
The Autotrophic Core: An Ancient Network of 404 Reactions Converts H2, CO2, and NH3 into Amino Acids, Bases, and Cofactors

 , , ,
, , ,
Abstract
:
1. Introduction
1.1. Metabolism vs. Genetics?
1.2. Autotrophic Origins and Energy First Link C1 Metabolism to Vents
2. Methods
2.1. Reaction Data Collection
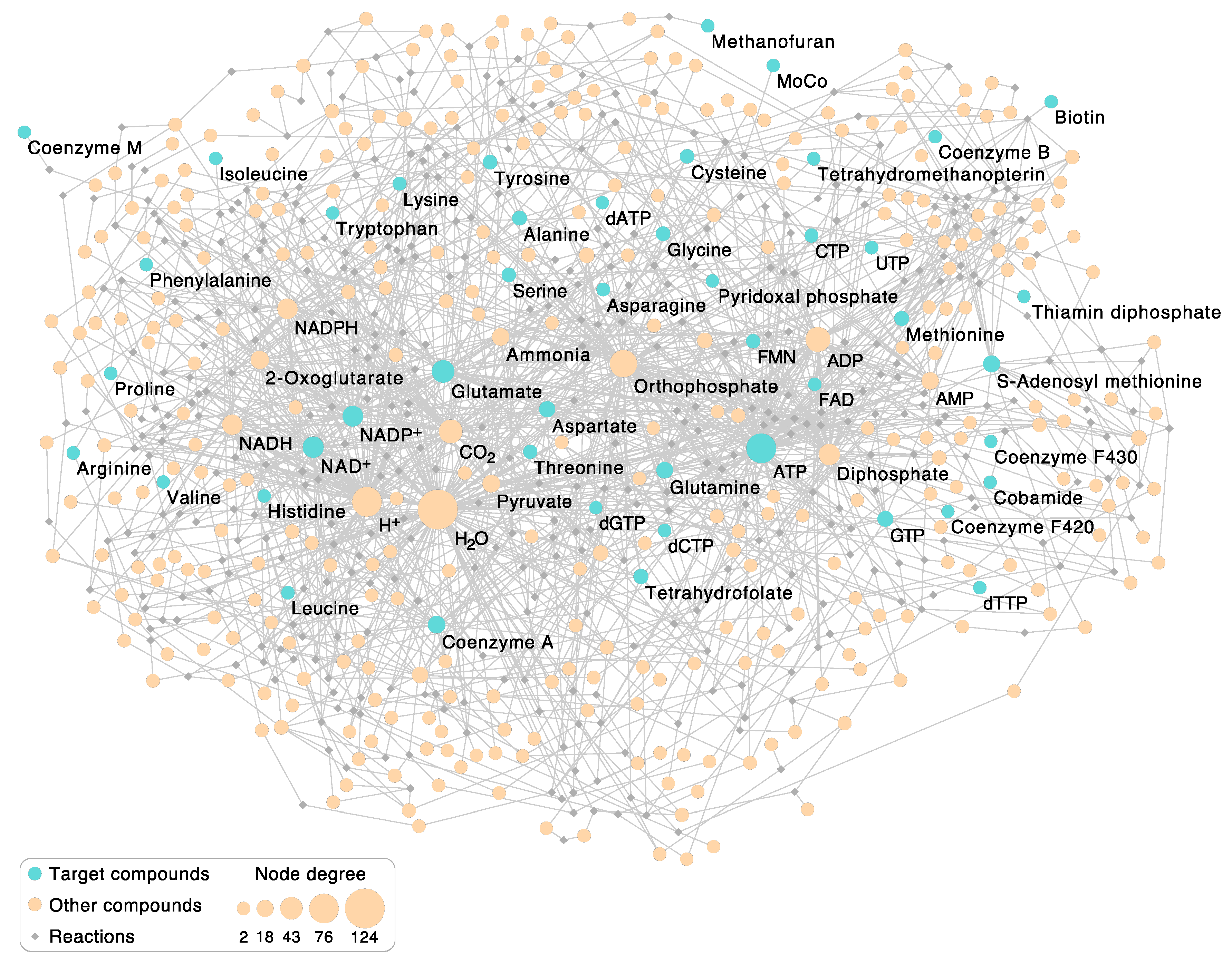
2.2. Visualization of the Autotrophic Core Network
2.3. Different Core Reaction Sets Based on Distinct Identification Approaches
2.4. Statistical Analysis
3. Results
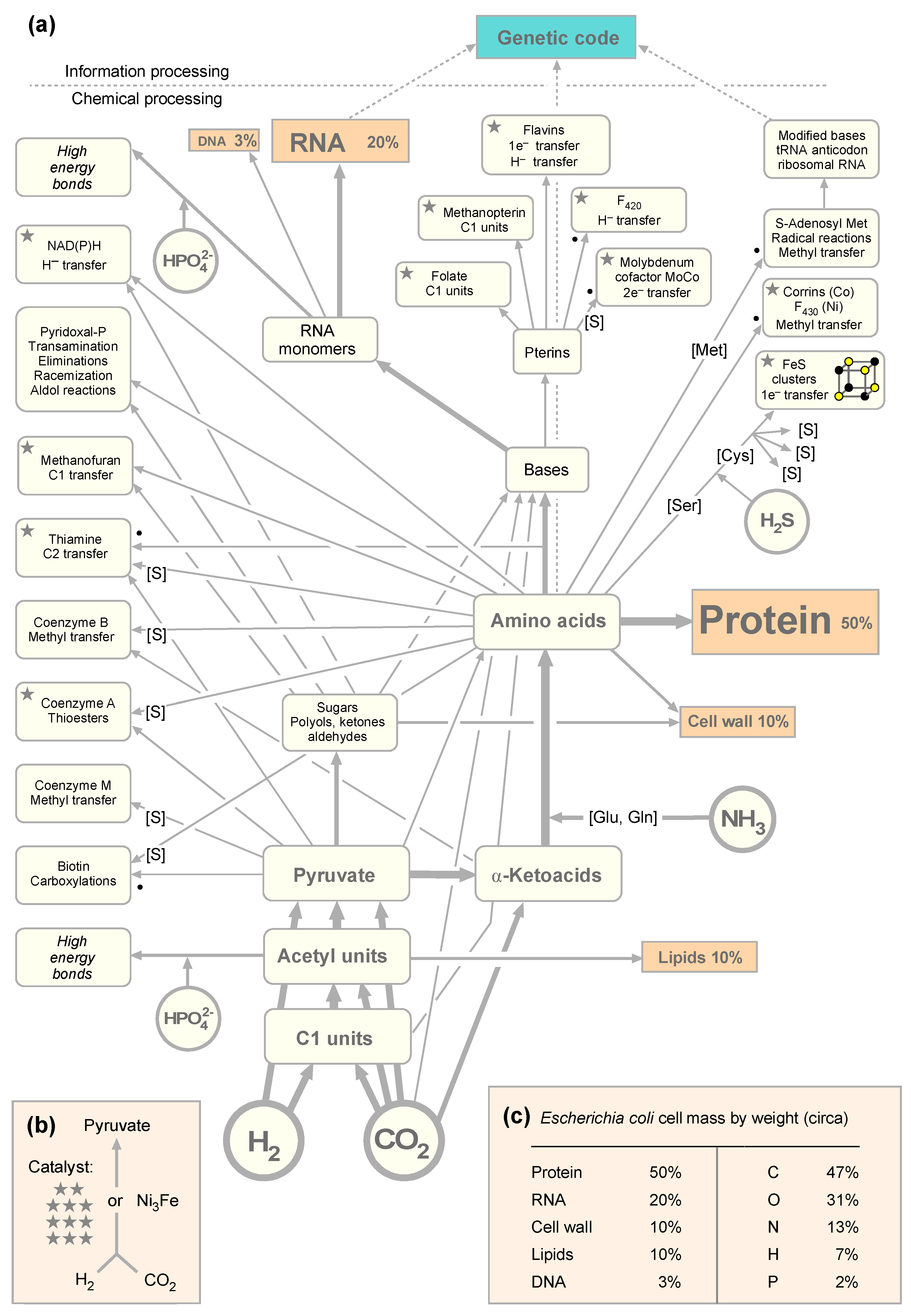
3.1. The Autotrophic Core of Biosynthesis Requires 19 Cofactors
3.2. Enzymatic Reactions in the Autotrophic Core
3.3. Comparison of the Autotrophic Core with LUCA’s Genes and Ancient Autocatalytic Sets
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haeckel, E. Natürliche Schöpfungs-Geschichte. Gemeinverständliche Wissenschaftliche Vorträge über die Entwickelungslehre Zehnte verbesserte Auflage. Zweiter Theil: Allgemeine Stammesgeschichte; Georg Reimer Verlag: Berlin, Germany, 1902. [Google Scholar]
- Winogradsky, S. Beiträge zur Morphologie und Physiologie der Bakterien. H. 1. Zur Morphologie und Physiologie der Schwefelbakterien; A. Felix: Leipzig, Germany, 1888. [Google Scholar]
- Madigan, M.T.; Martinko, J.M.; Bender, K.S.; Buckley, D.H.; Stahl, D.A. Brock Biology of Microorganisms, 15th ed.; Pearson Education: Harlow, UK, 2019. [Google Scholar]
- Decker, K.; Jungermann, K.; Thauer, R.K. Energy production in anaerobic organisms. Angew. Chem. Int. Ed. 1970, 9, 138–158. [Google Scholar] [CrossRef]
- Baross, J.A.; Hoffmann, S.E. Submarine hydrothermal vents and associated gradient environments as sites for the origin and evolution of life. Orig. Life Evol. Biosph. 1985, 15, 327–345. [Google Scholar] [CrossRef]
- Fuchs, G. Alternative pathways of carbon dioxide fixation: Insights into the early evolution of life? Annu. Rev. Microbiol. 2011, 65, 631–658. [Google Scholar] [CrossRef]
- Kowallik, K.V.; Martin, W.F. The origin of symbiogenesis: An annotated English translation of Mereschkowsky’s 1910 paper on the theory of two plasma lineages. Biosystems 2021, 199, 104281. [Google Scholar] [CrossRef]
- Miller, S.L. A production of amino acids under possible primitive earth conditions. Science 1953, 117, 528–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oró, J.; Kimball, A.P. Synthesis of purines under possible primitive earth conditions. I. Adenine from hydrogen cyanide. Arch. Biochem. Biophys. 1961, 94, 217–227. [Google Scholar] [CrossRef]
- Haruna, I.; Spiegelman, S. Specific template requirements of RNA replicases. Proc. Natl. Acad. Sci. USA 1965, 54, 579–587. [Google Scholar] [CrossRef] [Green Version]
- Cech, T.R. The chemistry of self-splicing enzymes. Science 1986, 236, 1532–1539. [Google Scholar] [CrossRef]
- Gilbert, W. The RNA world. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Lincoln, T.A.; Joyce, G.F. Self-sustained replication of an RNA enzyme. Science 2009, 323, 1229–1232. [Google Scholar] [CrossRef] [Green Version]
- Powner, M.W.; Gerland, B.; Sutherland, J.D. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature 2009, 459, 239–242. [Google Scholar] [CrossRef]
- Sutherland, J.D. The origin of life—Out of the blue. Angew. Chem. Int. Ed. 2016, 55, 104–121. [Google Scholar] [CrossRef]
- Shapiro, R. Small molecule interactions were central to the origin of life. Q. Rev. Biol. 2006, 81, 105–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wächtershäuser, G. Before enzymes and templates: Theory of surface metabolism. Microbiol. Rev. 1988, 52, 452–484. [Google Scholar] [CrossRef]
- Orgel, L.E. The implausibility of metabolic cycles on the prebiotic earth. PLoS Biol. 2008, 6, e18–e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamminga, H. Historical perspective: The problem of the origin of life in the context of developments in biology. Orig. Life Evol. Biosph. 1988, 18, 1–11. [Google Scholar] [CrossRef]
- Pascal, R.; Pross, A. Stability and its manifestation in the chemical and biological worlds. Chem. Commun. 2015, 51, 16160–16165. [Google Scholar] [CrossRef] [Green Version]
- Preiner, M.; Asche, S.; Becker, S.; Betts, H.C.; Boniface, A.; Camprubi, E.; Chandru, K.; Erastova, V.; Garg, S.G.; Khawaja, N.; et al. The future of origin of life research: Bridging decades-old divisions. Life 2020, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipmann, F. Projecting backward from the present stage of evolution of biosynthesis. In The Origin of Prebiological Systems and of Their Molecular Matrices; Fox, S.W., Ed.; Academic Press: New York, NY, USA, 1965; pp. 259–280. [Google Scholar]
- Ferry, J.G.; House, C.H. The stepwise evolution of early life driven by energy conservation. Mol. Biol. Evol. 2006, 23, 1286–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corliss, J.B.; Dymond, J.; Gor, L.I.; Edmond, J.M.; Von Herzen, R.P.; Bal, R.D.; Green, K.; Williams, D.; Bainbri, A.; Crane, K.; et al. Submarine thermal springs on the Galapágos rift. Science 1979, 203, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.S.; Karson, J.A.; Blackman, D.K.; Früh-Green, G.L.; Butterfield, D.A.; Lilley, M.D.; Olson, E.J.; Schrenk, M.O.; Roe, K.K.; Lebon, G.T.; et al. An off-axis hydrothermal vent field near the Mid-Atlantic Ridge at 30° N. Nature 2001, 412, 145–149. [Google Scholar] [CrossRef]
- Charlou, J.L.; Donval, J.P.; Fouquet, Y.; Jean-Baptiste, P.; Holm, N. Geochemistry of high H2 and CH4 vent fluids issuing from ultramafic rocks at the Rainbow hydrothermal field (36°14′ N, MAR). Chem. Geol. 2002, 191, 345–359. [Google Scholar] [CrossRef]
- Thauer, R.K.; Kaster, A.-K.; Seedorf, H.; Buckel, W.; Hedderich, R. Methanogenic archaea: Ecologically relevant differences in energy conservation. Nat. Rev. Microbiol. 2008, 6, 579–591. [Google Scholar] [CrossRef]
- Sleep, N.H.; Bird, D.K.; Pope, E.C. Serpentinite and the dawn of life. Philos. Trans. R. Soc. B 2011, 366, 2857–2869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, 6–805. [Google Scholar] [CrossRef] [PubMed]
- Preiner, M.; Igarashi, K.; Muchowska, K.B.; Yu, M.; Varma, S.J.; Kleinermanns, K.; Nobu, M.K.; Kamagata, Y.; Tüysüz, H.; Moran, J.; et al. A hydrogen dependent geochemical analogue of primordial carbon and energy metabolism. Nat. Ecol. Evol. 2020, 4, 534–542. [Google Scholar] [CrossRef]
- Schönheit, P.; Buckel, W.; Martin, W.F. On the origin of heterotrophy. Trends Microbiol. 2016, 24, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Harold, F.M. The Vital Force: A Study of Bioenergetics; WH Freeman: New York, NY, USA, 1986. [Google Scholar]
- Russell, J.B.; Cook, G.M. Energetics of bacterial growth: Balance of anabolic and catabolic reactions. Microbiol. Rev. 1995, 59, 48–62. [Google Scholar] [CrossRef]
- Russell, J.B. The energy spilling reactions of bacteria and other organisms. J. Mol. Microbiol. Biotechnol. 2007, 13, 1–11. [Google Scholar] [CrossRef]
- Van Dijken, J.P.; Scheffers, W.A. Redox balances in the metabolism of sugars by yeasts. FEMS Microbiol. Rev. 1986, 32, 199–224. [Google Scholar] [CrossRef] [Green Version]
- Martin, W.F.; Russell, M.J. On the origin of biochemistry at an alkaline hydrothermal vent. Philos. Trans. R. Soc. B 2007, 362, 1887–1925. [Google Scholar] [CrossRef] [PubMed]
- Muchowska, K.B.; Varma, S.J.; Moran, J. Synthesis and breakdown of universal metabolic precursors promoted by iron. Nature 2019, 569, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Ralser, M. An appeal to magic? The discovery of a non-enzymatic metabolism and its role in the origins of life. Biochem. J. 2018, 475, 2577–2592. [Google Scholar] [CrossRef] [Green Version]
- Thauer, R.K.; Jungermann, K.; Decker, K. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 1977, 41, 100–180. [Google Scholar] [CrossRef] [PubMed]
- Stouthamer, A.H. Energy-yielding pathways. In The Bacteria; Gunsalus, I.C., Ed.; Academic Press: New York, NY, USA, 1978; Volume 6, pp. 389–462. [Google Scholar]
- Hoehler, T.M.; Jørgensen, B.B. Microbial life under extreme energy limitation. Nat. Rev. Microbiol. 2013, 11, 83–94. [Google Scholar] [CrossRef]
- Arndt, N.T.; Nisbet, E.G. Processes on the young earth and the habitats of early life. Annu. Rev. Earth Planet. Sci. 2012, 40, 521–549. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.J.; Hall, A.J.; Martin, W. Serpentinization as a source of energy at the origin of life. Geobiology 2010, 8, 355–371. [Google Scholar] [CrossRef]
- Preiner, M.; Xavier, J.C.; Sousa, F.; Zimorski, V.; Neubeck, A.; Lang, S.Q.; Greenwell, H.C.; Kleinermanns, K.; Tüysüz, H.; McCollom, T.M.; et al. Serpentinization: Connecting geochemistry, ancient metabolism and industrial hydrogenation. Life 2018, 8, 41. [Google Scholar] [CrossRef] [Green Version]
- Proskurowski, G.; Lilley, M.D.; Seewald, J.S.; Früh-Green, G.L.; Olson, E.J.; Lupton, J.E.; Sylva, S.P.; Kelley, D.S. Abiogenic hydrocarbon production at lost city hydrothermal field. Science 2008, 319, 604–607. [Google Scholar] [CrossRef] [Green Version]
- Lang, S.Q.; Butterfield, D.A.; Schulte, M.; Kelley, D.S.; Lilley, M.D. Elevated concentrations of formate, acetate and dissolved organic carbon found at the Lost City hydrothermal field. Geochim. Cosmochim. Acta 2010, 74, 941–952. [Google Scholar] [CrossRef]
- Konn, C.; Charlou, J.L.; Holm, N.G.; Mousis, O. The production of methane, hydrogen and organic compounds in ultramafic-hosted hydrothermal vents of the Mid-Atlantic Ridge. Astrobiology 2015, 15, 381–399. [Google Scholar] [CrossRef] [PubMed]
- Schrenk, M.O.; Brazelton, W.J.; Lang, S.Q. Serpentinization, carbon, and deep life. Rev. Mineral. Geochem. 2013, 75, 575–606. [Google Scholar] [CrossRef] [Green Version]
- Varma, S.J.; Mochowska, K.B.; Chatelain, P.; Moran, J. Native iron reduces CO2 to intermediates and endproducts of the acetyl-CoA pathway. Nat. Ecol. Evol. 2018, 2, 1019–1024. [Google Scholar] [CrossRef]
- Weiss, M.C.; Sousa, F.L.; Mrnjavac, N.; Neukirchen, S.; Roettger, M.; Nelson-Sathi, S.; Martin, W.F. The physiology and habitat of the last universal common ancestor. Nat. Microbiol. 2016, 1, 16116. [Google Scholar] [CrossRef] [PubMed]
- Xavier, J.C.; Hordijk, W.; Kauffman, S.; Steel, M.; Martin, W.F. Autocatalytic chemical networks preceded proteins and RNA in evolution. Proc. R. Soc. Lond. B 2020, 287. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.Q. KEGG pathway database. In Encyclopedia of Systems Biology; Dubitzky, W., Wolkenhauer, O., Cho, K.H., Yokota, H., Eds.; Springer: New York, NY, USA, 2013; pp. 1068–1069. [Google Scholar]
- Hazra, A.B.; Han, A.W.; Mehta, A.P.; Mok, K.C.; Osadchiy, V.; Begley, T.P.; Taga, M.E. Anaerobic biosynthesis of the lower ligand of vitamin B12. Proc. Natl. Acad. Sci. USA 2015, 112, 10792–10797. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Berg, I.A.; Kockelkorn, D.; Ramos-Vera, W.H.; Say, R.F.; Zarzycki, J.; Hügler, M.; Alber, B.E.; Fuchs, G. Autotrophic carbon fixation in archaea. Nat. Rev. Microbiol. 2010, 8, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Hügler, M.; Sievert, S.M. Beyond the Calvin cycle: Autotrophic carbon fixation in the ocean. Ann. Rev. Mar. Sci. 2011, 3, 261–289. [Google Scholar] [CrossRef] [Green Version]
- Martin, W.F. Older than genes: The acetyl CoA pathway and origins. Front. Microbiol. 2020, 11, 11–817. [Google Scholar] [CrossRef]
- Mall, A.; Sobotta, J.; Huber, C.; Tschirner, C.; Kowarschik, S.; Bačnik, K.; Mergelsberg, M.; Boll, M.; Hügler, M.; Eisenreich, W.; et al. Reversibility of citrate synthase allows autotrophic growth of a thermophilic bacterium. Science 2018, 359, 563–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunoura, T.; Chikaraishi, Y.; Izaki, R.; Suwa, T.; Sato, T.; Harada, T.; Mori, K.; Kato, Y.; Miyazaki, M.; Shimamura, S.; et al. A primordial and reversible TCA cycle in a facultatively chemolithoautotrophic thermophile. Science 2018, 359, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Maden, B.E. Tetrahydrofolate and tetrahydromethanopterin compared: Functionally distinct carriers in C1 metabolism. Biochem. J. 2000, 350, 609–629. [Google Scholar] [CrossRef] [PubMed]
- Sousa, F.L.; Martin, W.F. Biochemical fossils of the ancient transition from geoenergetics to bioenergetics in prokaryotic one carbon compound metabolism. Biochim. Biophys. Acta 2014, 1837, 964–981. [Google Scholar] [CrossRef] [Green Version]
- Davidson, V.L. Protein-derived cofactors revisited: Empowering amino acid residues with new functions. Biochemistry 2018, 57, 3115–3125. [Google Scholar] [CrossRef]
- Canavelli, P.; Islam, S.; Powner, M.W. Peptide ligation by chemoselective aminonitrile coupling in water. Nature 2019, 571, 546–549. [Google Scholar] [CrossRef] [PubMed]
- Xavier, J.C.; Preiner, M.; Martin, W.F. Something special about CO-dependent CO2 fixation. FEBS J. 2018, 285, 4181–4195. [Google Scholar] [CrossRef] [Green Version]
- Knowles, C.J. Cyanide utilization and degradation by microorganisms. Ciba. Found. Symp. 1988, 140, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, R.F.; Dolghih, E.; Kunz, D.A. Enzymatic assimilation of cyanide via pterin-dependent oxygenolytic cleavage to ammonia and formate in Pseudomonas fluorescens NCIMB 11764. Appl. Environ. Microbiol. 2004, 70, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Huber, C.; Wächtershäuser, G. α-hydroxy and α-amino acids under possible hadean, volcanic origin-of-life conditions. Science 2006, 314, 630–632. [Google Scholar] [CrossRef]
- Liu, Y.; Beer, L.L.; Whitman, W.B. Sulfur metabolism in archaea reveals novel processes. Environ. Microbiol. 2012, 14, 2632–2644. [Google Scholar] [CrossRef]
- Marshall, M. How the first life on Earth survived its biggest threat—Water. Nature 2020, 588, 210–213. [Google Scholar] [CrossRef]
- Do Nascimento Vieira, A.; Kleinermanns, K.; Martin, W.F.; Preiner, M. The ambivalent role of water at the origins of life. FEBS Lett. 2020, 594, 2717–2733. [Google Scholar] [CrossRef] [PubMed]
- Lamadrid, H.M.; Rimstidt, J.D.; Schwarzenbach, E.M.; Klein, F.; Ulrich, S.; Dolocan, A.; Bodnar, R.J. Effect of water activity on rates of serpentinization of olivine. Nat. Commun. 2017, 8, 16107. [Google Scholar] [CrossRef] [PubMed]
- Berg, I.A.; Kockelkorn, D.; Buckel, W.; Fuchs, G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in archaea. Science 2007, 318, 1782–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raushel, F.M.; Thoden, J.B.; Holden, H.M. The amidotransferase family of enzymes: Molecular machines for the production and delivery of ammonia. Biochemistry 1999, 38, 7891–7899. [Google Scholar] [CrossRef]
- Goldford, J.E.; Hartman, H.; Smith, T.F.; Segrè, D. Remnants of an ancient metabolism without phosphate. Cell 2017, 168, 1126–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenov, S.N.; Kraft, L.J.; Ainla, A.; Zhao, M.; Baghbanzadeh, M.; Campbell, V.E.; Kang, K.; Fox, J.M.; Whitesides, G.M. Autocatalytic, bistable, oscillatory networks of biologically relevant organic reactions. Nature 2016, 537, 656–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muchowska, K.B.; Varma, S.J.; Moran, J. Nonenzymatic metabolic reactions and life’s origins. Chem. Rev. 2020, 120, 7708–7744. [Google Scholar] [CrossRef]
- Preiner, M.; Xavier, J.C.; Vieira, A.D.N.; Kleinermanns, K.; Allen, J.F.; Martin, W.F. Catalysts, autocatalysis and the origin of metabolism. Interface Focus 2019, 9, 9–20190072. [Google Scholar] [CrossRef]
- Morowitz, H.J.; Kostelnik, J.D.; Yang, J.; Cody, G.D. The origin of intermediary metabolism. Proc. Natl. Acad. Sci. USA 2000, 97, 7704–7708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfenden, R. Benchmark reaction rates, the stability of biological molecules in water, and the evolution of catalytic power in enzymes. Annu. Rev. Biochem. 2011, 80, 645–667. [Google Scholar] [CrossRef]
- Muchowska, K.B.; Varma, S.J.; Chevallot-Beroux, E.; Lethuillier-Karl, L.; Li, G.; Moran, J. Metals promote sequences of the reverse Krebs cycle. Nat. Ecol. Evol. 2017, 1, 1716–1721. [Google Scholar] [CrossRef]
- Messner, C.B.; Driscoll, P.C.; Piedrafita, G.; De Volder, M.F.L.; Ralser, M. Nonenzymatic gluconeogenesis-like formation of fructose 1,6-bisphosphate in ice. Proc. Natl. Acad. Sci. USA 2017, 114, 7403–7407. [Google Scholar] [CrossRef] [Green Version]
- Sousa, F.L.; Nelson-Sathi, S.; Martin, W.F. One step beyond a ribosome: The ancient anaerobic core. Biochim. Biophys. Acta. 2016, 1857, 1027–1038. [Google Scholar] [CrossRef] [Green Version]
- Krzycki, J.A. The direct genetic encoding of pyrrolysine. Curr. Opin. Microbiol. 2005, 8, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Böck, A.; Forchhammer, K.; Heider, J.; Leinfelder, W.; Sawers, G.; Veprek, B.; Zinoni, F. Selenocysteine: The 21st amino acid. Mol. Microbiol. 1991, 5, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Grosjean, H.; de Crécy-Lagard, V.; Marck, C. Deciphering synonymous codons in the three domains of life: Co-evolution with specific tRNA modification enzymes. FEBS Lett. 2010, 584, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Xavier, J.C.; Patil, K.R.; Rocha, I. Integration of biomass formulations of genome-scale metabolic models with experimental data reveals universally essential cofactors in prokaryotes. Metab. Eng. 2017, 39, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Compound | Frequency |
|---|---|
| H2O | 125 |
| ATP | 77 |
| H+ | 76 |
| Pi | 66 |
| ADP | 55 |
| CO2 | 49 |
| Glutamate | 44 |
| PPi | 37 |
| NAD+ | 37 |
| NADP+ | 35 |
| NADPH | 34 |
| NADH | 33 |
| 2-Oxoglutarate | 24 |
| Pyruvate | 22 |
| NH3 | 21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wimmer, J.L.E.; Vieira, A.d.N.; Xavier, J.C.; Kleinermanns, K.; Martin, W.F.; Preiner, M. The Autotrophic Core: An Ancient Network of 404 Reactions Converts H2, CO2, and NH3 into Amino Acids, Bases, and Cofactors. Microorganisms 2021, 9, 458. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020458
Wimmer JLE, Vieira AdN, Xavier JC, Kleinermanns K, Martin WF, Preiner M. The Autotrophic Core: An Ancient Network of 404 Reactions Converts H2, CO2, and NH3 into Amino Acids, Bases, and Cofactors. Microorganisms. 2021; 9(2):458. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020458
Chicago/Turabian StyleWimmer, Jessica L. E., Andrey do Nascimento Vieira, Joana C. Xavier, Karl Kleinermanns, William F. Martin, and Martina Preiner. 2021. "The Autotrophic Core: An Ancient Network of 404 Reactions Converts H2, CO2, and NH3 into Amino Acids, Bases, and Cofactors" Microorganisms 9, no. 2: 458. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020458