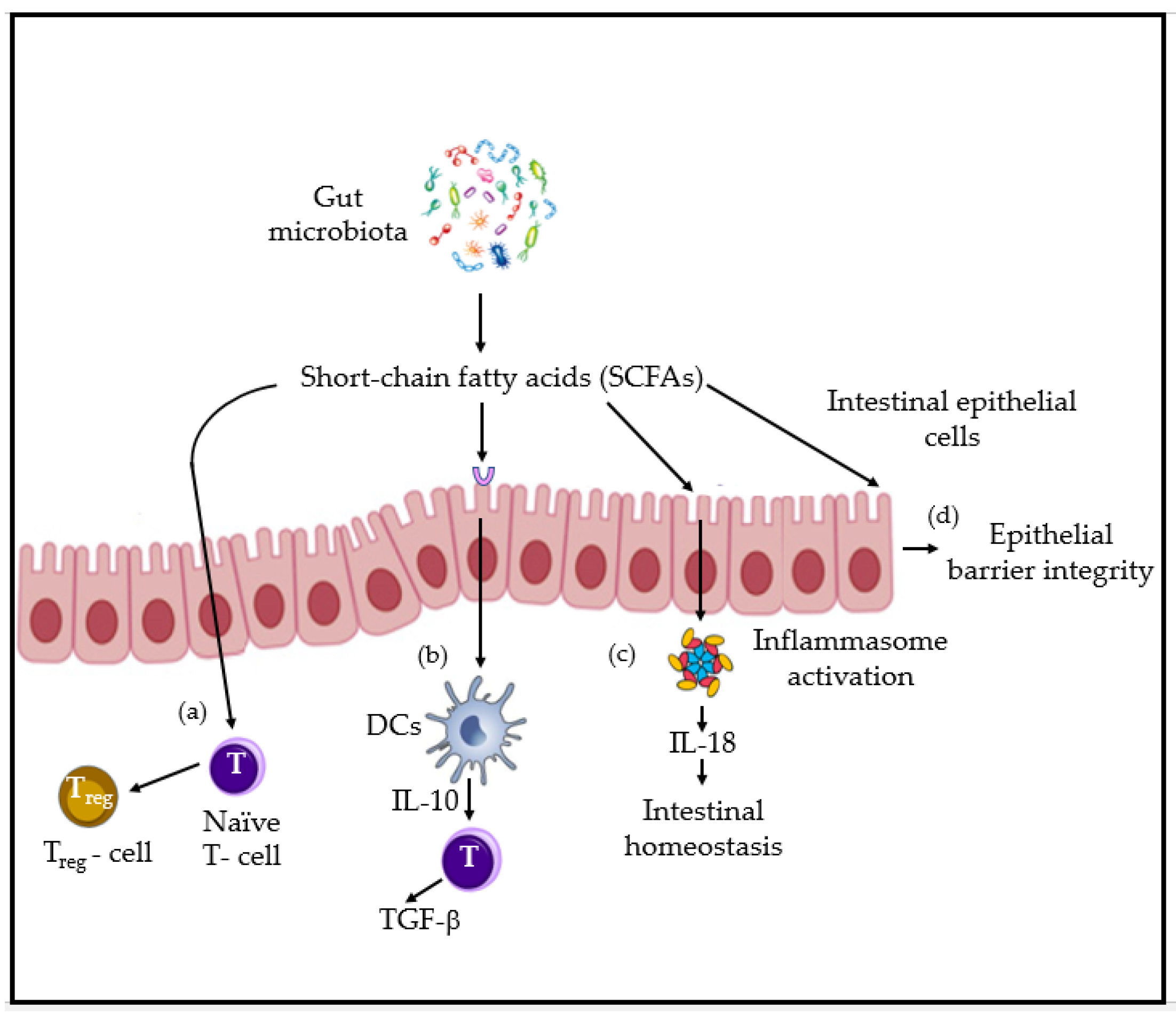
2. Gut and Airway Microbiota and Immune Homeostasis: Key Players, Interactions and Mechanisms Overview
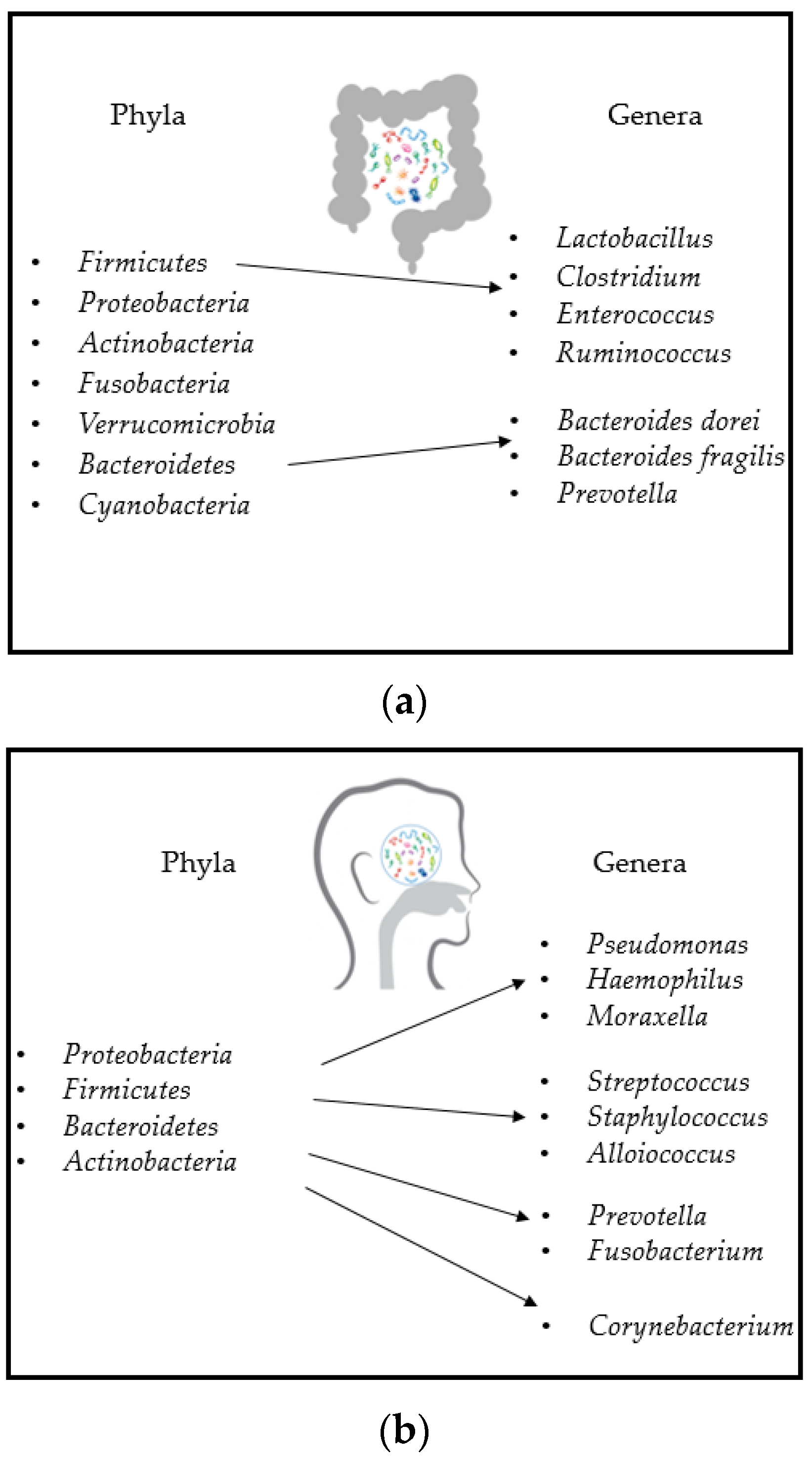
3. Gut and Airway Microbiota Composition

4. Gut and Airways Dysbiosis in Respiratory Diseases
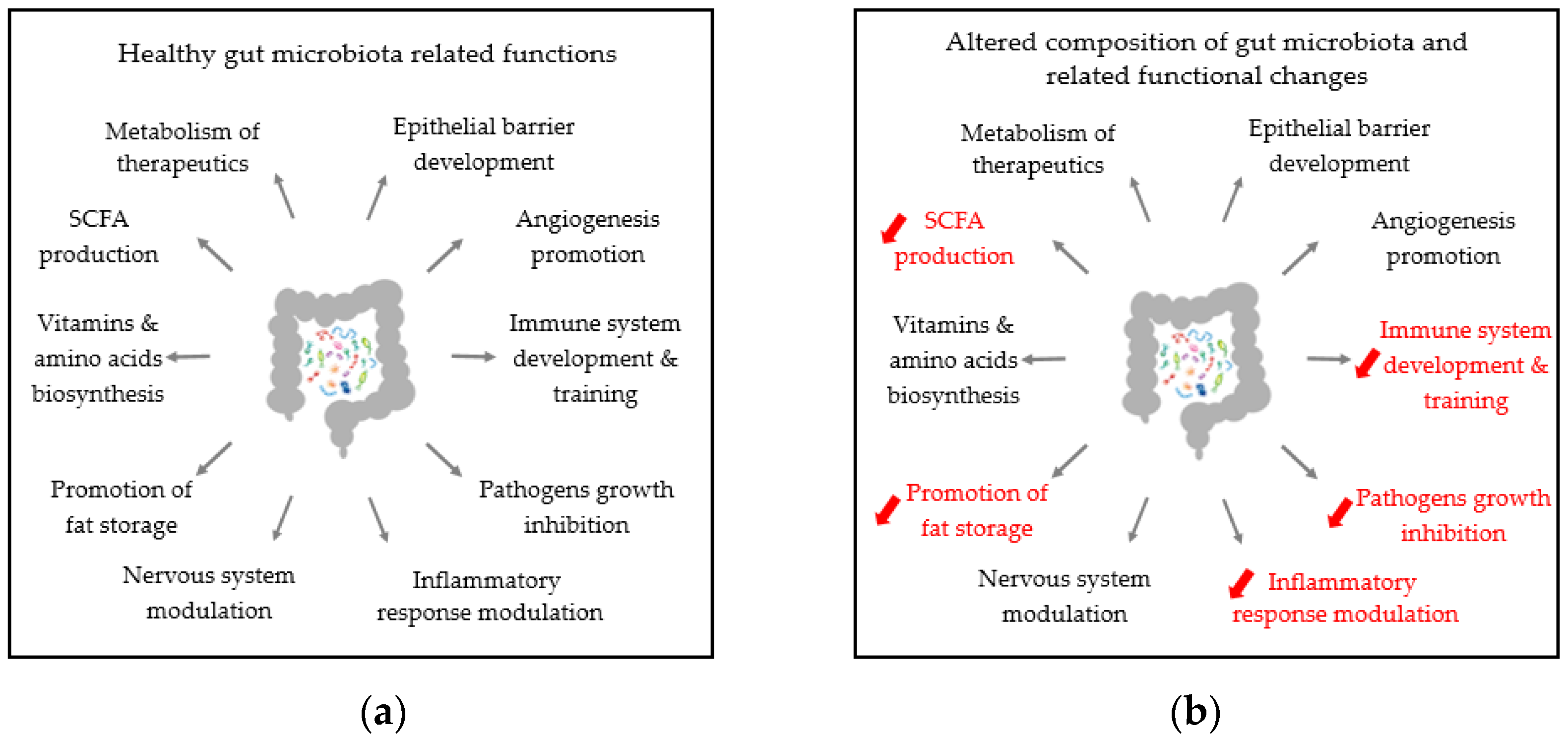
4.1. General Features of Gut and Airways Dysbiosis
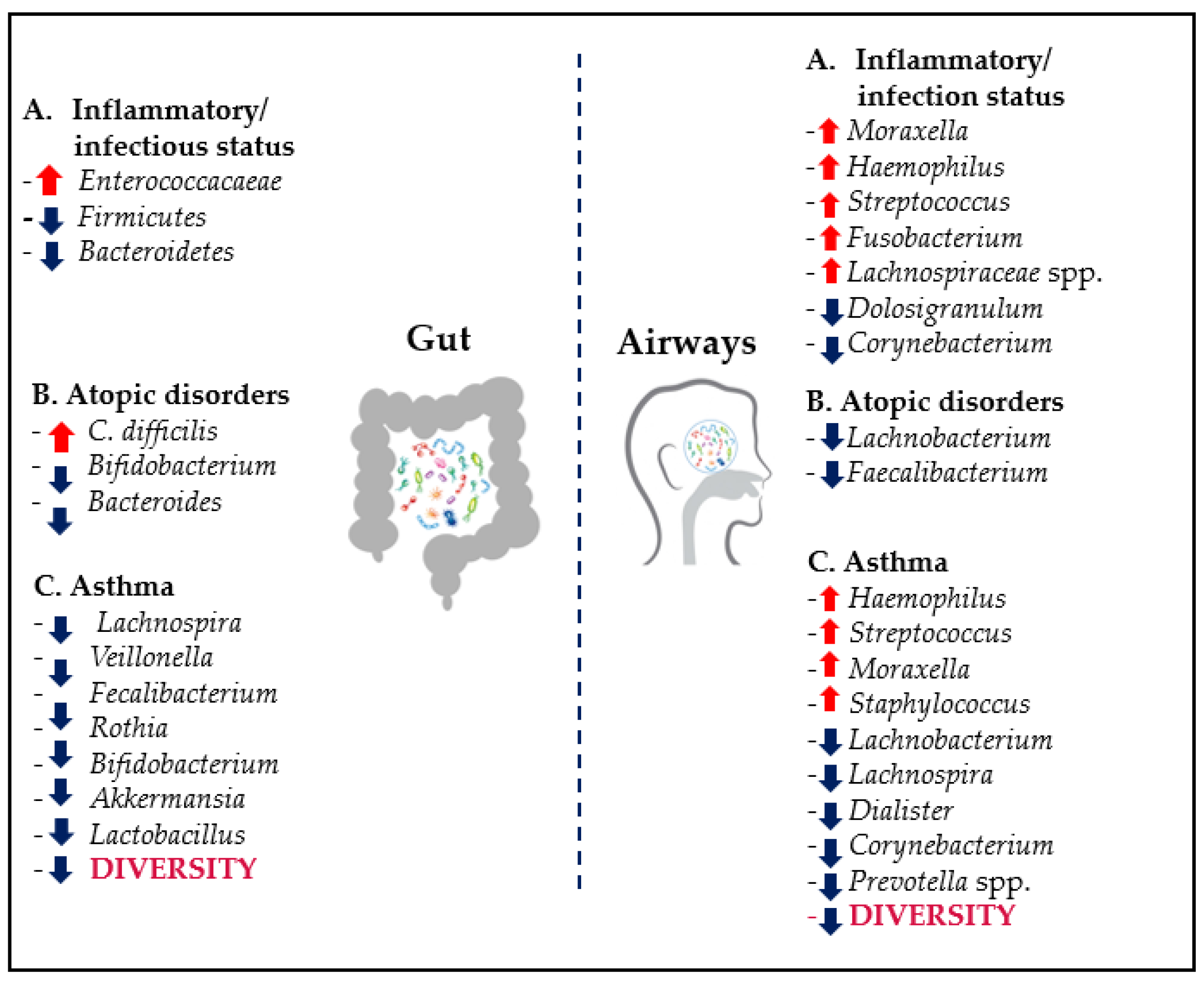
4.2. Dysbiosis and Respiratory Diseases
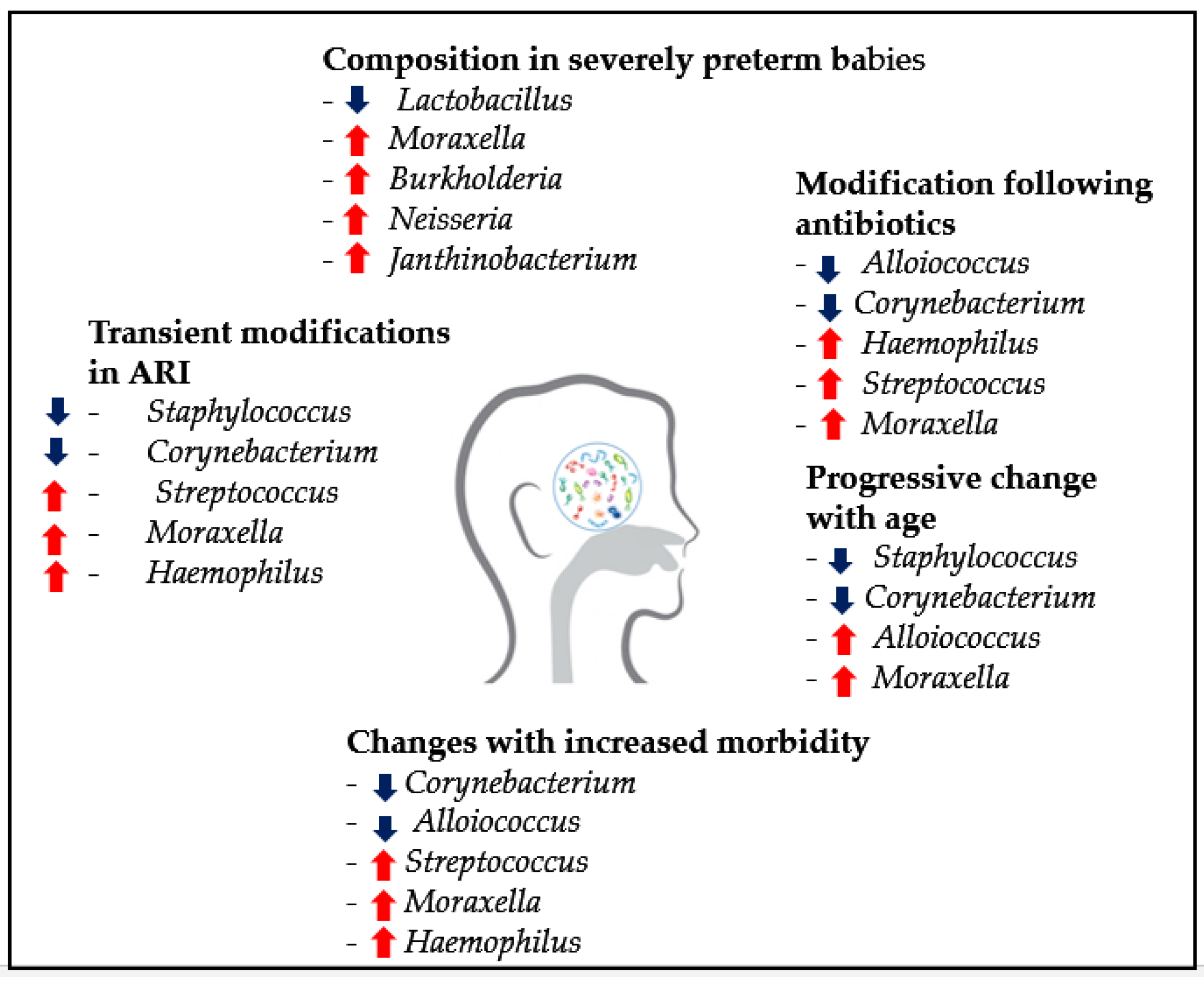
4.2.1. Pediatric RTIs and Recurrences
4.2.2. Pediatric Asthma
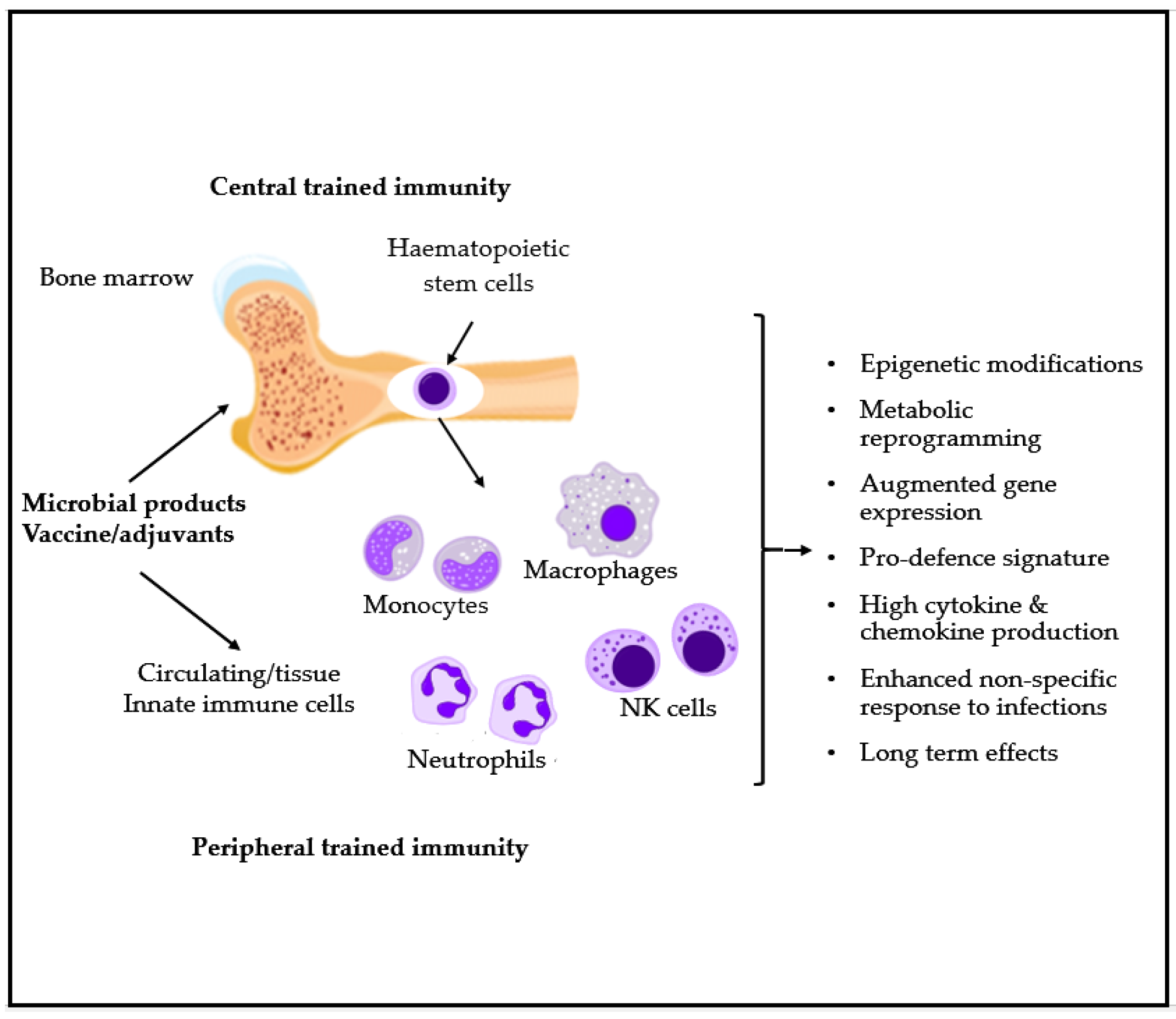
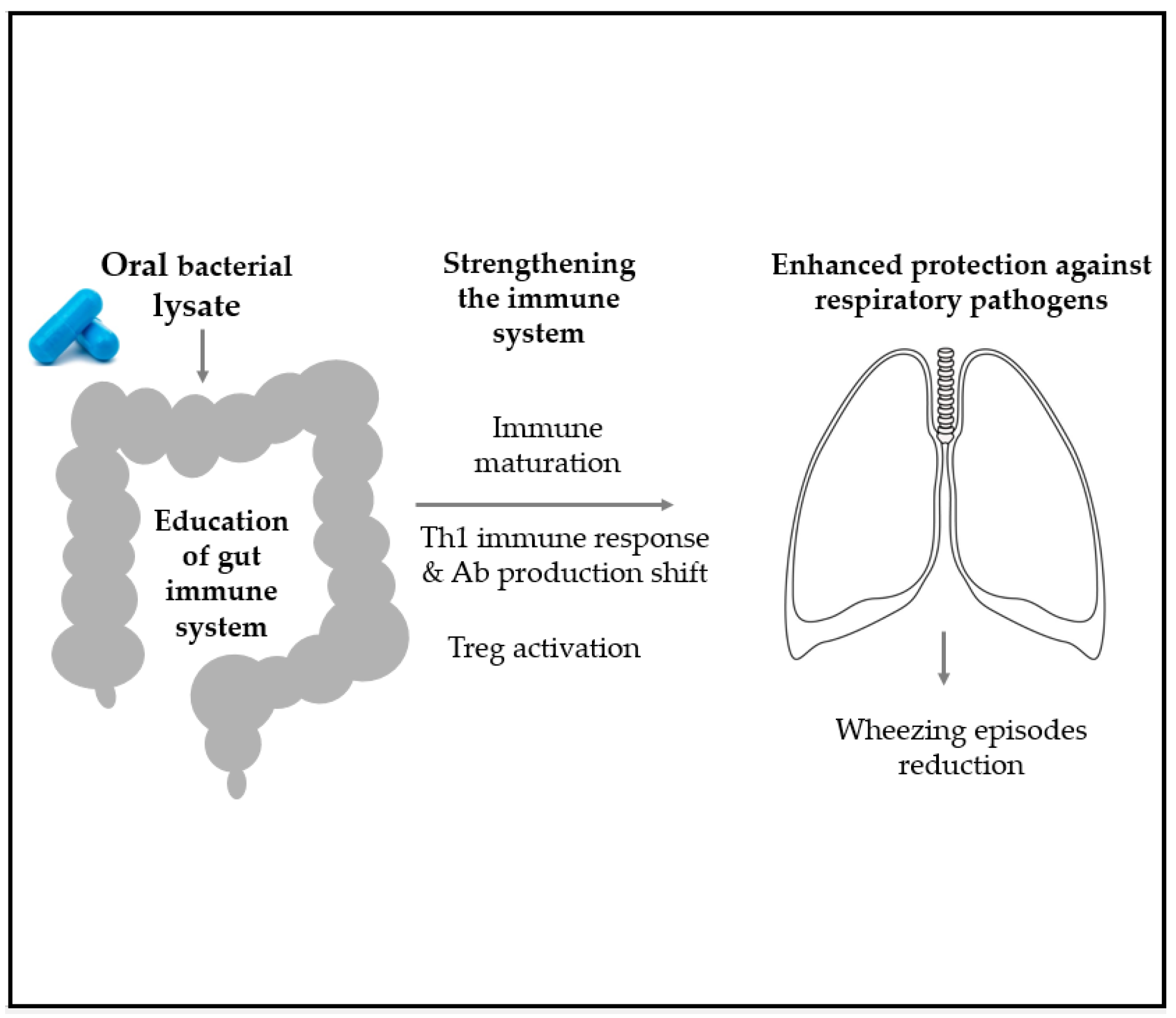
5. Microbial-Derived Products as Possible Interventions in Pediatric Respiratory Recurrence and Asthma
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kusel, M.M.H.; De Klerk, N.; Holt, P.G.; I Landau, L.; Sly, P.D. Occurrence and management of acute respiratory illnesses in early childhood. J. Paediatr. Child Health 2007, 43, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Jain, N.; Lodha, R.; Kabra, S.K. Upper respiratory tract infections. Indian J. Pediatr. 2001, 68, 1135–1138. [Google Scholar] [CrossRef] [Green Version]
- Morris, P.S. Upper Respiratory Tract Infections (Including Otitis Media). Pediatr. Clin. N. Am. 2009, 56, 101–117. [Google Scholar] [CrossRef]
- WHO. WHO Report Prepared for the WHO Annual Consultation on the Composition of Influenza Vaccine for the Northern Hemisphere 2017-18; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Esposito, S.; Daleno, C.; Tagliabue, C.; Scala, A.; Tenconi, R.; Borzani, I.; Fossali, E.; Pelucchi, C.; Piralla, A.; Principi, N. Impact of rhinoviruses on pediatric community-acquired pneumonia. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 31, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- Hament, J.M.; Kimpen, J.L.; Fleer, A.; Wolfs, T.F. Respiratory viral infection predisposing for bacterial disease: A concise review. FEMS Immunol. Med. Microbiol. 1999, 26, 189–195. [Google Scholar] [CrossRef]
- Schaad, U.; Principi, N. The management of recurrent respiratory tract infections in children. Eur. Infect. Dis. 2012, 6, 111–115. [Google Scholar]
- Dethlefsen, L.; Huse, S.; Sogin, M.L.; A Relman, D. The Pervasive Effects of an Antibiotic on the Human Gut Microbiota, as Revealed by Deep 16S rRNA Sequencing. PLoS Biol. 2008, 6, e280. [Google Scholar] [CrossRef] [PubMed]
- Sly, P.D.; Holt, P.G. Role of innate immunity in the development of allergy and asthma. Curr. Opin. Allergy Clin. Immunol. 2011, 11, 127–131. [Google Scholar] [CrossRef]
- Jesenak, M.; Ciljakova, M.; Rennerova, Z.; Babusikova, E.; Banovci, P. Recurrent Respiratory Infections in Children—Definition, Diagnostic Approach, Treatment and Prevention. Bronchitis 2011. [Google Scholar] [CrossRef] [Green Version]
- Bellanti, J.A. Recurrent Respiratory Tract Infections in Paediatric Patients. Drugs 1997, 54, 1–4. [Google Scholar] [CrossRef]
- Zomer-Kooijker, K.; van der Ent, C.K.; Ermers, M.J.; Uiterwaal, C.S.; Rovers, M.M.; Bont, L.J.; RSV Corticosteroid Study Group. Increased risk of wheeze and de-creased lung function after respiratory syncytial virus infection. PLoS ONE 2014, 9, e87162. [Google Scholar]
- Ratajczak, W.; Ryl, A.; Mizerski, A.; Walczakiewicz, K.; Sipak, O.; Laszczynska, M. Immunomodulatory potential of gut microbi-ome-derived short-chain fatty acids (SCFAs). Acta Biochim. Pol. 2019, 66, 1–12. [Google Scholar]
- Taylor, S.L.; Wesselingh, S.; Rogers, G.B. Host-microbiome interactions in acute and chronic respiratory infections. Cell. Microbiol. 2016, 18, 652–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, J.-U.; Lee, S.E.; Hwang, W.; Lee, C.; Park, J.-W.; Sohn, J.-H.; Nam, J.H.; Kim, Y.; Rhee, J.H.; Im, S.-H.; et al. Flagellin suppresses experimental asthma by generating regulatory dendritic cells and T cells. J. Allergy Clin. Immunol. 2016, 137, 426–435. [Google Scholar] [CrossRef]
- Ober, C.; Sperling, A.I.; Von Mutius, E.; Vercelli, D. Immune development and environment: Lessons from Amish and Hutterite children. Curr. Opin. Immunol. 2017, 48, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.; Mitchell, A.L.; Boland, M.; Forster, S.C.; Gloor, G.B.; Tarkowska, A.; Lawley, T.D.; Finn, R.D. A new genomic blueprint of the human gut microbiota. Nat. Cell Biol. 2019, 568, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Esposito, S.; Soto-Martinez, M.E.; Feleszko, W.; Jones, M.H.; Shen, K.-L.; Schaad, U.B. Nonspecific immunomodulators for recurrent respiratory tract infections, wheezing and asthma in children: A systematic review of mechanistic and clinical evidence. Curr. Opin. Allergy Clin. Immunol. 2018, 18, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, F.; Lombardi, E.; Rossi, O.; Bagnasco, D.; Bellocchi, A.; Menzella, F. Epithelial dysfunction, respiratory infections and asthma: The importance of immunomodulation. A focus on OM-85. Expert Rev. Respir. Med. 2020, 14, 1019–1026. [Google Scholar] [CrossRef]
- Koppen, I.J.N.; Bosch, A.A.T.M.; Sanders, E.A.M.; van Houten, M.A.; Bogaert, D. The respiratory microbiota during health and disease: A paediatric perspective. Pneumonia 2015, 6, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Bisgaard, H.; Li, N.; Bonnelykke, K.; Chawes, B.L.K.; Skov, T.; Paludan-Müller, G.; Stokholm, J.; Smith, B.; Krogfelt, K.A. Reduced diversity of the intestinal microbiota during infancy is associated with increased risk of allergic disease at school age. J. Allergy Clin. Immunol. 2011, 128, 646–652.e5. [Google Scholar] [CrossRef]
- Pandiyan, P.; Bhaskaran, N.; Zou, M.; Schneider, E.; Jayaraman, S.; Huehn, J. Microbiome Dependent Regulation of T(regs) and Th17 Cells in Mucosa. Front. Immunol. 2019, 10, 426. [Google Scholar] [CrossRef] [Green Version]
- Rossi, G.A.; Pohunek, P.; Feleszko, W.; Ballarini, S.; Colin, A.A. Viral infections and wheezing–asthma inception in childhood: Is there a role for immunomodulation by oral bacterial lysates? Clin. Transl. Allergy 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Artis, D. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat. Rev. Immunol. 2008, 8, 411–420. [Google Scholar] [CrossRef]
- Shapiro, H.; A Thaiss, C.; Levy, M.; Elinav, E. The cross talk between microbiota and the immune system: Metabolites take center stage. Curr. Opin. Immunol. 2014, 30, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Tramontano, M.; Andrejev, S.; Pruteanu, M.; Klünemann, M.; Kuhn, M.; Galardini, M.; Jouhten, P.; Zelezniak, A.; Zeller, G.; Bork, P.; et al. Nutritional preferences of human gut bacteria reveal their metabolic idiosyncrasies. Nat. Microbiol. 2018, 3, 514–522. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teo, S.M.; Mok, D.; Pham, K.; Kusel, M.; Serralha, M.; Troy, N.; Holt, B.J.; Hales, B.J.; Walker, M.L.; Hollams, E.; et al. The Infant Nasopharyngeal Microbiome Impacts Severity of Lower Respiratory Infection and Risk of Asthma Development. Cell Host Microbe 2015, 17, 704–715. [Google Scholar] [CrossRef] [Green Version]
- Gollwitzer, E.S.; Saglani, S.; Trompette, A.; Yadava, K.; Sherburn, R.; McCoy, K.D.; Nicod, L.P.; Lloyd, C.M.; Marsland, B.J. Lung microbiota promotes tolerance to allergens in neonates via PD-L1. Nat. Med. 2014, 20, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The Impact of the Gut Microbiota on Human Health: An Integrative View. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cait, A.; Hughes, M.R.; Antignano, F.; Cait, J.; Dimitriu, P.A.; Maas, K.R.; Reynolds, L.A.; Hacker, L.; Mohr, J.; Finlay, B.B.; et al. Microbiome-driven allergic lung inflammation is ameliorated by short-chain fatty acids. Mucosal Immunol. 2018, 11, 785–795. [Google Scholar] [CrossRef] [PubMed]
- de Laval, B.; Maurizio, J.; Kandalla, P.K.; Brisou, G.; Simonnet, L.; Huber, C.; Gimenez, G.; Matcovitch-Natan, O.; Reinhardt, S.; David, E.; et al. C/EBPβ-Dependent Epigenetic Memory Induces Trained Immunity in Hematopoietic Stem Cells. Cell Stem Cell 2020, 26, 657–674.e8. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Giamarellos-Bourboulis, E.J.; Domínguez-Andrés, J.; Curtis, N.; van Crevel, R.; van de Veerdonk, F.L.; Bonten, M. Trained Immunity: A Tool for Reducing Susceptibility to and the Severity of SARS-CoV-2 Infection. Cell 2020, 181, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Clarke, T.B.; Davis, K.M.; Lysenko, E.S.; Zhou, A.Y.; Yu, Y.; Weiser, J.N. Recognition of peptidoglycan from the microbiota by Nod1 enhances systemic innate immunity. Nat. Med. 2010, 16, 228–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnenburg, J.L.; Sonnenburg, E.D. Vulnerability of the industrialized microbiota. Science 2019, 366, eaaw9255. [Google Scholar] [CrossRef] [Green Version]
- Roquilly, A.; Torres, A.; Villadangos, J.A.; Netea, M.G.; Dickson, R.; Becher, B.; Asehnoune, K. Pathophysiological role of respiratory dysbiosis in hospital-acquired pneumonia. Lancet Respir. Med. 2019, 7, 710–720. [Google Scholar] [CrossRef]
- Gosens, R.; Hiemstra, P.S.; Adcock, I.M.; Bracke, K.R.; Dickson, R.P.; Hansbro, P.M.; Krauss-Etschmann, S.; Smits, H.H.; Stassen, F.R.M.; Bartel, S. Host-microbe cross-talk in the lung microenvironment: Implications for understanding and treating chronic lung disease. Eur. Respir. J. 2020, 56, 1902320. [Google Scholar] [CrossRef] [PubMed]
- Dickson, R.P.; Erb-Downward, J.R.; Martinez, F.J.; Huffnagle, G.B. The Microbiome and the Respiratory Tract. Annu. Rev. Physiol. 2016, 78, 481–504. [Google Scholar] [CrossRef] [Green Version]
- Sakwinska, O.; Schmid, V.B.; Berger, B.; Bruttin, A.; Keitel, K.; Lepage, M.; Moine, D.; Bru, C.N.; Brüssow, H.; Gervaix, A. Nasopharyngeal Microbiota in Healthy Children and Pneumonia Patients. J. Clin. Microbiol. 2014, 52, 1590–1594. [Google Scholar] [CrossRef] [Green Version]
- Murphy, T.F.; Bakaletz, L.O.; Smeesters, P.R. Microbial Interactions in the Respiratory Tract. Pediatr. Infect. Dis. J. 2009, 28, S121–S126. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Marsland, B.J. Influences of the Microbiome on the Early Origins of Allergic Asthma. Ann. Am. Thorac. Soc. 2013, 10, S165–S169. [Google Scholar] [CrossRef]
- Hilty, M.; Burke, C.; Pedro, H.; Cardenas, P.; Bush, A.; Bossley, C.; Davies, J.; Ervine, A.; Poulter, L.; Pachter, L.; et al. Disordered microbial communities in asthmatic airways. PLoS ONE 2010, 5, e8578. [Google Scholar] [CrossRef] [Green Version]
- Pichon, M.; Lina, B.; Josset, L. Impact of the Respiratory Microbiome on Host Responses to Respiratory Viral Infection. Vaccines 2017, 5, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claassen, S.; Du Toit, E.; Kaba, M.; Moodley, C.; Zar, H.J.; Nicol, M.P. A comparison of the efficiency of five different commercial DNA extraction kits for extraction of DNA from faecal samples. J. Microbiol. Methods 2013, 94, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut Microbiota in Health and Disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.J.; Lynch, D.B.; Murphy, K.; Ulaszewska, M.; Jeffery, I.B.; O’Shea, C.A.; Watkins, C.; Dempsey, E.; Mattivi, F.; Tuohy, K.; et al. Evolution of gut microbiota composition from birth to 24 weeks in the INFANTMET Cohort. Microbiome 2017, 5, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, F.; Liu, Y.; Gu, F. Intestinal microbiota dysbiosis in children with recurrent respiratory tract infections. Microb. Pathog. 2019, 136, 103709. [Google Scholar] [CrossRef]
- Bosch, A.A.T.M.; Piters, W.A.A.D.S.; Van Houten, M.A.; Chu, M.L.J.N.; Biesbroek, G.; Kool, J.; Pernet, P.; De Groot, P.-K.C.M.; Eijkemans, M.J.C.; Keijser, B.J.F.; et al. Maturation of the Infant Respiratory Microbiota, Environmental Drivers, and Health Consequences. A Prospective Cohort Study. Am. J. Respir. Crit. Care Med. 2017, 196, 1582–1590. [Google Scholar] [CrossRef]
- Robinson, P.F.M.; Pattaroni, C.; Cook, J.; Gregory, L.; Alonso, A.M.; Fleming, L.J.; Lloyd, C.M.; Bush, A.; Marsland, B.J.; Saglani, S. Lower airway microbiota associates with inflammatory phenotype in severe preschool wheeze. J. Allergy Clin. Immunol. 2019, 143, 1607–1610.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, R.K.K.; Zhou, J.W.; Guan, W.; Li, S.K.; Yang, Z.F.; Tsui, S.K. Modulation of potential respiratory pathogens by pH1N1 viral in-fection. Clin. Microbiol. Infect. 2013, 19, 930–935. [Google Scholar] [CrossRef] [Green Version]
- Tregoning, J.S.; Schwarze, J. Respiratory Viral Infections in Infants: Causes, Clinical Symptoms, Virology, and Immunology. Clin. Microbiol. Rev. 2010, 23, 74–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, G.A.; Fanous, H.; Colin, A.A. Viral strategies predisposing to respiratory bacterial superinfections. Pediatr. Pulmonol. 2020, 55, 1061–1073. [Google Scholar] [CrossRef]
- Rosas-Salazar, C.; Shilts, M.H.; Tovchigrechko, A.; Schobel, S.; Chappell, J.D.; Larkin, E.K.; Shankar, J.; Yooseph, S.; Nelson, K.E.; Halpin, R.A.; et al. Differences in the Nasopharyngeal Microbiome During Acute Respiratory Tract Infection with Human Rhinovirus and Respiratory Syncytial Virus in Infancy. J. Infect. Dis. 2016, 214, 1924–1928. [Google Scholar] [CrossRef] [PubMed]
- de Steenhuijsen Piters, W.A.; Heinonen, S.; Hasrat, R.; Bunsow, E.; Smith, B.; Suarez-Arrabal, M.C.; Chaussabel, D.; Cohen, D.M.; Sanders, E.A.; Ramilo, O.; et al. Nasopharyngeal Microbiota, Host Transcriptome, and Disease Severity in Children with Respiratory Syncytial Virus Infection. Am. J. Respir. Crit. Care Med. 2016, 194, 1104–1115. [Google Scholar] [CrossRef]
- Ichinohe, T.; Pang, I.K.; Kumamoto, Y.; Peaper, D.R.; Ho, J.H.; Murray, T.S.; Hiwasaki, A. Microbiota regulates immune defense against respiratory tract influenza A virus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 5354–5359. [Google Scholar] [CrossRef] [Green Version]
- Martinez, F.D.; Guerra, S. Early Origins of Asthma. Role of Microbial Dysbiosis and Metabolic Dysfunction. Am. J. Respir. Crit. Care Med. 2018, 197, 573–579. [Google Scholar] [CrossRef]
- Sigurs, N.; AlJassim, F.; Kjellman, B.; Robinson, P.D.; Sigurbergsson, F.; Bjarnason, R.; Gustafsson, P.M. Asthma and allergy patterns over 18 years after severe RSV bronchiolitis in the first year of life. Thorax 2010, 65, 1045–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, M.R.; Bartlett, N.W.; Hussell, T.; Openshaw, P.; Johnston, S.L. The microbiology of asthma. Nat. Rev. Genet. 2012, 10, 459–471. [Google Scholar] [CrossRef]
- Penders, J.; Thijs, C.; Brandt, P.A.V.D.; Kummeling, I.; Snijders, B.; Stelma, F.; Adams, H.; Van Ree, R.; Stobberingh, E.E. Gut microbiota composition and development of atopic manifestations in infancy: The KOALA Birth Cohort Study. Gut 2007, 56, 661–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Nimwegen, F.A.; Penders, J.; Stobberingh, E.E.; Postma, D.S.; Koppelman, G.H.; Kerkhof, M.; Reijmerink, N.E.; Dompeling, E.; van den Brandt, P.A.; Ferreira, I.; et al. Mode and place of delivery, gastrointestinal microbiota, and their influence on asthma and atopy. J. Allergy Clin. Immunol. 2011, 128, e1–e3. [Google Scholar] [CrossRef] [PubMed]
- Galazzo, G.; Van Best, N.; Bervoets, L.; Dapaah, I.O.; Savelkoul, P.H.; Hornef, M.W.; Lau, S.; Hamelmann, E.; Penders, J.; Hutton, E.K.; et al. Development of the Microbiota and Associations with Birth Mode, Diet, and Atopic Disorders in a Longitudinal Analysis of Stool Samples, Collected from Infancy through Early Childhood. Gastroenterology 2020, 158, 1584–1596. [Google Scholar] [CrossRef]
- Arrieta, M.-C.; Stiemsma, L.T.; Dimitriu, P.A.; Thorson, L.; Russell, S.; Yurist-Doutsch, S.; Kuzeljevic, B.; Gold, M.J.; Britton, H.M.; Lefebvre, D.L.; et al. Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci. Transl. Med. 2015, 7, 307ra152. [Google Scholar] [CrossRef] [PubMed]
- van den Elsen, L.W.; Poyntz, H.C.; Weyrich, L.S.; Young, W.; Forbes-Blom, E.E. Embracing the gut microbiota: The new frontier for inflammatory and infectious diseases. Clin. Transl. Immunol. 2017, 6, e125. [Google Scholar] [CrossRef]
- Fujimura, K.E.; Sitarik, A.R.; Havstad, S.; Lin, D.L.; LeVan, S.; Fadrosh, D.; Panzer, A.R.; LaMere, B.; Rackaityte, E.; Lukacs, N.W.; et al. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat. Med. 2016, 22, 1187–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vael, C.; Nelen, V.; Verhulst, S.L.; Goossens, H.; Desager, K.N. Early intestinal Bacteroides fragilis colonisation and development of asthma. BMC Pulm. Med. 2008, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Lazar, V.; Ditu, L.-M.; Pircalabioru, G.G.; Gheorghe, I.; Curutiu, C.; Holban, A.M.; Picu, A.; Petcu, L.; Chifiriuc, M.C. Aspects of Gut Microbiota and Immune System Interactions in Infectious Diseases, Immunopathology, and Cancer. Front. Immunol. 2018, 9, 1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, P.G. Key Factors in the Development of Asthma: Atopy. Am. J. Respir. Crit. Care Med. 2000, 161, S172–S175. [Google Scholar] [CrossRef]
- Smits, H.H.; Hiemstra, P.S.; Prazeres da Costa, C.; Ege, M.; Edwards, M.; Garn, H.; Howarth, P.H.; Jartti, T.; de Jong, E.C.; Maizels, R.M.; et al. Microbes and asthma: Opportunities for intervention. J. Allergy Clin. Immunol. 2016, 137, 690–697. [Google Scholar] [CrossRef] [Green Version]
- Hufnagl, K.; Pali-Schöll, I.; Roth-Walter, F.; Jensen-Jarolim, E. Dysbiosis of the gut and lung microbiome has a role in asthma. Semin. Immunopathol. 2020, 42, 75–93. [Google Scholar] [CrossRef] [Green Version]
- van den Bergh, M.R.; Biesbroek, G.; Rossen, J.W.; de Steenhuijsen Piters, W.A.; Bosch, A.A.; van Gils, E.J.; Wang, X.; Boonacker, C.W.; Veenhoven, R.H.; Bruin, J.P.; et al. Associations between pathogens in the upper respiratory tract of young children: Interplay between viruses and bacteria. PLoS ONE 2012, 7, e47711. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, F.; Sun, R.; Gao, X.; Wei, H.; Li, L.J.; Tian, Z. Bacterial colonization dampens influenza-mediated acute lung injury via induction of M2 alveolar macrophages. Nat. Commun. 2013, 4, 2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsythe, P. Probiotics and Lung Diseases. Chest 2011, 139, 901–908. [Google Scholar] [CrossRef]
- Giovannini, M.; Agostoni, C.; Riva, E.; Salvini, F.; Ruscitto, A.; Zuccotti, G.V.; Radaelli, G. A Randomized Prospective Double Blind Controlled Trial on Effects of Long-Term Consumption of Fermented Milk Containing Lactobacillus casei in Pre-School Children with Allergic Asthma and/or Rhinitis. Pediatr. Res. 2007, 62, 215–220. [Google Scholar] [CrossRef]
- Helin, T.; Haahtela, S. No effect of oral treatment with an intestinal bacterial strain, Lactobacillus rhamnosus (ATCC 53103), on birch-pollen allergy: A placebo-controlled double-blind study. Allergy 2002, 57, 243–246. [Google Scholar] [CrossRef]
- Ciprandi, G.; La Mantia, I.; Damiani, V.; Passali, D. Local Bacteriotherapy—A promising preventive tool in recurrent respiratory infections. Expert Rev. Clin. Immunol. 2020, 16, 1047–1052. [Google Scholar] [CrossRef] [PubMed]
- Meade, K.G.; O’Farrelly, C. β-Defensins: Farming the Microbiome for Homeostasis and Health. Front. Immunol. 2019, 9, 3072. [Google Scholar] [CrossRef] [PubMed]
- Strickland, D.H.; Judd, S.; Thomas, J.A.; Larcombe, A.N.; Sly, P.D.; Holt, P.G. Boosting airway T-regulatory cells by gastrointestinal stimulation as a strategy for asthma control. Mucosal Immunol. 2010, 4, 43–52. [Google Scholar] [CrossRef]
- Navarro, S.; Cossalter, G.; Chiavaroli, C.; Kanda, A.; Fleury, S.; Lazzari, A.; Cazareth, J.; Sparwasser, T.; Dombrowicz, D.; Glaichenhaus, N.; et al. The oral administration of bacterial extracts prevents asthma via the recruitment of regulatory T cells to the airways. Mucosal Immunol. 2010, 4, 53–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, A.; Gualdi, L.P.; de Souza, R.G.; Vargas, M.H.; Nuñez, N.K.; da Cunha, A.A.; Jones, M.H.; Pinto, L.A.; Stein, R.T.; Pitrez, P.M. Bacterial extract (OM-85) with human-equivalent doses does not inhibit the development of asthma in a murine model. Allergol. Immunopathol. 2016, 44, 504–511. [Google Scholar] [CrossRef]
- Holt, P.; Strickland, D. Low dose treatment of mice with bacterial extract (OM-85) for attenuation of experimental atopic asthma in mice. Allergol. Immunopathol. 2017, 45, 310–311. [Google Scholar] [CrossRef]
- Dang, A.T.; Pasquali, C.; Ludigs, K.; Guarda, G. OM-85 is an immunomodulator of interferon-β production and inflammasome activity. Sci. Rep. 2017, 7, 43844. [Google Scholar] [CrossRef] [Green Version]
- Esposito, S.; Bianchini, S.; Bosis, S.; Tagliabue, C.; Coro, I.; Argentiero, A.; Principi, N. A randomized, placebo-controlled, double-blinded, single-centre, phase IV trial to assess the efficacy and safety of OM-85 in children suffering from recurrent respiratory tract infections. J. Transl. Med. 2019, 17, 1–9. [Google Scholar] [CrossRef]
- Razi, C.H.; Harmancı, K.; Abacı, A.; Özdemir, O.; Hızlı, Ş.; Renda, R.; Keskin, F. The immunostimulant OM-85 BV prevents wheezing attacks in preschool children. J. Allergy Clin. Immunol. 2010, 126, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Karimi, K.; Inman, M.D.; Bienenstock, J.; Forsythe, P. Lactobacillus reuteri–induced Regulatory T cells Protect against an Allergic Airway Response in Mice. Am. J. Respir. Crit. Care Med. 2009, 179, 186–193. [Google Scholar] [CrossRef]
- A Rossi, G.; Esposito, S.; Feleszko, W.; Melioli, G.; Olivieri, D.; Piacentini, G.; Scaglione, F.; Vercelli, D. Immunomodulation Therapy—Clinical Relevance of Bacterial Lysates OM-85. Eur. Respir. Pulm. Dis. 2019, 5, 17–23. [Google Scholar] [CrossRef]
- Norton, E.B.; Clements, J.D.; Voss, T.G.; Cárdenas-Freytag, L. Prophylactic Administration of Bacterially Derived Immunomodulators Improves the Outcome of Influenza Virus Infection in a Murine Model. J. Virol. 2010, 84, 2983–2995. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; Domínguez-Andrés, J.; Barreiro, L.B.; Chavakis, T.; Divangahi, M.; Fuchs, E.; Joosten, L.A.B.; Van Der Meer, J.W.M.; Mhlanga, M.M.; Mulder, W.J.M.; et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 2020, 20, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Mahooti, M.; Miri, S.M.; Abdolalipour, E.; Ghaemi, A. The immunomodulatory effects of probiotics on respiratory viral infections: A hint for COVID-19 treatment? Microb. Pathog. 2020, 148, 104452. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Acronym | Indication | Patients/Children | Registry Number |
|---|---|---|---|
| OMPER | Prevention of RTIs | 288 | EudraCT2016-002705-19 |
| ORBEX | Prevention of LWRI | >800 1 | NCT02148796 |
| POWER | Prevention of severe LWRI | 200 | ACTRN12619000864123 |
| CIRCAN | Prevention of recurrent LWRI requiring hospitalization | 2268 | ACTRN12620001370998 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballarini, S.; Rossi, G.A.; Principi, N.; Esposito, S. Dysbiosis in Pediatrics Is Associated with Respiratory Infections: Is There a Place for Bacterial-Derived Products? Microorganisms 2021, 9, 448. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020448
Ballarini S, Rossi GA, Principi N, Esposito S. Dysbiosis in Pediatrics Is Associated with Respiratory Infections: Is There a Place for Bacterial-Derived Products? Microorganisms. 2021; 9(2):448. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020448
Chicago/Turabian StyleBallarini, Stefania, Giovanni A. Rossi, Nicola Principi, and Susanna Esposito. 2021. "Dysbiosis in Pediatrics Is Associated with Respiratory Infections: Is There a Place for Bacterial-Derived Products?" Microorganisms 9, no. 2: 448. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020448