
Native Vineyard Non-Saccharomyces Yeasts Used for Biological Control of Botrytis cinerea in Stored Table Grape

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast and Pathogen Isolation and Culture Conditions
2.2. Molecular Identifications
2.3. In vitro Assay
2.4. Extracellular Lytic Enzymes Activity
2.5. Bioassays Antagonistic Activity
2.6. Statistical Analysis
3. Results
3.1. Yeast Isolates and Molecular Identification
3.2. Extracellular Lytic Enzymes Activity
3.3. In Vitro Assay
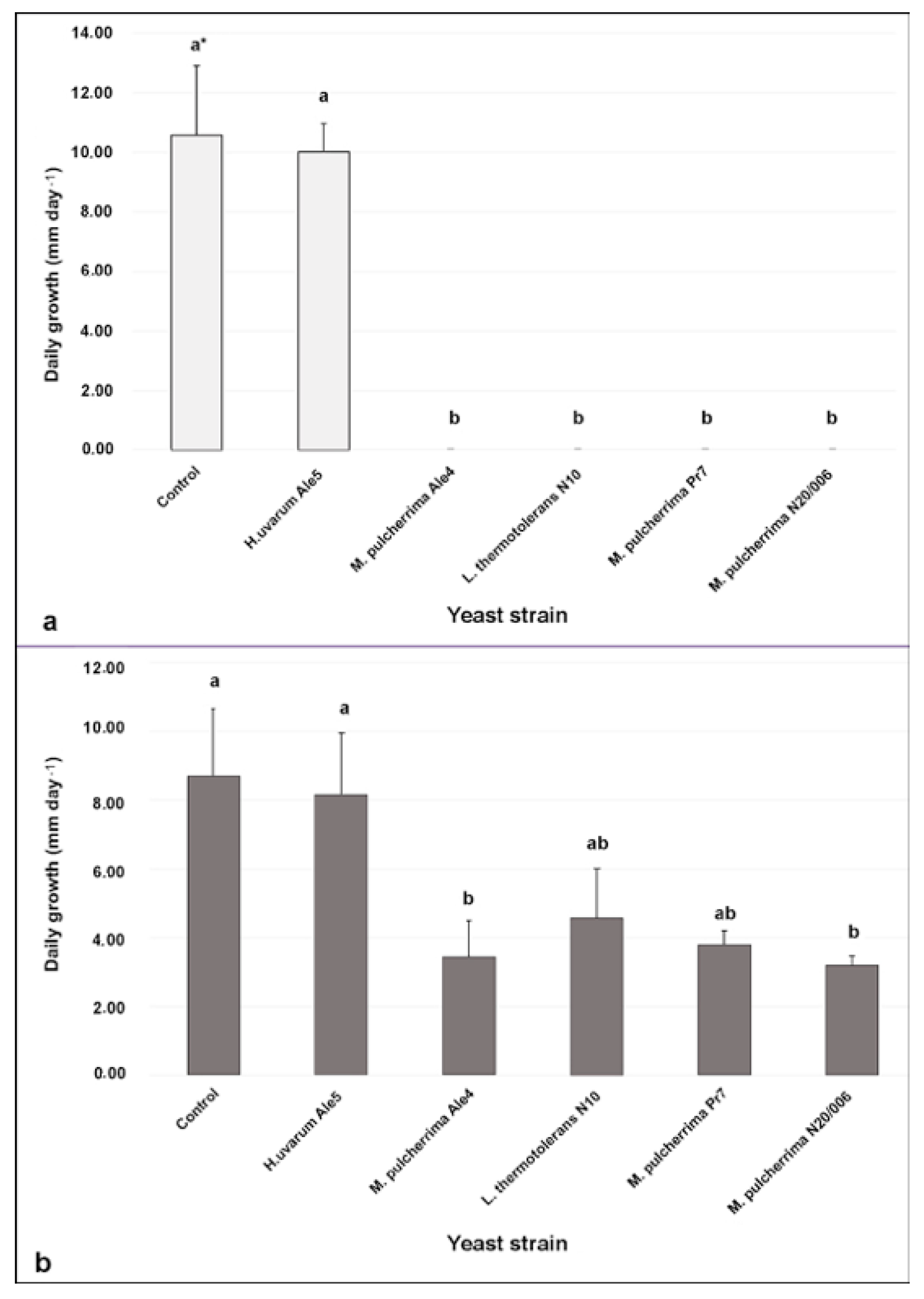
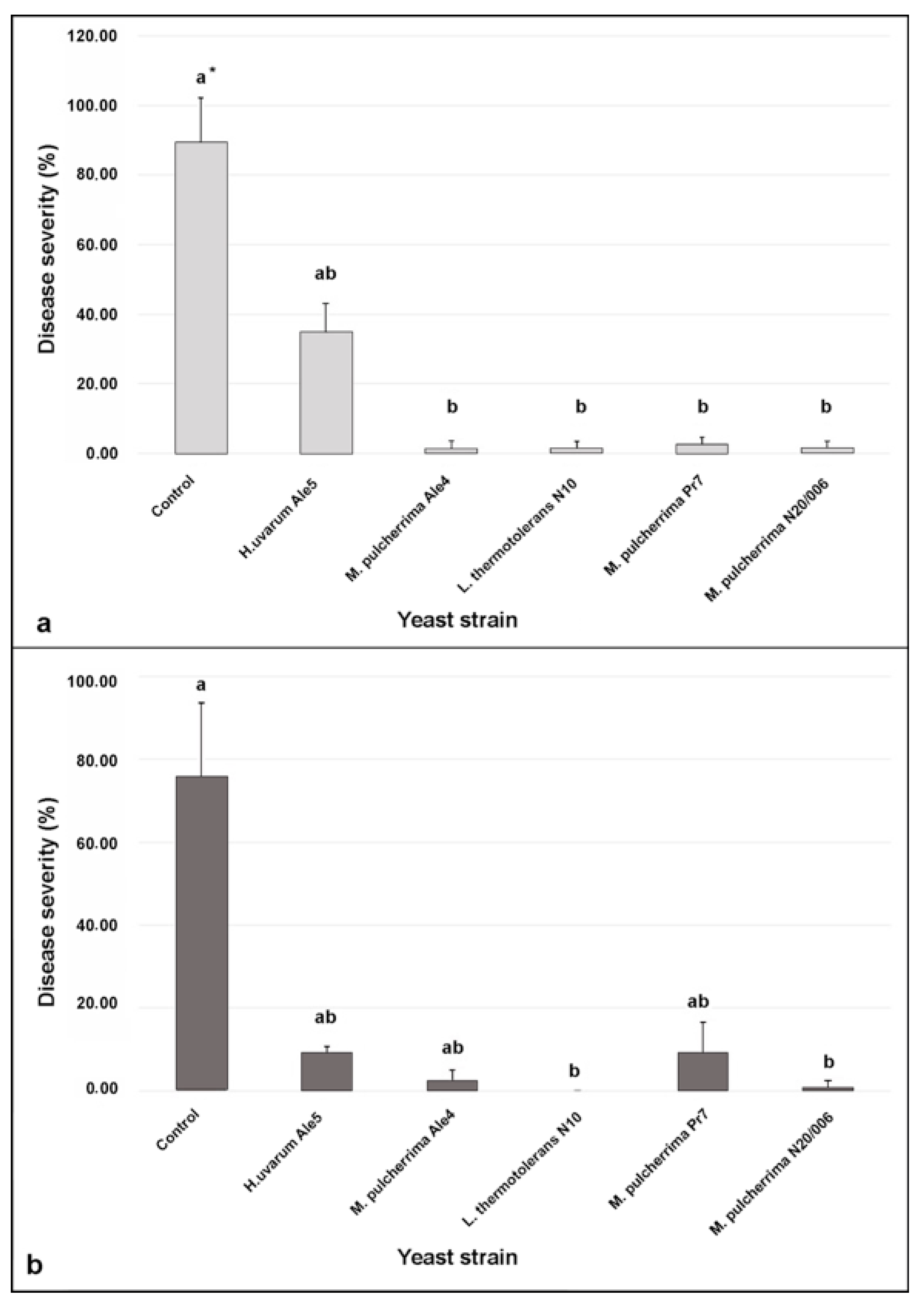
3.4. Efficacy of the Yeast Cells in Controlling Botrytis Bunch Rot in Wounded Berries
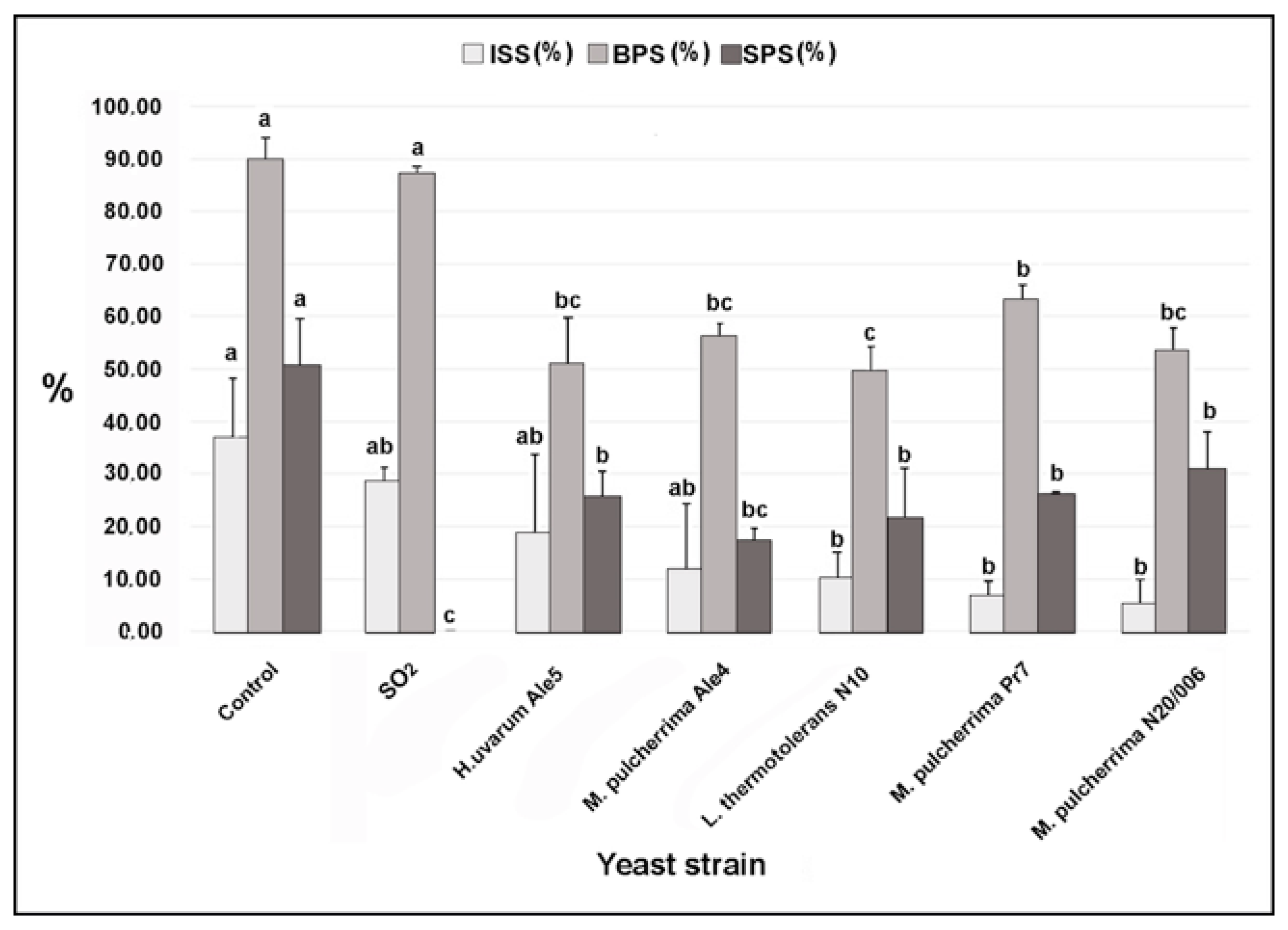
3.5. Efficacy of the Yeast Cells in Controlling Botrytis Bunch Rot in Semi-Commercial Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- ISTAT. ISTAT 2019. 2019. ISBN 978-88-458-2003-8. Available online: https://www.istat.it/it/files//2019/12/Asi-2019.pdf (accessed on 22 February 2021).
- Lydakis, D.; Aked, J. Vapour heat treatment of Sultanina table grapes. II: Effects on postharvest quality. Postharvest Biol. Technol. 2003, 27, 117–126. [Google Scholar] [CrossRef]
- Pinto, L.; Malfeito-Ferreira, M.; Quintieri, L.; Silva, A.; Baruzzi, F. Growth and metabolite production of a grape sour rot yeast-bacterium consortium on different carbon sources. Int. J. Food Microbiol. 2019, 296, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.; Caputo, L.; Quintieri, L.; de Candia, S.; Baruzzi, F. Efficacy of gaseous ozone to counteract postharvest table grape sour rot. Food Microbiol. 2017, 66, 190–198. [Google Scholar] [CrossRef]
- Droby, S.; Lichter, A. Postharvest Botrytis infection: Etiology, development and management. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Kluwer Academic Publishers: London, UK, 2004; pp. 349–367. [Google Scholar]
- Staples, R.; Mayer, A. Putative virulence factors of Botrytis cinerea acting as a wound pathogen. FEMS Microbiol. Lett. 1995, 134, 1–7. [Google Scholar] [CrossRef]
- Jacometti, M.A.; Wratten, S.D.; Walter, M. Review: Alternatives to synthetic fungicides for Botrytis cinerea management in vineyards. Aust. J. Grape Wine Res. 2010, 16, 154–172. [Google Scholar] [CrossRef]
- Ahmed, S.; Roberto, S.R.; Domingues, A.R.; Shahab, M.; Junior, O.J.C.; Sumida, C.H.; De Souza, R.T. Effects of Different Sulfur Dioxide Pads on Botrytis Mold in ‘Italia’ Table Grapes under Cold Storage. Horticulturae 2018, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- Mustonen, H. The efficacy of a range of sulfur dioxide generating pads against Botrytis cinerea infection and on out-turn quality of Calmeria table grapes. Aust. J. Exp. Agric. 1992, 32, 389–393. [Google Scholar] [CrossRef]
- Romanazzi, G.; Feliziani, E.; Baños, S.B.; Sivakumar, D. Shelf life extension of fresh fruit and vegetables by chitosan treatment. Crit. Rev. Food Sci. Nutr. 2017, 57, 579–601. [Google Scholar] [CrossRef]
- Romanazzi, G.; Lichter, A.; Gabler, F.M.; Smilanick, J.L. Recent advances on the use of natural and safe alternatives to conventional methods to control postharvest gray mold of table grapes. Postharvest Biol. Technol. 2012, 63, 141–147. [Google Scholar] [CrossRef]
- Candir, E.; Ozdemir, A.E.; Kamiloglu, O.; Soylu, E.M.; Dilbaz, R.; Ustun, D. Modified atmosphere packaging and ethanol vapor to control decay of ‘Red Globe’ table grapes during storage. Postharvest Biol. Technol. 2012, 63, 98–106. [Google Scholar] [CrossRef]
- Beardsley, P.M.; Hauser, K.F. Glial modulators as potential treatments of psychostimulant abuse. Adv. Pharmacol. 2014, 69, 1–69. [Google Scholar] [CrossRef] [Green Version]
- Chanchaichaovivat, A.; Ruenwongsa, P.; Panijpan, B. Screening and identification of yeast strains from fruits and vegeta-bles: Potential for biological control of postharvest chilli anthracnose (Colletotrichum capsici). Biol. Control 2007, 42, 326–335. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Sivasithamparam, K. Potential of yeasts as biocontrol agents of soil-borne fungal plant pathogens and as plant growth promoters. Mycoscience 2006, 47, 25–35. [Google Scholar] [CrossRef]
- Nunes, C.A. Biological control of postharvest diseases of fruit. Eur. J. Plant Pathol. 2011, 133, 181–196. [Google Scholar] [CrossRef]
- De Simone, N.; Pace, B.; Grieco, F.; Chimienti, M.; Tyibilika, V.; Santoro, V.; Capozzi, V.; Colelli, G.; Spano, G.; Russo, P. Botrytis cinerea and Table Grapes: A Review of the Main Physical, Chemical, and Bio-Based Control Treatments in Post-Harvest. Foods 2020, 9, 1138. [Google Scholar] [CrossRef] [PubMed]
- Leneveu-Jenvrin, C.; Charles, F.; Barba, F.J.; Remize, F. Role of biological control agents and physical treatments in main-taining the quality of fresh and minimally-processed fruit and vegetables. Crit. Rev. Food Sci. Nutr. 2020, 60, 2837–2855. [Google Scholar] [CrossRef] [PubMed]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit: The importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Macarisin, D.; Wilson, C. Twenty years of postharvest biocontrol research: Is it time for a new paradigm? Postharvest Biol. Technol. 2009, 52, 137–145. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Korsten, L. Biological control of postharvest diseases of fruits. Annu. Rev. Phytopathol. 2002, 40, 411–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eder, M.L.R.; Reynoso, C.; Lauret, S.C.; Rosa, A.L. Isolation and Identification of the Indigenous Yeast Population during Spontaneous Fermentation of Isabella (Vitis labrusca L.) Grape Must. Front. Microbiol. 2017, 8, 532. [Google Scholar] [CrossRef] [Green Version]
- Pallmann, C.L.; Brown, J.A.; Olineka, T.L.; Cocolin, L.; Mills, D.A.; Bisson, L.F. Use of WL medium t profile native flora fermetations. Am. J. Enol. Vitic. 2001, 52, 6. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Shinsky, J., White, T., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Valente, P.; Gouveia, F.C.; de Lemos, G.A.; Pimentel, D.; van Elsas, J.D.; Mendonça-Hagler, L.C.; Hagler, A.N. PCR amplifi-cation of the rDNA internal transcribed spacer region for differentiation of Saccharomyces cultures. FEMS Microbiol. Lett. 1996, 137, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Valente, P.; Gouveia, F.C.; Lemos, G.A.; Pimentel, D.; Mendonca-Hagler, L.C.; Hagler, A.N. PCR-amplified ITS length varia-tion within the yeast genus Metschnikowia. J. Gen. Appl. Microbiol. 1997, 43, 179–181. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Li, M.; Zhan, Y.; Ban, X.; Zeng, T.; Zhu, Y.; Yun, J.; Guan, X.-Y.; Li, Y. Down-regulation of POTEG predicts poor prognosis in esophageal squamous cell carcinoma patients. Mol. Carcinog. 2018, 57, 886–895. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, J.N. A Study of the Epiphytic Growth Habit of Fomes annosus. Ann. Bot. 1967, 31, 755–774. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Biocontrol ability and action mechanism of food-isolated yeast strains against Botrytis cinerea causing post-harvest bunch rot of table grape. Food Microbiol. 2015, 47, 85–92. [Google Scholar] [CrossRef]
- Roberts, W.K.; Selitrennikoff, C.P. Plant and Bacterial Chitinases Differ in Antifungal Activity. Microbiology 1988, 134, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Souza, C.P.; Burbano-Rosero, E.M.; Almeida, B.C.; Martins, G.G.; Albertini, L.S.; Rivera, I.N.G. Culture medium for isolating chitinolytic bacteria from seawater and plankton. World J. Microbiol. Biotechnol. 2009, 25, 2079–2082. [Google Scholar] [CrossRef]
- Renwick, A.; Campbell, R.; Coe, S. Assessment of in vivo screening systems for potential biocontrol agents of Gaeuman-nomyces graminis. Plant Pathol. 1991, 40, 524–532. [Google Scholar] [CrossRef]
- Buzzini, P.; Martini, A. Extracellular enzymatic activity profiles in yeast and yeast-like strains isolated from tropical envi-ronments. J. Appl. Microbiol. 2002, 93, 1020–1025. [Google Scholar] [CrossRef] [PubMed]
- Strauss, M.L.; Jolly, N.P.; Lambrechts, M.G.; van Rensburg, P. Screening for the production of extracellular hydrolytic en-zymes by non-Saccharomyces wine yeasts. J. Appl. Microbiol. 2001, 91, 182–190. [Google Scholar] [CrossRef]
- Townsend, G.R.; Heuberger, J.W. Methods for estimating losses caused by diseases in fungicidal experiments. Plant Dis. Report. 1943, 27, 4. [Google Scholar]
- Servili, A.; Feliziani, E.; Romanazzi, G. Exposure to volatiles of essential oils alone or under hypobaric treatment to control postharvest gray mold of table grapes. Postharvest Biol. Technol. 2017, 133, 36–40. [Google Scholar] [CrossRef]
- Tristezza, M.; Vetrano, C.; Bleve, G.; Spano, G.; Capozzi, V.; Logrieco, A.; Mita, G.; Grieco, F. Biodiversity and safety aspects of yeast strains characterized from vineyards and spontaneous fermentations in the Apulia Region, Italy. Food Microbiol. 2013, 36, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Boland, G.J. Biological control of plant diseases with fungal antagonists: Challenges and opportunities. Can. J. Plant Pathol. 1990, 12, 295–299. [Google Scholar] [CrossRef]
- Liu, H.; Guo, J.; Luo, L.; Liu, P.; Wang, B.; Cheng, Y.; Deng, B.; Long, C. Improvement of Hanseniaspora uvarum biocontrol activity against gray mold by the addition of ammonium molybdate and the possible mechanisms involved. Crop. Prot. 2010, 29, 277–282. [Google Scholar] [CrossRef]
- Usall, J.; Sanchis, V. Tolerance of Penicillium expansum to postharvest fungicide treatments in apple packingshouses in Lerida (Spain). Mycopathol. 1991, 113, 15–18. [Google Scholar] [CrossRef]
- Holmes, G.J.; Eckert, J.W. Sensitivity of Penicillium digitatum and P. italicum to Postharvest Citrus Fungicides in California. Phytopathology 1999, 89, 716–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Bardas, G.; Veloukas, T.; Koutita, O.; Karaoglanidis, G.S. Multiple resistance of Botrytis cinerea from kiwifruit to SDHIs, QoIs and fungicides of other chemical groups. Pest Manag. Sci. 2010, 66, 967–973. [Google Scholar] [CrossRef]
- Angelini, R.M.D.M.; Rotolo, C.; Masiello, M.; Gerin, D.; Pollastro, S.; Faretra, F. Occurrence of fungicide resistance in populations of Botryotinia fuckeliana (Botrytis cinerea ) on table grape and strawberry in southern Italy. Pest Manag. Sci. 2014, 70, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, A.; Castello, I.; Cirvilleri, G.; Perrone, G.; Epifani, F.; Ferrara, M.; Polizzi, G.; Walters, D.R.; Vitale, A. Detection of Botrytis cinerea field isolates with multiple fungicide resistance from table grape in Sicily. Crop. Prot. 2015, 77, 65–73. [Google Scholar] [CrossRef]
- Gabler, F.M.; Smilanick, J.L. Postharvest control of table grape gray mold on detached berries with carbonate and bicar-bonate salts and disinfectants. Am. J. Enol. Vitic. 2001, 52, 9. [Google Scholar]
- Cordero-Bueso, G.; Mangieri, N.; Maghradze, D.; Foschino, R.; Valdetara, F.; Cantoral, J.M.; Vigentini, I. Wild Grape-Associated Yeasts as Promising Biocontrol Agents against. Front. Microbiol. 2017, 8, 2025. [Google Scholar] [CrossRef] [Green Version]
- Manso, T.; Nunes, C. Metschnikowia andauensis as a new biocontrol agent of fruit postharvest diseases. Postharvest Biol. Technol. 2011, 61, 64–71. [Google Scholar] [CrossRef]
- Qin, X.; Xiao, H.; Xue, C.; Yu, Z.; Yang, R.; Cai, Z.; Si, L. Biocontrol of gray mold in grapes with the yeast Hanseniapora uvarum alone and in combination with salicylic acid or sodium bicarbonate. Postharvest Biol. Technol. 2015, 100, 8. [Google Scholar] [CrossRef]
- Doss, R.P.; Potter, S.W.; Soeldner, A.H.; Christian, J.K.; E Fukunaga, L. Adhesion of germlings of Botrytis cinerea. Appl. Environ. Microbiol. 1995, 61, 260–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-ad, N.L.; Bar-Nun, N.; Mayer, A.M. The possible function of the glucan sheath of Botrytis cinerea: Effects on the distri-bution of enzyme activities. FEMS Microbiol. Lett. 2001, 199, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Doss, R.P. Composition and Enzymatic Activity of the Extracellular Matrix Secreted by Germlings of Botrytis cinerea. Appl. Environ. Microbiol. 1999, 65, 404–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Volatile organic compounds from Wickerhamomyces anomalus, Metschnikowia pulcherrima and Saccharomyces cerevisiae inhibit growth of decay causing fungi and control postharvest diseases of strawberries. Int. J. Food Microbiol. 2018, 265, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Junior, W.J.L.; Binati, R.L.; Felis, G.E.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Volatile organic compounds from Starmerella bacillaris to control gray mold on apples and modulate cider aroma profile. Food Microbiol. 2020, 89, 103446. [Google Scholar] [CrossRef] [PubMed]
- Janisiewicz, W.J.; Tworkoski, T.J.; Kurtzman, C.P. Biocontrol Potential of Metchnikowia pulcherrima Strains against Blue Mold of Apple. Phytopathology 2001, 91, 1098–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Selection and evaluation of new antagonists for their efficacy against postharvest brown rot of peaches. Postharvest Biol. Technol. 2010, 55, 174–181. [Google Scholar] [CrossRef]
- Zhang, D.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Potential biocontrol activity of a strain of Pichia guilliermondii against grey mold of apples and its possible modes of action. Biol. Control. 2011, 57, 193–201. [Google Scholar] [CrossRef]
- Piano, S.; Neyrotti, V.; Migheli, Q.; Gullino, M.L. Biocontrol capability of Metschnikowia pulcherrima aganist Botrytis postharvest rot of apple. Postharvest Biol. Technol. 1997, 11, 10. [Google Scholar] [CrossRef]
- Spadaro, D.; Vola, R.; Piano, S.; Gullino, M.L. Mechanisms of action and efficacy of four isolates of the yeast Metschnikowia pulcherrima active against postharvest pathogens on apples. Postharvest Biol. Technol. 2002, 24, 123–134. [Google Scholar] [CrossRef]
- Saravanakumar, D.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Detection of enzymatic activity and partial sequence of a chi-tinase gene in Metschnikowia pulcherrima strain MACH1 used as postharvest biocontrol agent. Eur. J. Plant Pathol. 2009, 123, 10. [Google Scholar] [CrossRef]
- Lemos, W.J.; Bovo, B.; Nadai, C.; Crosato, G.; Carlot, M.; Favaron, F.; Giacomini, A.; Corich, V. Biocontrol Ability and Action Mechanism of Starmerella bacillaris (Synonym Candida zemplinina) Isolated from Wine Musts against Gray Mold Disease Agent Botrytis cinerea on Grape and Their Effects on Alcoholic Fermentation. Front. Microbiol. 2016, 7, 1249. [Google Scholar] [CrossRef] [PubMed]
- Janisiewicz, W. Biocontrol of postharvest diseases of temperate fruit. In Plant–Microbe Interactions and Biological Control; Boland, G.J., Kuykendall, L.D., Eds.; Marcel Dekker: New York, NY, USA, 1997; pp. 171–198. [Google Scholar]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Biocontrol of postharvest brown rot of sweet cherries by Saccha-romyces cerevisiae Disva 599, Metschnikowia pulcherrima Disva 267 and Wickerhamomyces anomalus Disva 2 strains. Postharvest Biol. Technol. 2014, 96, 5. [Google Scholar] [CrossRef]
- Ponsone, M.L.; Chiotta, M.L.; Combina, M.; Dalcero, A.; Chulze, S. Biocontrol as a strategy to reduce the impact of ochratoxin A and Aspergillus section Nigri in grapes. Int. J. Food Microbiol. 2011, 151, 70–77. [Google Scholar] [CrossRef]
- Nally, M.C.; Ponsone, M.L.; Pesce, V.M.; Toro, M.E.; Vazquez, F.; Chulze, S. Evaluation of behaviour of Lachancea thermo-tolerans biocontrol agents on grape fermentations. Lett. Appl. Microbiol. 2018, 67, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Bleve, G.; Grieco, F.; Cozzi, G.; Logrieco, A.; Visconti, A. Isolation of epiphytic yeasts with potential for biocontrol of Asper-gillus carbonarius and A. niger on grape. Int. J. Food Microbiol. 2006, 108, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.; Baruzzi, F.; Cocolin, L.; Malfeito-Ferreira, M. Emerging technologies to control Brettanomyces spp. in wine: Recent advances and future trends. Trends Food Sci. Technol. 2020, 99, 88–100. [Google Scholar] [CrossRef]
- Zott, K.; Thibon, C.; Bely, M.; Lonvaud-Funel, A.; Dubourdieu, D.; Masneuf-Pomarede, I. The grape must non-Saccharomyces microbial community: Impact on volatile thiol release. Int. J. Food Microbiol. 2011, 151, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Loira, I.; Escott, C.; del Fresno, J.M.l.; Bañuelos, M.A.; Suárez-Lepe, J.A. Applications of Metschnikowia pul-cherrima in Wine Biotechnology. Fermentation 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Escott, C.; del Fresno, J.M.; Loira, I.; Morata, A.; Tesfaye, W.; González, M.d.; Suarez-Lepe, J.A. Formation of polymeric pigments in red wines through sequential fermentationof flavanol-enriched musts with non-Saccharomyces yeasts. Food Chem. 2018, 239, 975–983. [Google Scholar] [CrossRef]
- Alimardani-Theuil, P.; Gainvors-Claisse, A.; Duchiron, F. Yeasts: An attractive source of pectinases—From gene expression to potential applications: A review. Process. Biochem. 2011, 46, 1525–1537. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Yeast Strain | Extracellular Lytic Enzymes Activity | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Chitinase | β-1,3-glucanase | Pectinase | Protease | Lipase | ||||||
| 25 °C | 0 °C | 25 °C | 0 °C | 25 °C | 0 °C | 25 °C | 0 °C | 25 °C | 0 °C | |
| M. pulcherrima N20/066 | − | − | − | − | + | + | + | + | + | + |
| M. pulcherrima Ale4 | − | − | + | + | + | + | − | − | + | + |
| M. pulcherrima Pr7 | − | − | − | − | − | − | − | − | + | + |
| L. thermotolerans N10 | − | − | − | − | + | + | − | − | + | + |
| H. uvarum Ale5 | − | − | + | + | + | + | + | + | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marsico, A.D.; Velenosi, M.; Perniola, R.; Bergamini, C.; Sinonin, S.; David-Vaizant, V.; Maggiolini, F.A.M.; Hervè, A.; Cardone, M.F.; Ventura, M. Native Vineyard Non-Saccharomyces Yeasts Used for Biological Control of Botrytis cinerea in Stored Table Grape. Microorganisms 2021, 9, 457. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020457
Marsico AD, Velenosi M, Perniola R, Bergamini C, Sinonin S, David-Vaizant V, Maggiolini FAM, Hervè A, Cardone MF, Ventura M. Native Vineyard Non-Saccharomyces Yeasts Used for Biological Control of Botrytis cinerea in Stored Table Grape. Microorganisms. 2021; 9(2):457. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020457
Chicago/Turabian StyleMarsico, Antonio Domenico, Matteo Velenosi, Rocco Perniola, Carlo Bergamini, Scott Sinonin, Vanessa David-Vaizant, Flavia Angela Maria Maggiolini, Alexandre Hervè, Maria Francesca Cardone, and Mario Ventura. 2021. "Native Vineyard Non-Saccharomyces Yeasts Used for Biological Control of Botrytis cinerea in Stored Table Grape" Microorganisms 9, no. 2: 457. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020457