
Investigation of the Physiology of the Obligate Alkaliphilic Bacillus marmarensis GMBE 72T Considering Its Alkaline Adaptation Mechanism for Poly(3-hydroxybutyrate) Synthesis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cultivation of B. marmarensis in the Absence of Carbon Sources
2.3. Investigation of the Physiology of B. marmarensis Using K+ and M2+ Ions and Different Carbon Sources
2.3.1. Effect of K+ and Mg2+ Ions on PHB Synthesis
2.3.2. Effect of Lactose and Galactose on PHB Synthesis
2.3.3. Effect of Different Nitrogen Sources and Nitrogen Limitation on PHB Synthesis
2.3.4. Effect of Vegetable Oil as Carbon Source on PHB Synthesis
2.4. Extraction of PHB from B. marmarensis
2.5. Determination of Reducing Sugar Concentration
3. Results
3.1. The Effect of K+ and Mg2+ Ions on PHB Synthesis
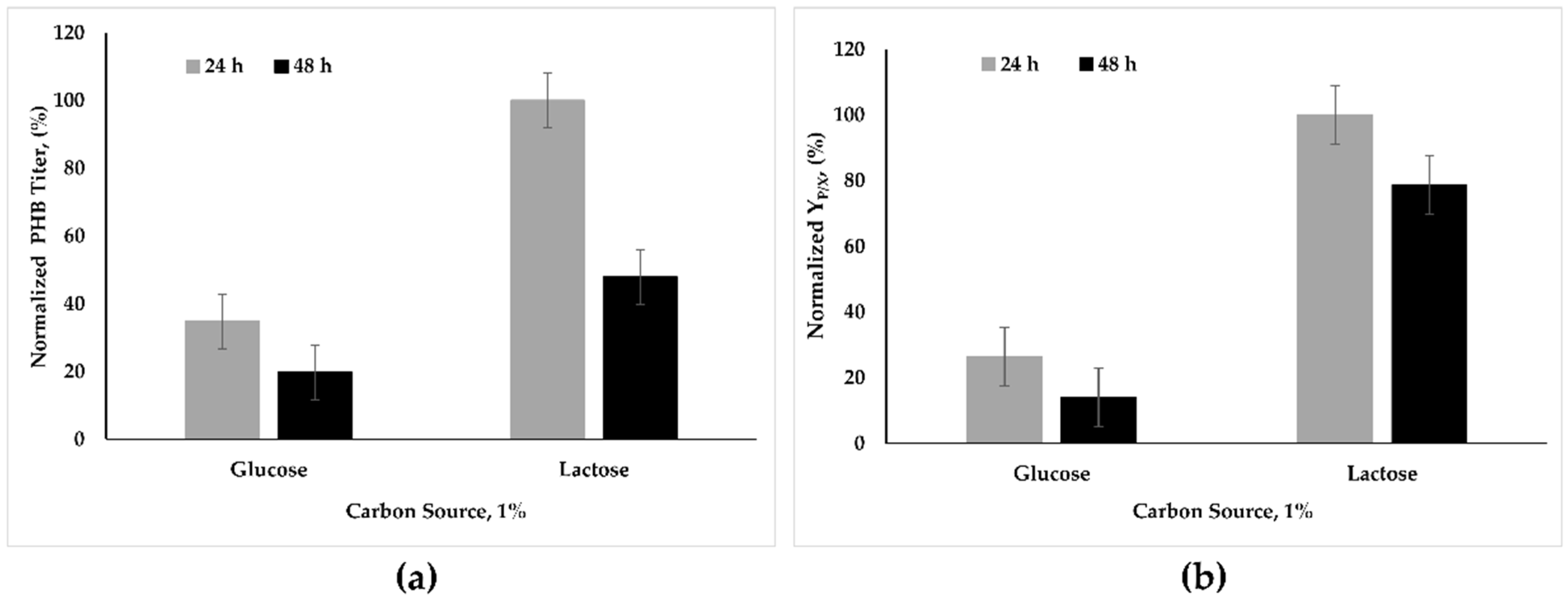
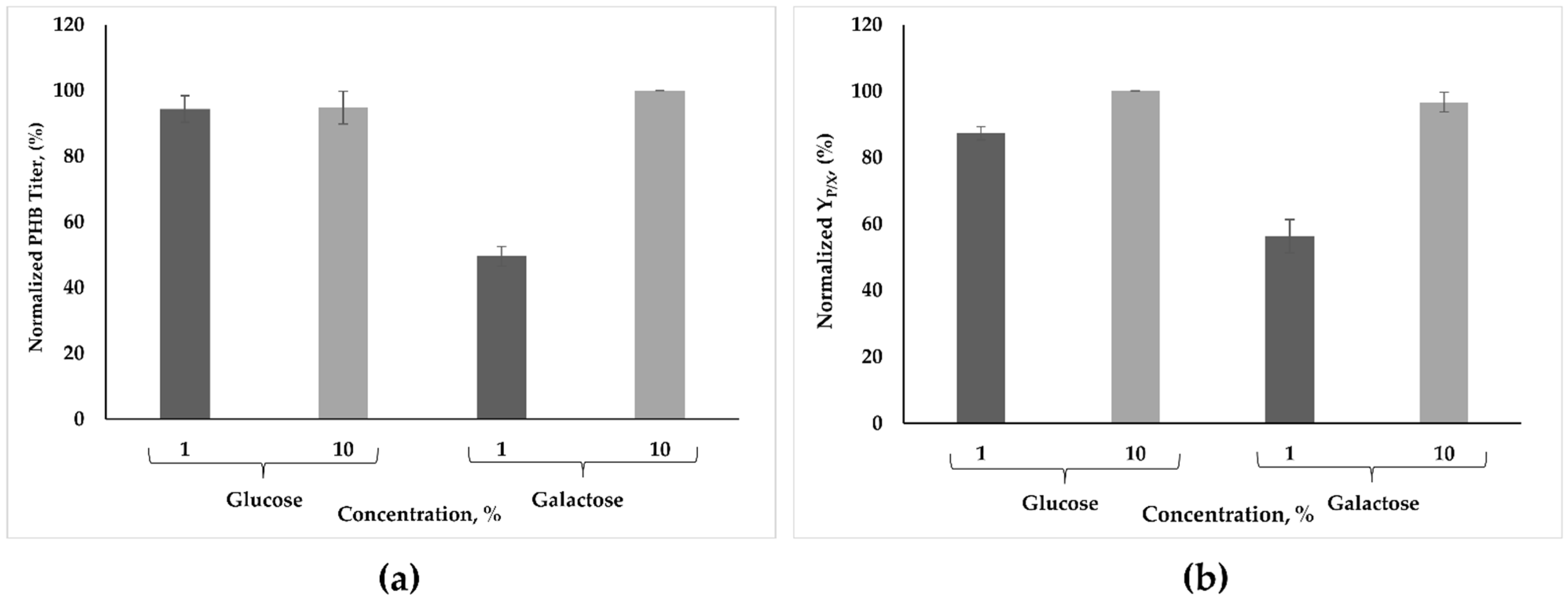
3.2. Effect of Lactose, Galactose, and Abundant Carbon Sources on PHB Synthesis
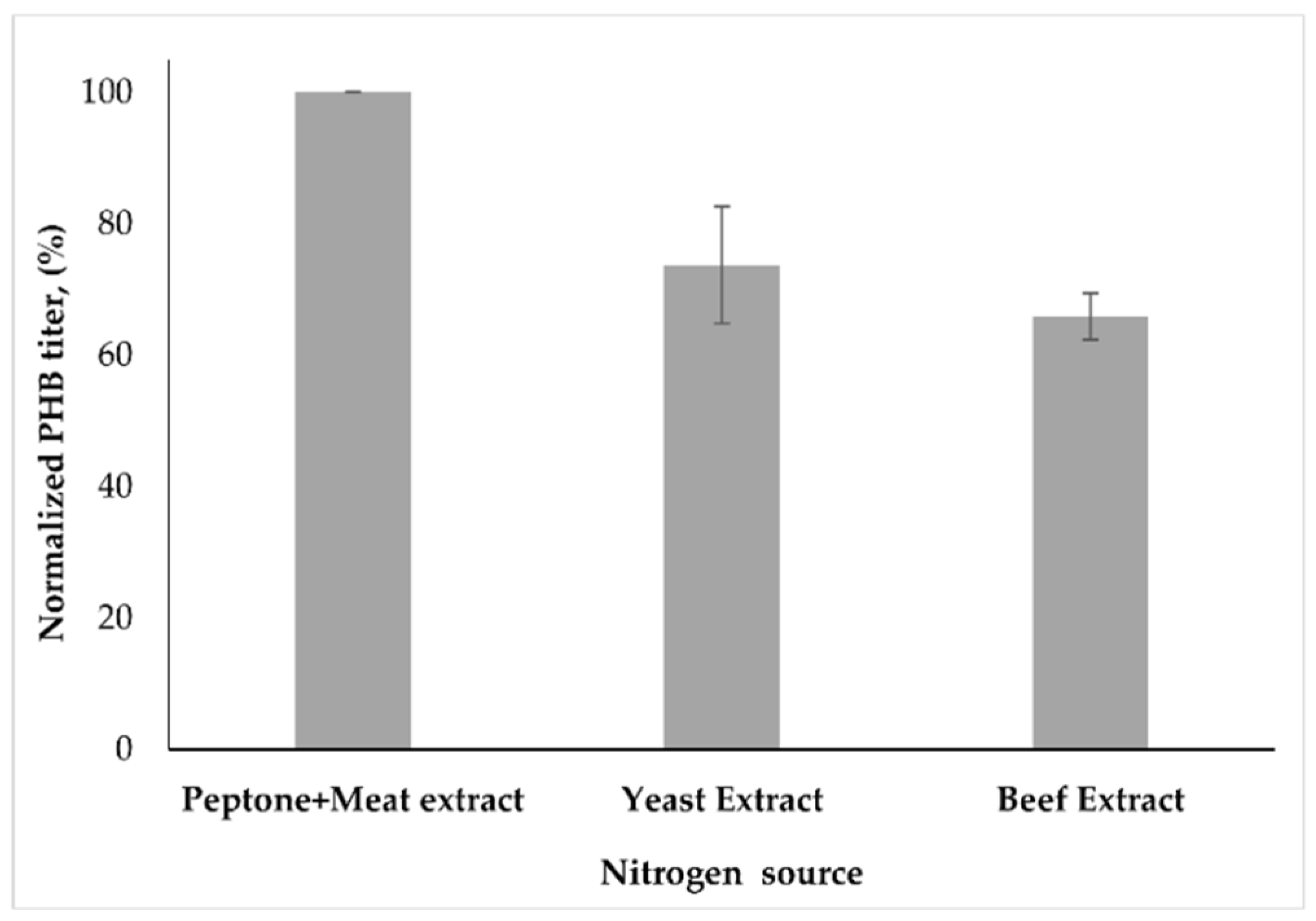
3.3. Effect of Nitrogen Sources and Nitrogen Limitation on PHB Synthesis
3.4. Effect of Vegetable Oil and Peptone Removal on PHB Synthesis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Penkhrue, W.; Jendrossek, D.; Khanongnuch, C.; Pathom-aree, W.; Aizawa, T.; Behrens, R.L.; Lumyong, S. Response surface method for polyhydroxybutyrate (PHB) bioplastic accumulation in Bacillus drentensis BP17 using pineapple peel. PLoS ONE 2020, 15, e0230443. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.Y.; Kim, C.; Moon, J.; Heo, J.; Jung, S.P.; Kim, J.R. Polymer film-based screening and isolation of polylactic acid (PLA)-degrading microorganisms. J. Microbiol. Biotechnol. 2017, 27, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Zikmanis, P.; Kolesovs, S.; Semjonovs, P. Production of biodegradable microbial polymers from whey. Bioresour. Bioprocess. 2020, 7, 1–15. [Google Scholar] [CrossRef]
- Lemoigne, M. Etudes sur L’autolyse microbienne acidification par formation D’acide β-oxybutyrique. Ann. Inst. Pasteur 1925, 39, 144–173. [Google Scholar]
- Nehra, K.; Jaglan, A.; Shaheen, A.; Yadav, J.; Lathwal, P.; Singh, M. Production of Poly-β-Hydroxybutyrate (PHB) by bacteria isolated from rhizospheric soils. Int. J. Microb. Resour. Technol. 2015, 2, 38–48. [Google Scholar]
- Bora, L. Polyhydroxybutyrate Accumulation in Bacillus megaterium and Optimization of Process Parameters Using Response Surface Methodology. J. Polym. Environ. 2013, 21, 415–420. [Google Scholar] [CrossRef]
- Smith, M.K.; Paleri, D.M.; Abdelwahab, M.; Mielewski, D.F.; Misra, M.; Mohanty, A.K. Sustainable composites from poly (3-hydroxybutyrate) (PHB) bioplastic and agave natural fibre. Green Chem. 2020, 22, 3906. [Google Scholar] [CrossRef]
- Chen, G.; Wang, Y. Medical applications of biopolyesters polyhydroxyalkanoates. Chin. J. Polym. Sci. 2013, 31, 719–736. [Google Scholar] [CrossRef]
- Shivam, P. Recent Developments on biodegradable polymers and their future trends. Int. Res. J. Sci. Eng. 2016, 4, 17–26. [Google Scholar]
- Polyhydroxybutyrate (PHB) Market—Global Industry Analysis, Size, Share, Growth, Trends, and Forecast 2018–2026. Available online: https://www.transparencymarketresearch.com/polyhydroxybutyrate-market.html (accessed on 29 December 2020).
- Hassan, M.A.; Bakhiet, E.K.; Ali, S.G.; Hussien, H.R. Production and characterization of polyhydroxybutyrate (PHB) produced by Bacillus sp. isolated from Egypt. J. App. Pharm. Sci. 2016, 6, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Kourmentza, C.; Plácido, J.; Venetsaneas, N.; Burniol-Figols, A.; Varrone, C.; Gavala, H.N.; Reis, M.A. Recent advances and challenges towards sustainable polyhydroxyalkanoate (PHA) production. Bioengineering 2017, 4, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favaro, L.; Basaglia, M.; Casella, S. Improving polyhydroxyalkanoate production from inexpensive carbon sources by genetic approaches: A review. Biofuels Bioprod. Biorefin. 2019, 13, 208–227. [Google Scholar] [CrossRef] [Green Version]
- Denizci, A.A.; Kazan, D.; Erarslan, A. Bacillus marmarensis sp. nov., an alkaliphilic, protease-producing bacterium isolated from mushroom compost. Int. J. Syst. Evol. Microbiol. 2010, 60, 1590–1594. [Google Scholar] [CrossRef] [PubMed]
- Wernick, D.G.; Choi, K.Y.; Tat, C.A.; Rivera, J.G.L.; Liao, J.C. Genome sequence of the extreme obligate alkaliphile Bacillus marmarensis strain DSM 21297. Genome Announc. 2013, 1, e00967-13. [Google Scholar] [CrossRef] [Green Version]
- Wernick, D.G.; Pontrelli, S.P.; Pollock, A.W.; Liao, J.C. Sustainable biorefining in wastewater by engineered extreme alkaliphile Bacillus marmarensis. Sci. Rep. 2016, 6, 20224. [Google Scholar] [CrossRef] [PubMed]
- Özgören, T.; Pinar, O.; Bozdağ, G.; Denizci, A.A.; Gündüz, O.; Hatır, P.Ç.; Kazan, D. Assessment of poly (3-hydroxybutyrate) synthesis from a novel obligate alkaliphilic Bacillus marmarensis and generation of its composite scaffold via electrospinning. Int. J. Biol. Macromol. 2018, 119, 982–991. [Google Scholar] [CrossRef]
- Altinisik Kaya, F.E.; Avci, F.G.; Sayar, N.A.; Kazan, D.; Sayar, A.A.; Sariyar Akbulut, B. What Are the Multi-Omics Mechanisms for Adaptation by Microorganisms to High Alkalinity? A Transcriptomic and Proteomic Study of a Bacillus Strain with Industrial Potential. OMICS 2018, 22, 717–732. [Google Scholar] [CrossRef]
- Berwig, K.H.; Baldasso, C.; Dettmer, A. Production and characterization of poly (3-hydroxybutyrate) generated by Alcaligenes latus using lactose and whey after acid protein precipitation process. Bioresour. Technol. 2016, 218, 31–37. [Google Scholar] [CrossRef]
- Peña, C.; Castillo, T.; García, A.; Millán, M.; Segura, D. Biotechnological strategies to improve production of microbial poly-(3-hydroxybutyrate): A review of recent research work. Microb. Biotechnol. 2014, 7, 278–293. [Google Scholar] [CrossRef]
- Zafar, M.; Kumar, S.; Kumar, S.; Dhiman, A.K. Artificial intelligence based modeling and optimization of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) production process by using Azohydromonas lata MTCC 2311 from cane molasses supplemented with volatile fatty acids: A genetic algorithm paradigm. Bioresour. Technol. 2012, 104, 631–641. [Google Scholar] [CrossRef]
- Mozumder, M.S.I.; De Wever, H.; Volcke, E.I.; Garcia-Gonzalez, L. A robust fed-batch feeding strategy independent of the carbon source for optimal polyhydroxybutyrate production. Process. Biochem. 2014, 49, 365–373. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sindhu, R.; Ammu, B.; Binod, P.; Deepthi, S.K.; Ramachandran, K.B.; Soccol, C.R.; Pandey, A. Production and characterization of poly-3-hydroxybutyrate from crude glycerol by Bacillus sphaericus NII 0838 and improving its thermal properties by blending with other polymers. Braz. Arch. Biol. Technol. 2011, 54, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Sun, W.; Wang, H.; Geng, A. Polyhydroxybutyrate production from oil palm empty fruit bunch using Bacillus megaterium R11. Bioresour. Technol. 2013, 147, 307–314. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Chai, Y.; Beauregard, P.B.; Vlamakis, H.; Losick, R.; Kolter, R. Galactose Metabolism Plays a Crucial Role in Biofilm Formation by Bacillus subtilis. mBio 2012, 3, e00184-12. [Google Scholar] [CrossRef] [Green Version]
- Eggers, J.; Steinbüchel, A. Poly(3-Hydroxybutyrate) Degradation in Ralstonia eutropha H16 Is Mediated Stereoselectively to (S)-3-Hydroxybutyryl Coenzyme A (CoA) via Crotonyl-CoA. J. Bacteriol. 2013, 195, 3213–3223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verlinden, R.A.J.; Hill, D.J.; Kenward, M.A.; Williams, C.D.; Radecka, L. Bacterial synthesis of biodegradable polyhydroxyalkanoates. J. Appl. Microbiol. 2007, 102, 1437–1449. [Google Scholar] [CrossRef] [PubMed]
- Velázquez-Sánchez, C.; Espín, G.; Peña, C.; Segura, D. The Modification of regulatory circuits involved in the control of polyhydroxyalkanoates metabolism to improve their production. Front. Bioeng. Biotechnol. 2020, 8, 386. [Google Scholar] [CrossRef]
- Hassan, K.A.; Fagerlund, A.; Elbourne, L.D.H.; VoÈroÈs, A.; Kroeger, J.K.; Simm, R.; Tourasse, N.J.; Finke, S.; Henderson, P.J.F.; Økstad, O.A.; et al. The Putative Drug Efflux Systems of the Bacillus cereus Group. PLoS ONE 2017, 12, e0176188. [Google Scholar] [CrossRef]
- Stülke, J.; Hillen, W. Regulation of Carbon Catabolism in Bacillus Species. Annu. Rev. Microbiol. 2000, 54, 849–880. [Google Scholar] [CrossRef] [PubMed]
- Thirumala, M.; Reddy, S.V.; Mahmood, S.K. Production and characterization of PHB from two novel strains of Bacillus spp. Isolated from soil and activated sludge. J. Ind. Microbiol. Biotechnol. 2010, 37, 271–278. [Google Scholar] [CrossRef]
- Bachem, S.; Stülke, J. Regulation of the Bacillus subtilis GlcT antiterminator protein by components of the phosphotransferase system. J. Bacteriol. 1998, 180, 5319–5326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verlinden, R.A.; Hill, D.J.; Kenward, M.A.; Williams, D.C.; Piotrowska-Seget, Z.; Radecka, I.K. Production of polyhydroxyalkanoates from waste frying oil by Cupriavidus necator. AMB Express 2011, 1, 11. [Google Scholar] [CrossRef] [Green Version]
- Madison, L.L.; Huisman, G.W. Metabolic engineering of poly (3-hydroxyalkanoates): From DNA to plastic. Microbiol. Mol. Biol. Rev. 1999, 63, 21–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaguchi, A.; Iwamoto-Kihara, A.; Sato, N. Biosynthesis and Degradation of Fatty Acids. Compr. Nat. Prod. Chem. 1999, 23–59. [Google Scholar] [CrossRef]
- Eggink, G.; van der Wal, H.; Hujbert, G.N.M.; Waard, P. Oleic acid as a substrate for poly-3-hydroxyalkanoate formation in Alcaligenes eutrophus and Pseudomonas putida. Ind. Crops Prod. 1992, 1, 157–163. [Google Scholar] [CrossRef]
- Siliakus, M.F.; Oost, J.; Kengen, S.W.M. Adaptations of archaeal and bacterial membranes to variations in temperature, pH and pressure. Extremophiles 2017, 21, 651–670. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C Source (1%) | N Source | PHB Titer (mg/L) | YP/X (mg PHB/g Dry Cell Mass) |
|---|---|---|---|
| Lactose | Yeast extract (0.8%) | 20 ± 4.86 | 30.30 ± 3.636 |
| Meat extract (0.3%) and peptone (0.5%) | 140 ± 32.20 | 125 ± 25.40 | |
| Glucose | Yeast extract (0.8%) | 35 ± 6.65 | 32.71 ± 3.108 |
| Meat extract (0.3%) and peptone (0.5%) | 60 ± 16.60 | 35.29 ± 1.605 | |
| Olive oil | Yeast extract (0.8%) | 10 ± 2.16 | 12.50 ± 2.625 |
| Meat extract (0.3%) and peptone (0.5%) | 30 ± 4.21 | 25 ± 3.762 |
| Enzyme | EC Number | Target Substrate | Product | |
|---|---|---|---|---|
| Lactose to Glucose and Galactose | Beta-galactosidase | 3.2.1.23 | Lactose | D-Glucose D-Galactose |
| Galactose to Glucose | UDP-glucose 4-epimerase | 5.1.3.2 | D-Glucose | D-Galactose |
| Glucose to Acetyl-CoA | Glucokinase (hexokinase) | 2.7.1.2 | D-glucose | D-glucose 6-phosphate |
| Phosphohexose isomerase | 5.3.1.9 | D-glucose 6-phosphate | D-fructose 6-phosphate | |
| 6-phosphofructokinase | 2.7.1.11 | D-fructose 6-phosphate | D-fructose 1,6-bisphosphate | |
| Aldolase | 4.1.2.13 | D-fructose 1,6-bisphosphate | D-glyceraldehyde 3-phosphate Glycerone phosphate | |
| Triosephosphate isomerase | 5.3.1.1 | D-glyceraldehyde 3-phosphate | Glycerone phosphate | |
| Glyceraldehyde-3-phosphate dehydrogenase | 1.2.1.12 | D-glyceraldehyde 3-phosphate | 3-phospho-D-glyceroyl phosphate | |
| Phosphoglycerate kinase | 2.7.2.3 | 3-phospho-D-glyceroyl phosphate | 3-phospho-D-glycerate | |
| Phosphoglycerate mutase | 5.4.2.1 (5.4.2.11) | 3-phospho-D-glycerate | 2-phospho-D-glycerate | |
| Enolase | 4.2.1.11 | 2-phospho-D-glycerate | Phosphoenol pyruvate | |
| Pyruvate kinase | 2.7.1.40 | Phosphoenol pyruvate | Pyruvate | |
| Pyruvate dehydrogenase complex | 1.2.4.1 2.3.1.12 1.8.1.4 | Pyruvate | Acetyl-CoA | |
| Acetyl-CoA to PHB Synthesis | Acetyl-CoA acetyltransferase | 2.3.1.9 | Acetyl-CoA | Acetoacetyl-CoA |
| 3-hydroxybutyryl-CoA dehydrogenase | 1.1.1.157 | Acetoacetyl-CoA | 3-Hydroxybutyryl-CoA | |
| Polyhydroxyalkanoic acid synthase | 2.3.1.- | 3-Hydroxybutyryl-CoA | Polyhydroxybutyrate (PHB) | |
| Pyruvate to Lactate | L-lactate dehydrogenase | 1.1.1.27 | Pyruvate | Lactate |
| Acetyl CoA to TCA cycle and Succinate | Citrate synthase | 2.3.3.1 | Acetyl-CoA + oxaloacetate | Citrate |
| Fatty acid degradation for PHB Synthesis | Long-chain fatty-acid-CoA ligase | 6.2.1.3 | Fatty acids | Fatty-acyl-CoA |
| Acyl-CoA dehydrogenase | 1.3.8.7 | Fatty-acyl-CoA | 2-Enoyl-CoA | |
| Enoyl-CoA hydratase | 4.2.1.17 | 2-Enoyl-CoA | (R)-3-hydroxyacyl-CoA | |
| Polyhydroxyalkanoic acid synthase | 2.3.1.- | (R)-3-hydroxyacyl-CoA | Polyhydroxybutyrate (PHB) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atakav, Y.; Pinar, O.; Kazan, D. Investigation of the Physiology of the Obligate Alkaliphilic Bacillus marmarensis GMBE 72T Considering Its Alkaline Adaptation Mechanism for Poly(3-hydroxybutyrate) Synthesis. Microorganisms 2021, 9, 462. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020462
Atakav Y, Pinar O, Kazan D. Investigation of the Physiology of the Obligate Alkaliphilic Bacillus marmarensis GMBE 72T Considering Its Alkaline Adaptation Mechanism for Poly(3-hydroxybutyrate) Synthesis. Microorganisms. 2021; 9(2):462. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020462
Chicago/Turabian StyleAtakav, Yağmur, Orkun Pinar, and Dilek Kazan. 2021. "Investigation of the Physiology of the Obligate Alkaliphilic Bacillus marmarensis GMBE 72T Considering Its Alkaline Adaptation Mechanism for Poly(3-hydroxybutyrate) Synthesis" Microorganisms 9, no. 2: 462. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020462