
Morphological and Chemical Traits of Cladonia Respond to Multiple Environmental Factors in Acidic Dry Grasslands

 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
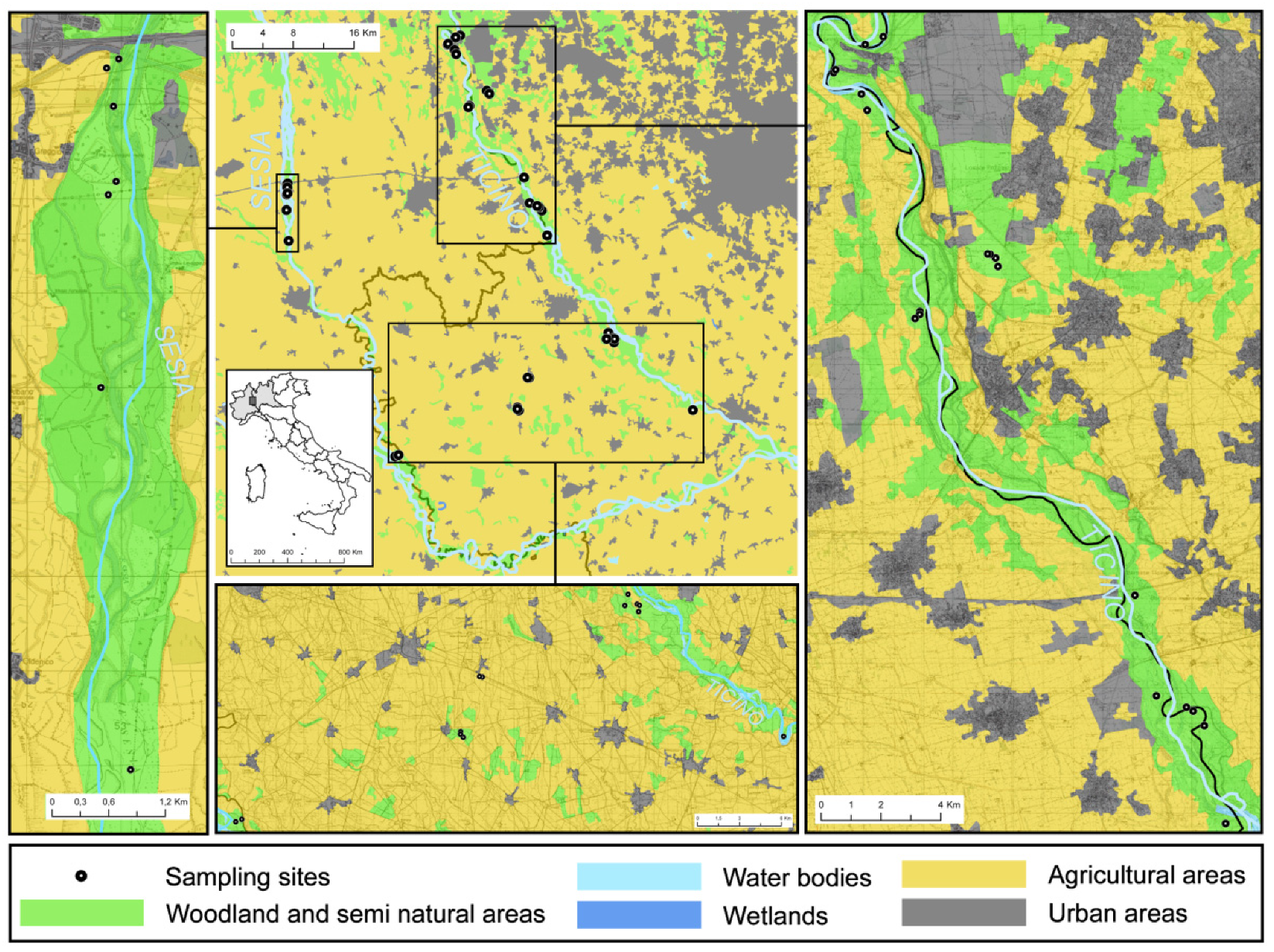
2.1. Study Area
2.2. Sampling
2.3. The Genus Cladonia
2.4. Functional Traits
2.5. Environmental Variables
2.6. Data Analysis
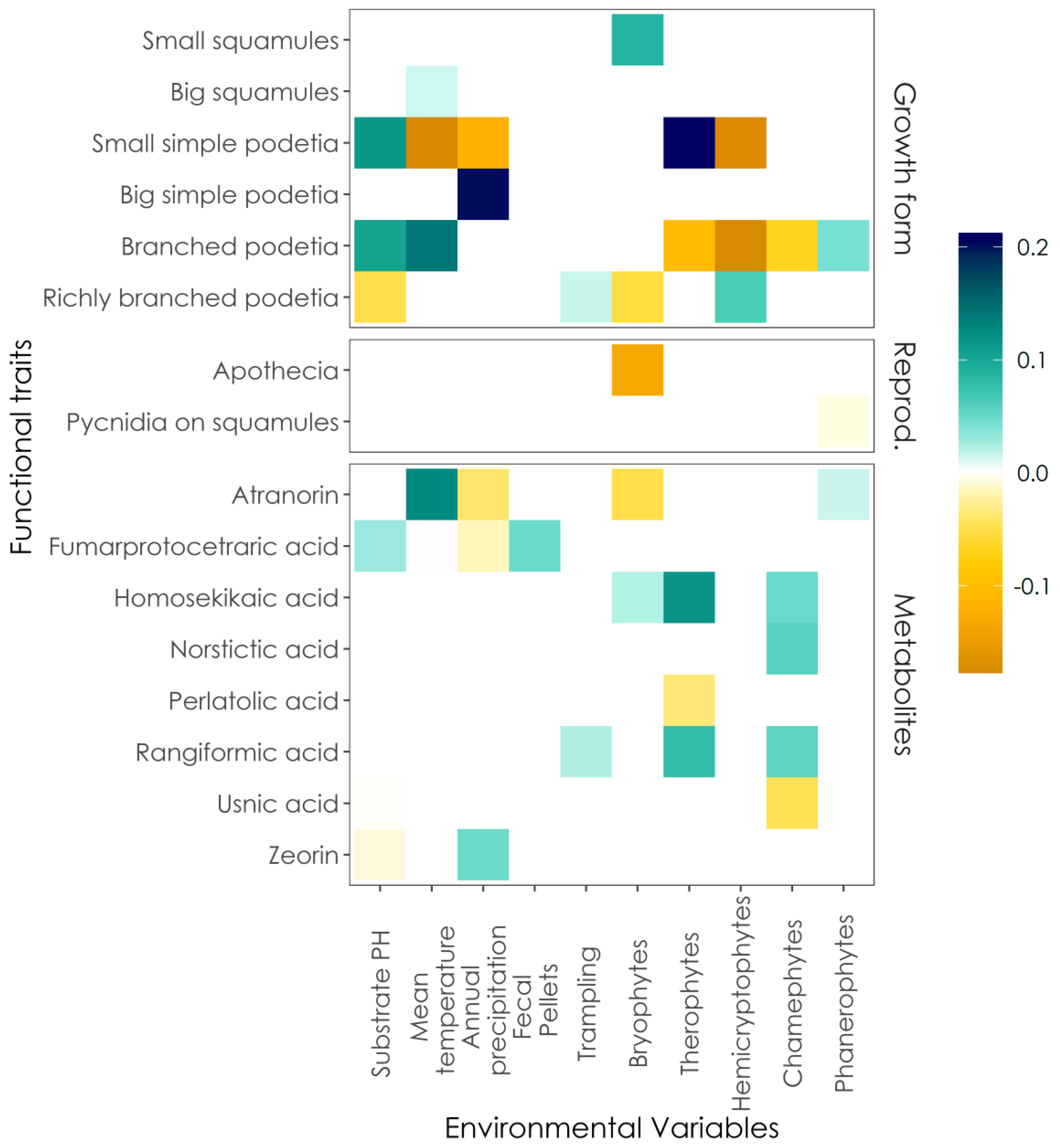
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nimis, P.L.; Martellos, S.; Chiarucci, A.; Ongaro, S.; Peplis, M.; Pittao, E.; Nascimbene, J. Exploring the Relationships between Ecology and Species Traits in Cyanolichens: A Case Study on Italy. Fungal Ecol. 2020, 47, 100950. [Google Scholar] [CrossRef]
- Saiz, H.; Dainese, M.; Chiarucci, A.; Nascimbene, J. Networks of Epiphytic Lichens and Host Trees along Elevation Gradients: Climate Change Implications in Mountain Ranges. J. Ecol. 2020. [Google Scholar] [CrossRef]
- Giordani, P.; Brunialti, G.; Bacaro, G.; Nascimbene, J. Functional Traits of Epiphytic Lichens as Potential Indicators of Environmental Conditions in Forest Ecosystems. Ecol. Indic. 2012, 18, 413–420. [Google Scholar] [CrossRef]
- Jüriado, I.; Paal, J. Epiphytic Lichen Synusiae and Functional Trait Groups in Boreo-Nemoral Deciduous Forests Are Influenced by Host Tree and Environmental Factors. Nord. J. Bot. 2019, 37, e01939. [Google Scholar] [CrossRef]
- Hurtado, P.; Prieto, M.; Martínez-Vilalta, J.; Giordani, P.; Aragón, G.; López-Angulo, J.; Košuthová, A.; Merinero, S.; Díaz-Peña, E.M.; Rosas, T.; et al. Disentangling Functional Trait Variation and Covariation in Epiphytic Lichens along a Continent-Wide Latitudinal Gradient. Proc. R. Soc. B 2020, 287, 20192862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leppik, E.; Jüriado, I.; Suija, A.; Liira, J. Functional Ecology of Rare and Common Epigeic Lichens in Alvar Grasslands. Fungal Ecol. 2015, 13, 66–76. [Google Scholar] [CrossRef]
- Jüriado, I.; Kämärä, M.-L.; Oja, E. Environmental Factors and Ground Disturbance Affecting the Composition of Species and Functional Traits of Ground Layer Lichens on Grey Dunes and Dune Heaths of Estonia. Nord. J. Bot. 2016, 34, 244–255. [Google Scholar] [CrossRef]
- Vallese, C.; Nascimbene, J.; Giordani, P.; Benesperi, R.; Casazza, G. Modelling Range Dynamics of Terricolous Lichens of the Genus Peltigera in the Alps under a Climate Change Scenario. Fungal Ecol. 2021, 49, 101014. [Google Scholar] [CrossRef]
- Maestre, F.T.; Bowker, M.A.; Cantón, Y.; Castillo-Monroy, A.P.; Cortina, J.; Escolar, C.; Escudero, A.; Lázaro, R.; Martínez, I. Ecology and Functional Roles of Biological Soil Crusts in Semi-Arid Ecosystems of Spain. J. Arid Environ. 2011, 75, 1282–1291. [Google Scholar] [CrossRef] [Green Version]
- Gheza, G.; Assini, S.; Lelli, C.; Marini, L.; Mayrhofer, H.; Nascimbene, J. Biodiversity and Conservation of Terricolous Lichens and Bryophytes in Continental Lowlands of Northern Italy: The Role of Different Dry Habitat Types. Biodivers. Conserv. 2020, 29, 3533–3550. [Google Scholar] [CrossRef]
- Burgaz, R.A.; Ahti, T. Cladoniaceae; Flora liquenológica ibérica; Sociedad Española de Liquenología: Madrid, Spain, 2009; ISBN 1696-0513. [Google Scholar]
- Ahti, T.; Stenroos, S.; Moberg, R. Cladoniaceae; Nordic Lichen Flora; Museum of Evolution, Uppsala University: Uppsala, Sweden, 2013; Volume 5. [Google Scholar]
- Gheza, G. Chiave Alle Specie Italiane Del Genere Cladonia. Not. Della Soc. Lichenol. Ital. 2018, 31, 70–96. [Google Scholar]
- Bültmann, H. Zeigerwerte von Erdflechten in Trockenrasen: Vorschläge zur Ergänzung und Korrektur. In Trockenrasen auf Unterschiedlichen Betrachtungsebenen; Bültmann, H., Fartmann, T., Hasse, T., Eds.; Institut fur Landschaftsökologie Münster: Munster, Germany, 2006; Volume 15, pp. 127–143. [Google Scholar]
- Sulavik, J.; Auestad, I.; Halvorsen, R.; Rydgren, K. Assessing Recovery of Alpine Spoil Heaps by Vascular Plant, Bryophyte, and Lichen Functional Traits. Restor. Ecol. 2020. [Google Scholar] [CrossRef]
- Heinken, T. Dispersal Patterns of Terricolous Lichens by Thallus Fragments. Lichenologist 1999, 31, 603–612. [Google Scholar] [CrossRef]
- Molnár, K.; Farkas, E. Current Results on Biological Activities of Lichen Secondary Metabolites: A Review. Z. Für Nat. C 2010, 65, 157–173. [Google Scholar] [CrossRef]
- Stanojkovic, T. Lichen Secondary Metabolites: Bioactive Properties and Pharmaceutical Potential; Ranković, B., Ed.; Springer International Publishing: New York, NY, USA, 2015; ISBN 978-3-319-35516-0. [Google Scholar]
- Goga, M.; Elečko, J.; Marcinčinová, M.; Ručová, D.; Bačkorová, M.; Bačkor, M. Lichen Metabolites: An Overview of Some Secondary Metabolites and Their Biological Potential. In Co-Evolution of Secondary Metabolites; Mérillon, J.-M., Ramawat, K.G., Eds.; Reference Series in Phytochemistry; Springer International Publishing: Cham, Switzerland, 2020; pp. 175–209. ISBN 978-3-319-96397-6. [Google Scholar]
- Park, Y.S. Habitat Selection in a Pair of Sibling Chemospecies of the Lichen Genus Cladonia. Am. Midl. Nat. 1985, 114, 180–183. [Google Scholar] [CrossRef]
- Zraik, M.; Booth, T.; Piercey-Normore, M.D. Relationship between Lichen Species Composition, Secondary Metabolites and Soil PH, Organic Matter, and Grain Characteristics in Manitoba. Botany 2018, 96, 267–279. [Google Scholar] [CrossRef] [Green Version]
- Hauck, M.; Jürgens, S.-R.; Leuschner, C. Norstictic Acid: Correlations between Its Physico-Chemical Characteristics and Ecological Preferences of Lichens Producing This Depsidone. Environ. Exper. Bot. 2010, 68, 309–313. [Google Scholar] [CrossRef]
- Bültmann, H.; Daniels, F.J. Lichen Richness-Biomass Relationship in Terricolous Lichen Vegetation on Non-Calcareous Substrates. Phytocoenologia 2001, 31, 537–570. [Google Scholar] [CrossRef] [Green Version]
- Ketner-Oostra, R.; Sýkora, K.V. Vegetation Change in a Lichen-Rich Inland Drift Sand Area in the Netherlands. Phytocoenologia 2009, 267–286. [Google Scholar] [CrossRef]
- Gheza, G.; Assini, S.; Valcuvia Passadore, M. Terricolous Lichen Communities of Corynephorus Canescens Grasslands of Northern Italy. Tuexenia 2016, 36, 121–142. [Google Scholar] [CrossRef]
- Gheza, G.; Barcella, M.; Assini, S. Terricolous Lichen Communities in Thero-Airion Dry Grasslands of the Po Plain (Northern Italy): Syntaxonomy, Ecology and Conservation Value. Tuexenia 2019, 39, 377–400. [Google Scholar] [CrossRef]
- Janssen, J.A.M.; Rodwell, J.S.; Criado, M.G.; Arts, G.H.P.; Bijlsma, R.J.; Schaminee, J.H.J. European Red List of Habitats: Part 2. Terrestrial and Freshwater Habitats; European Union: Luxembourg, 2016. [Google Scholar]
- Strijker, D. Marginal Lands in Europe—Causes of Decline. Basic Appl. Ecol. 2005, 6, 99–106. [Google Scholar] [CrossRef]
- Assini, S. Habitat 2330 (Inland Dunes with Open Corynephorus and Agrostis Grasslands): Problematiche Di Conservazione e Ipotesi Di Intervento (Habitat 2330 (Inland Dunes with Open Corynephorus and Agrostis Grasslands): Conservation Problems and Intervention Hypotheses). Arch. Geobot. 2008, 14, 23–28. [Google Scholar]
- Assini, S.; Mondino, G.P.; Varese, P.; Barcella, M.; Bracco, F. A Phytosociological Survey of the Corynephorus Canescens (L.) Beauv. Communities of Italy. Plant. Biosyst. 2013, 147, 64–78. [Google Scholar] [CrossRef]
- Leppik, E.; Jüriado, I.; Suija, A.; Liira, J. The Conservation of Ground Layer Lichen Communities in Alvar Grasslands and the Relevance of Substitution Habitats. Biodivers. Conserv. 2013, 22, 591–614. [Google Scholar] [CrossRef]
- Jentsch, A.; Beyschlag, W. Vegetation Ecology of Dry Acidic Grasslands in the Lowland Area of Central Europe. Flora 2003, 198, 3–25. [Google Scholar] [CrossRef]
- Gheza, G.; Assini, S.; Marini, L.; Nascimbene, J. Impact of an Invasive Herbivore and Human Trampling on Lichen-Rich Dry Grasslands: Soil-Dependent Response of Multiple Taxa. Sci. Total Environ. 2018, 639, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Gheza, G. Terricolous Lichens of the Western Padanian Plain: New Records of Phytogeographical Interest. Acta Bot. Gall. 2015, 162, 339–348. [Google Scholar] [CrossRef]
- Gheza, G. I Licheni Terricoli Degli Ambienti Aperti Aridi Della Pianura Piemontese. Riv. Piemont. Stor. Nat. 2020, 41, 23–31. [Google Scholar]
- Lange, O.L.; Green, T.G.A.; Türk, R. An Unusual Growth Form of Cladonia Furcata: The Trampling-Resistant Primary Thallus Colonizing a Paved Pathway. Lichenologist 1998, 30, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Huovinen, K.; Ahti, T. Biosequential Patterns for the Formation of Depsides, Depsidones and Dibenzofurans in the Genus Cladonia (Lichen-Forming Ascomycetes). Ann. Bot. Fenn. 1982, 19, 225–234. [Google Scholar]
- Huovinen, K.; Ahti, T. The Composition and Contents of Aromatic Lichen Substances in the Genus Cladina. Ann. Bot. Fenn. 1986, 23, 93–106. [Google Scholar]
- Huovinen, K.; Ahti, T. The Composition and Contents of Aromatic Lichen Substances in Cladonia, Section Unciales. Ann. Bot. Fenn. 1986, 23, 173–188. [Google Scholar]
- Huovinen, K.; Ahti, T. The Composition and Contents of Aromatic Lichen Substances in Cladonia Section Perviae. Ann. Bot. Fenn. 1988, 25, 371–383. [Google Scholar]
- Huovinen, K.; Ahti, T.; Stenroos, S. The Composition and Contents of Aromatic Lichen Substances in Cladonia, Section Cocciferae. Ann. Bot. Fenn. 1989, 26, 133–148. [Google Scholar]
- Huovinen, K.; Ahti, T.; Stenroos, S. The Composition and Contents of Aromatic Lichen Substances in Cladonia Section Helopodium and Subsection Foliosae. Ann. Bot. Fenn. 1989, 26, 297–306. [Google Scholar]
- Huovinen, K.; Ahti, T.; Stenroos, S. The Composition and Contents of Aromatic Lichen Substances in Cladonia Section Cladonia and Group Furcatae. Bibl. Lichenol. 1990, 209–241. Available online: https://www.cabdirect.org/?target=%2fcabdirect%2fabstract%2f20198610189 (accessed on 26 January 2021).
- Lokajová, V.; Bačkorová, M.; Bačkor, M. Allelopathic Effects of Lichen Secondary Metabolites and Their Naturally Occurring Mixtures on Cultures of Aposymbiotically Grown Lichen Photobiont Trebouxia Erici (Chlorophyta). S. Afr. J. Bot. 2014, 93, 86–91. [Google Scholar] [CrossRef] [Green Version]
- Peres, M.T.; Cândido, A.C.S.; Faccenda, O.; Gianini, A.S.; Honda, N.K.; Peres, M.T.; Cândido, A.C.S.; Faccenda, O.; Gianini, A.S.; Honda, N.K. Phytotoxicity of Perlatolic Acid and Derivatives. Braz. Arch. Biol. Technol. 2016, 59. [Google Scholar] [CrossRef] [Green Version]
- Goga, M.; Antreich, S.J.; Bačkor, M.; Weckwerth, W.; Lang, I. Lichen Secondary Metabolites Affect Growth of Physcomitrella Patens by Allelopathy. Protoplasma 2017, 254, 1307–1315. [Google Scholar] [CrossRef]
- Slansky, F., Jr. Effect of the Lichen Chemicals Atranorin and Vulpinic Acid upon Feeding and Growth of Larvae of the Yellow-Striped Armyworm, Spodoptera Ornithogalli. Environ. Entomol. 1979, 8, 865–868. [Google Scholar] [CrossRef]
- Cook, W.E.; Raisbeck, M.F.; Cornish, T.E.; Williams, E.S.; Brown, B.; Hiatt, G.; Kreeger, T.J. Paresis and Death in Elk (Cervus Elaphus) Due to Lichen Intoxication in Wyoming. J. Wildl. Dis. 2007, 43, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Cetin, H.; Tufan-Cetin, O.; Turk, A.O.; Tay, T.; Candan, M.; Yanikoglu, A.; Sumbul, H. Insecticidal Activity of Major Lichen Compounds, (-)- and (+)-Usnic Acid, against the Larvae of House Mosquito, Culex Pipiens L. Parasitol. Res. 2008, 102, 1277–1279. [Google Scholar] [CrossRef] [PubMed]
- Nybakken, L.; Julkunen-Tiitto, R. UV-B Induces Usnic Acid in Reindeer Lichens. Lichenologist 2006, 38, 477–485. [Google Scholar] [CrossRef]
- Millot, M.; Di Meo, F.; Tomasi, S.; Boustie, J.; Trouillas, P. Photoprotective Capacities of Lichen Metabolites: A Joint Theoretical and Experimental Study. J. Photochem. Photobiol. B 2012, 111, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Farkas, E.; Biró, B.; Szabó, K.; Veres, K.; Csintalan, Z.; Engel, R. The Amount of Lichen Secondary Metabolites in Cladonia Foliacea (Cladoniaceae, Lichenised Ascomycota). Acta Bot. Hung. 2020, 62, 33–48. [Google Scholar] [CrossRef]
- Hauck, M.; Jürgens, S.-R.; Huneck, S.; Leuschner, C. High Acidity Tolerance in Lichens with Fumarprotocetraric, Perlatolic or Thamnolic Acids Is Correlated with Low PKa1 Values of These Lichen Substances. Environ. Pollut. 2009, 157, 2776–2780. [Google Scholar] [CrossRef]
- James, P.W.; Cladonia, P.B. Lichens of Great Britain and Ireland; Smith, C.W., Aptroot, A., Coppins, B.J., Fletcher, A., Gilbert, O.L., James, P.W., Wolseley, P.A., Eds.; The British Lichen Society & The Natural History Museum: London, UK, 2009; pp. 309–338. [Google Scholar]
- Nimis, P.L.; Martellos, S. ITALIC 6.0—The Information System on Italian Lichens. Available online: http://dryades.units.it/italic (accessed on 20 January 2021).
- Elix, J.A. A Catalogue of Standardized Chromatographic Data and Biosynthetic Relationships for Lichen Substances; The Author: Canberra, Australia, 2014. [Google Scholar]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Frey, W.; Lösch, R. Geobotanik—Pflanze und Vegetation in Raum und Zeit, 3rd ed.; Springer Spektrum: Wiesbaden, Germany, 2010; ISBN 978-3-8274-2335-1. [Google Scholar]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at High Resolution for the Earth’s Land Surface Areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef] [Green Version]
- Dray, S.; Legendre, P. Testing the Species Traits–Environment Relationships: The Fourth-Corner Problem Revisited. Ecology 2008, 89, 3400–3412. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.M.; Warton, D.I.; Andrew, N.R.; Binns, M.; Cassis, G.; Gibb, H. The Fourth-Corner Solution—Using Predictive Models to Understand How Species Traits Interact with the Environment. Methods Ecol. Evol. 2014, 5, 344–352. [Google Scholar] [CrossRef]
- Wang, Y.; Naumann, U.; Eddelbuettel, D.; Wilshire, J.; Warton, D.; Byrnes, J.; dos Santos Silva, R.; Niku, J.; Renner, I.; Wright, S. Mvabund: Statistical Methods for Analysing Multivariate Abundance Data; R Package Version 4.1.3.; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Rao, D.N.; LeBlanc, F. A Possible Role of Atranorin in the Lichen Thallus. Bryologist 1965, 68, 284–289. [Google Scholar] [CrossRef]
- Paus, S. Die Erdflechtenvegetation Nordwestdeutschlands Und Einiger-Randgebiete. Bibl. Lichenol. 1997, 66, 1–222. [Google Scholar]
- Osyczka, P.; Rola, K. Cladonia Lichens as the Most Effective and Essential Pioneers in Strongly Contaminated Slag Dumps. Cent. Eur. J. Biol. 2013, 8, 876–887. [Google Scholar] [CrossRef]
- Rola, K.; Osyczka, P.; Nobis, M. Cryptogamic Communities Dominated by the Lichen Cladonia Rei—A Case Study of Polish Post-Smelting Dumps in a Worldwide Context. Herzogia 2014, 27, 121–135. [Google Scholar] [CrossRef]
- Benesperi, R.; Nascimbene, J.; Lazzaro, L.; Bianchi, E.; Tepsich, A.; Longinotti, S.; Giordani, P. Successful Conservation of the Endangered Forest Lichen Lobaria Pulmonaria Requires Knowledge of Fine-Scale Population Structure. Fungal Ecol. 2018, 33, 65–71. [Google Scholar] [CrossRef]
- Bayfield, N.G.; Urquhart, U.H.; Cooper, S.M. Susceptibility of Four Species of Cladonia to Disturbance by Trampling in the Cairngorm Mountains, Scotland. J. Appl. Ecol. 1981, 18, 303–310. [Google Scholar] [CrossRef]
- Heggenes, J.; Odland, A.; Chevalier, T.; Ahlberg, J.; Berg, A.; Larsson, H.; Bjerketvedt, D.K. Herbivore Grazing—or Trampling? Trampling Effects by a Large Ungulate in Cold High-Latitude Ecosystems. Ecol. Evol. 2017, 7, 6423–6431. [Google Scholar] [CrossRef] [Green Version]
- Hauck, M.; Huneck, S. The Putative Role of Fumarprotocetraric Acid in the Manganese Tolerance of the Lichen Lecanora Conizaeoides. Lichenologist 2007, 39, 301–304. [Google Scholar] [CrossRef]
- Ranković, B.; Mišić, M. The Antimicrobial Activity of the Lichen Substances of the Lichens Cladonia Furcata, Ochrolechia Androgyna, Parmelia Caperata and Parmelia Conspresa. Biotechnol. Biotechnol. Equip. 2008, 22, 1013–1016. [Google Scholar] [CrossRef] [Green Version]
- Aptroot, A.; van Herk, C.M. Further Evidence of the Effects of Global Warming on Lichens, Particularly Those with Trentepohlia Phycobionts. Environ. Pollut. 2007, 146, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Concostrina-Zubiri, L.; Valencia, E.; Ochoa, V.; Gozalo, B.; Mendoza, B.J.; Maestre, F.T. Species-Specific Effects of Biocrust-Forming Lichens on Soil Properties under Simulated Climate Change Are Driven by Functional Traits. New Phytol. 2020. [Google Scholar] [CrossRef]
- Gauslaa, Y. Rain, Dew, and Humid Air as Drivers of Morphology, Function and Spatial Distribution in Epiphytic Lichens. Lichenologist 2014, 46, 1–16. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Abbreviation | Description |
|---|---|
| Small squamules | Squamulose thallus with small squamules: species usually without podetia and with squamules usually shorter than 5 mm; e.g., Cladonia strepsilis. |
| Big squamules | Squamulose thallus with big squamules: species usually without podetia and with squamules usually longer than 5 mm; e.g., Cladonia foliacea. |
| Small simple podetia | Thallus with small simple podetia: species with persistent primary thallus and usually with bacillar or capitiform podetia shorter than 10 mm; e.g., Cladonia peziziformis. |
| Big simple podetia | Thallus with big simple podetia: species with persistent or ephemeral primary thallus and usually with bacillar of scyphipherous podetia taller than 10 mm; e.g., Cladonia pyxidata, Cladonia rei. |
| Branched podetia | Thallus with branched podetia: species with ephemeral primary thallus and with sparingly branched podetia; e.g., Cladonia furcata. |
| Richly branched podetia | Thallus with richly branched podetia: species with ephemeral primary thallus and with richly branched, bush-shaped podetia; e.g., Cladonia rangiformis and species belonging to subgenus Cladina. |
| Species | Growth Form | Position of Pycnidia | Reproduction | Metabolites |
|---|---|---|---|---|
| Cladonia cariosa (Ach.) Spreng. | Small simple podetia | Squamules | Spores | Atranorin, rangiformic acid |
| Cladonia chlorophaea (Sommerf.) Spreng. | Big simple podetia | Podetia | Soredia | Fumarprotocetraric acid |
| Cladonia coccifera (L.) Willd. | Big simple podetia | Podetia | Spores | Usnic acid, zeorin |
| Cladonia fimbriata (L.) Fr. | Big simple podetia | Podetia | Soredia | Fumarprotocetraric acid |
| Cladonia foliacea (Huds.) Willd. | Big squamules | Squamules | Spores | Fumarprotocetraric acid, usnic acid |
| Cladonia furcata (Huds.) Schrad. | Branched podetia | Podetia | Spores | Atranorin, fumarprotocetraric acid |
| Cladonia peziziformis (With.) J.R.Laundon | Small simple podetia | Squamules | Spores | Fumarprotocetraric acid |
| Cladonia polycarpoides Nyl. | Big squamules | Squamules | Spores | Norstictic acid |
| Cladonia portentosa (Dufour) Coem. | Richly branched podetia | Podetia | Spores | Perlatolic acid, usnic acid |
| Cladonia pyxidata (L.) Hoffm. | Big simple podetia | Podetia | Spores | Fumarprotocetraric acid |
| Cladonia rangiformis Hoffm. | Richly branched podetia | Podetia | Spores | Atranorin, rangiformic acid |
| Cladonia rei Schaer. | Big simple podetia | Podetia | Soredia | Fumarprotocetraric acid, homosekikaic acid |
| Cladonia strepsilis (Ach.) Grognot | Small squamules | Squamules | Spores | Baeomycesic acid, squamatic acid, strepsilin |
| Cladonia uncialis (L.) F.H.Wigg. | Branched podetia | Podetia | Spores | Usnic acid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gheza, G.; Di Nuzzo, L.; Vallese, C.; Barcella, M.; Benesperi, R.; Giordani, P.; Nascimbene, J.; Assini, S. Morphological and Chemical Traits of Cladonia Respond to Multiple Environmental Factors in Acidic Dry Grasslands. Microorganisms 2021, 9, 453. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020453
Gheza G, Di Nuzzo L, Vallese C, Barcella M, Benesperi R, Giordani P, Nascimbene J, Assini S. Morphological and Chemical Traits of Cladonia Respond to Multiple Environmental Factors in Acidic Dry Grasslands. Microorganisms. 2021; 9(2):453. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020453
Chicago/Turabian StyleGheza, Gabriele, Luca Di Nuzzo, Chiara Vallese, Matteo Barcella, Renato Benesperi, Paolo Giordani, Juri Nascimbene, and Silvia Assini. 2021. "Morphological and Chemical Traits of Cladonia Respond to Multiple Environmental Factors in Acidic Dry Grasslands" Microorganisms 9, no. 2: 453. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020453