
The Environmental Spread of Aspergillus terreus in Tyrol, Austria


 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Environmental Samples
2.2. Clinical A. terreus
2.3. Species Identification of Environmental and Clinical A. terreus
2.4. Meteorological and Geographical Data
2.5. Antifungal Susceptibility Testing
2.6. Microsatellite Typing of A. terreus
2.7. Statistical Analysis
3. Results
3.1. A. Terreus Collected from Environmental Samples
3.2. Environmental Sampling Conditions
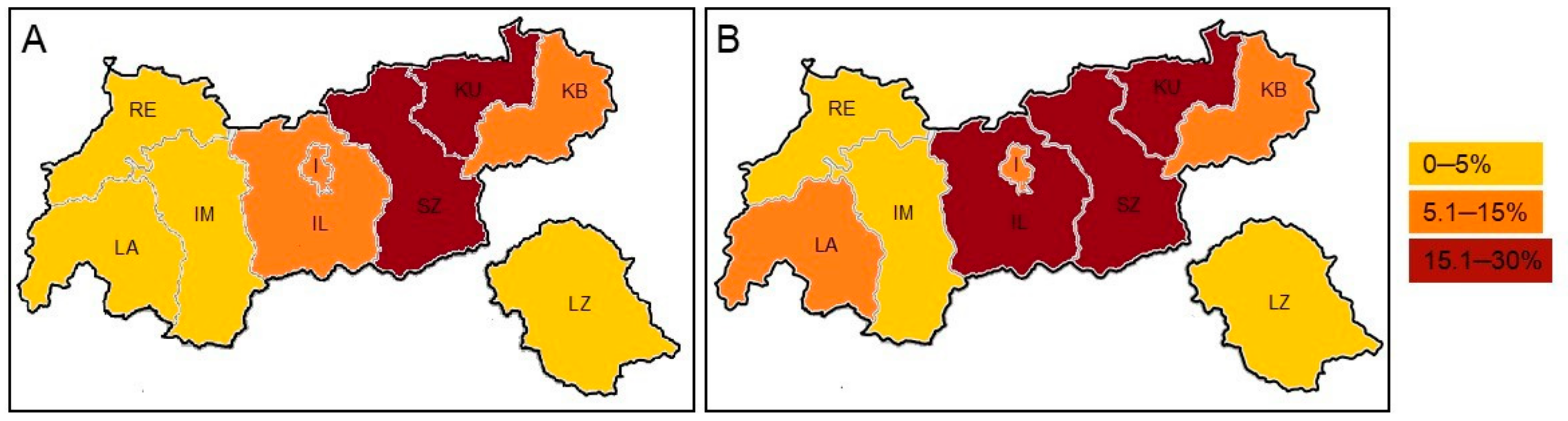
3.3. Environmental Distribution of A. terreus
3.4. A. terreus Isolates from Clinical Samples
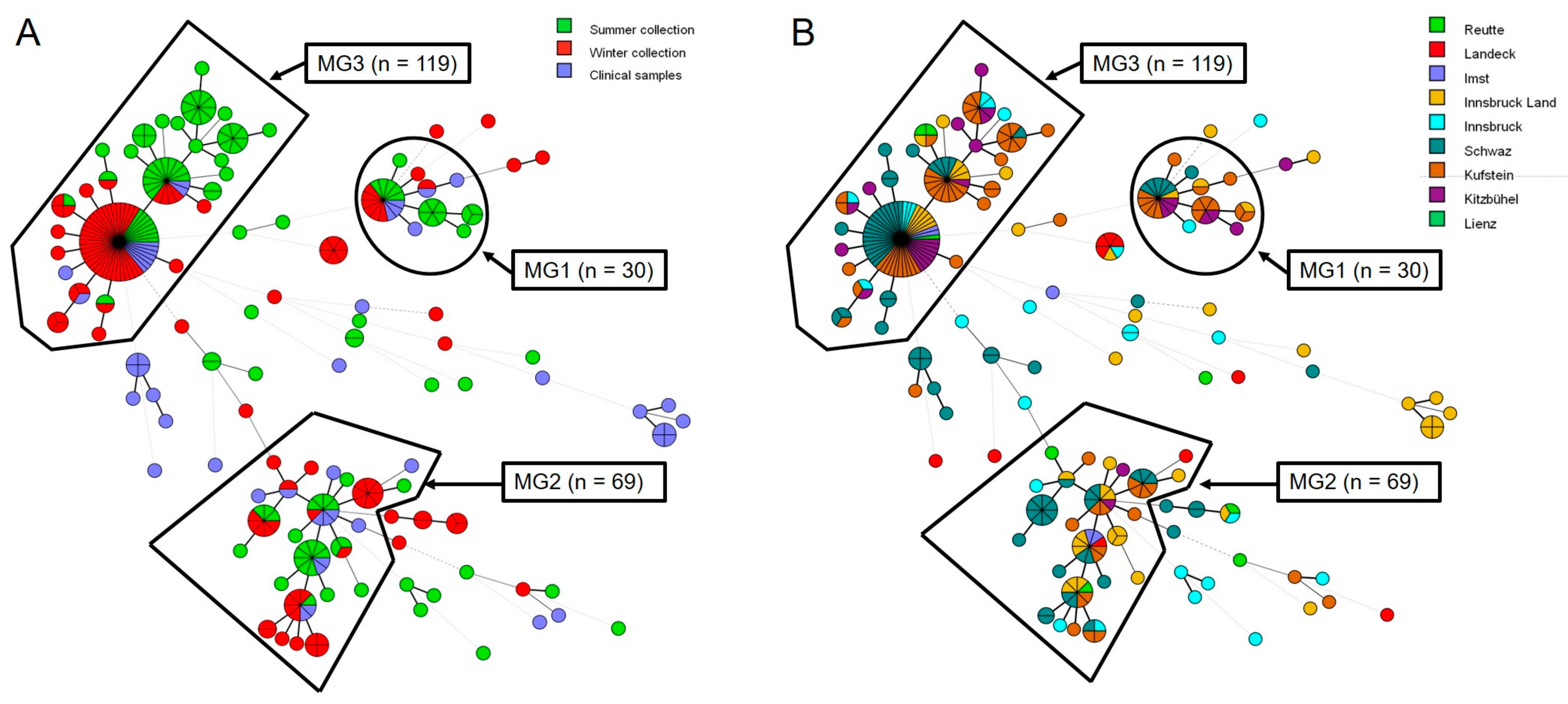
3.5. Microsatellite Typing
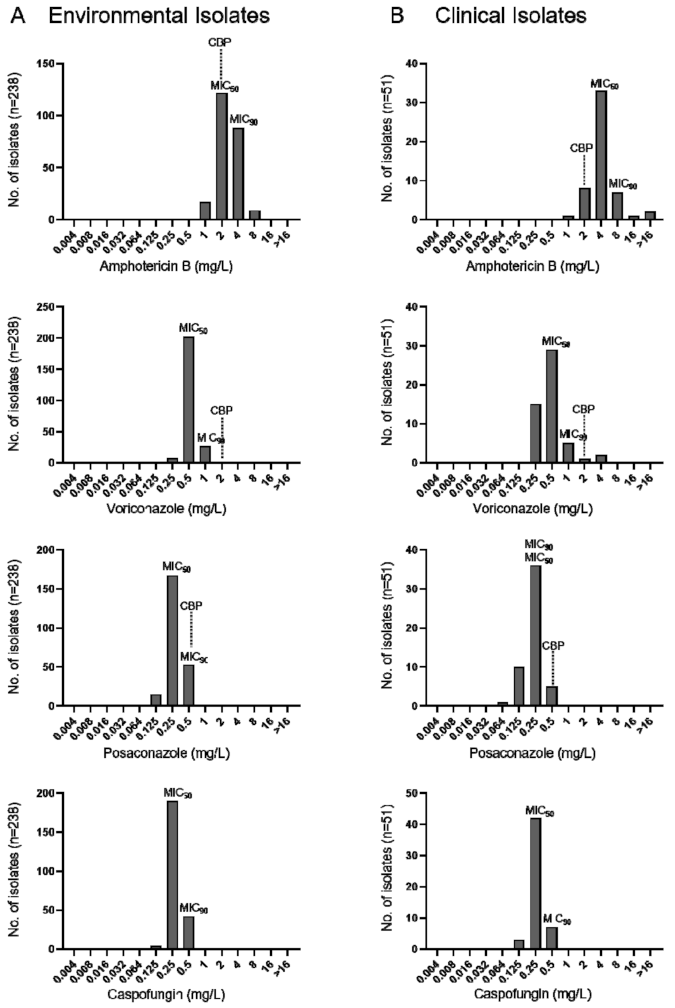
3.6. Antifungal Susceptibility Testing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165. [Google Scholar] [CrossRef] [Green Version]
- Walsh, T.J.; Stevens, D.A. Aspergillosis. In Goldman’s Cecil Medicine; Goldman, L., Schafer, A.I., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2012; Volume 1, pp. 1990–1994. [Google Scholar]
- Steinbach, W.J.; Marr, K.A.; Anaissie, E.J.; Azie, N.; Quan, S.P.; Meier-Kriesche, H.U.; Apewokin, S.; Horn, D.L. Clinical epidemiology of 960 patients with invasive aspergillosis from the PATH Alliance registry. J. Infect. 2012, 65, 453–464. [Google Scholar] [CrossRef]
- Risslegger, B.; Zoran, T.; Lackner, M.; Aigner, M.; Sanchez-Reus, F.; Rezusta, A.; Chowdhary, A.; Taj-Aldeen, S.J.; Arendrup, M.C.; Oliveri, S.; et al. A prospective international Aspergillus terreus survey: An EFISG, ISHAM and ECMM joint study. Clin. Microbiol. Infect. 2017, 23, 776.e1–776.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, M.; Lass-Florl, C. Changing epidemiology of systemic fungal infections. Clin. Microbiol. Infect. 2008, 14, 5–24. [Google Scholar] [CrossRef] [Green Version]
- Lass-Florl, C.; Cuenca-Estrella, M. Changes in the epidemiological landscape of invasive mould infections and disease. J. Antimicrob. Chemother. 2017, 72, i5–i11. [Google Scholar] [CrossRef]
- Perfect, J.R.; Cox, G.M.; Lee, J.Y.; Kauffman, C.A.; de Repentigny, L.; Chapman, S.W.; Morrison, V.A.; Pappas, P.; Hiemenz, J.W.; Stevens, D.A.; et al. The impact of culture isolation of Aspergillus species: A hospital-based survey of aspergillosis. Clin. Infect. Dis. 2001, 33, 1824–1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alastruey-Izquierdo, A.; Mellado, E.; Pelaez, T.; Peman, J.; Zapico, S.; Alvarez, M.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M.; Group, F.S. Population-based survey of filamentous fungi and antifungal resistance in Spain (FILPOP Study). Antimicrob. Agents Chemother. 2013, 57, 4604. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.A.; Peterson, S.W.; Frisvad, J.C.; Varga, J. New species in Aspergillus section Terrei. Stud. Mycol. 2011, 69, 39–55. [Google Scholar] [CrossRef] [Green Version]
- Baddley, J.W.; Pappas, P.G.; Smith, A.C.; Moser, S.A. Epidemiology of Aspergillus terreus at a university hospital. J. Clin. Microbiol. 2003, 41, 5525–5529. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, M.S.; Rojas, F.D.; Cattana, M.E.; Sosa, M.D.; Mangiaterra, M.L.; Giusiano, G.E. Aspergillus terreus complex: An emergent opportunistic agent of onychomycosis. Mycoses 2013, 56, 477–481. [Google Scholar] [CrossRef]
- Lass-Flörl, C.; Griff, K.; Mayr, A.; Petzer, A.; Gastl, G.; Bonatti, H.; Freund, M.; Kropshofer, G.; Dierich, M.P.; Nachbaur, D. Epidemiology and outcome of infections due to Aspergillus terreus: 10-year single centre experience. Br. J. Haematol. 2005, 131, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Laham, M.N.; Carpenter, J.L. Aspergillus terreus, a pathogen capable of causing infective endocarditis, pulmonary mycetoma, and allergic bronchopulmonary aspergillosis. Am. Rev. Respir. Dis. 1982, 125, 769–772. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Agudo, L.; Aznar-Marin, P.; Galan-Sanchez, F.; Garcia-Martos, P.; Marin-Casanova, P.; Rodriguez-Iglesias, M. Otomycosis due to filamentous fungi. Mycopathologia 2011, 172, 307–310. [Google Scholar] [CrossRef]
- Dunne, K.; Prior, A.R.; Murphy, K.; Wall, N.; Leen, G.; Rogers, T.R.; Elnazir, B.; Greally, P.; Renwick, J.; Murphy, P. Emergence of persistent Aspergillus terreus colonisation in a child with cystic fibrosis. Med. Mycol. Case Rep. 2015, 9, 26–30. [Google Scholar] [CrossRef]
- Sabino, R.; Ferreira, J.A.; Moss, R.B.; Valente, J.; Verissimo, C.; Carolino, E.; Clemons, K.V.; Everson, C.; Banaei, N.; Penner, J.; et al. Molecular epidemiology of Aspergillus collected from cystic fibrosis patients. J. Cyst. Fibros. 2015, 14, 474–481. [Google Scholar] [CrossRef] [Green Version]
- Cimon, B.; Zouhair, R.; Symoens, F.; Carrere, J.; Chabasse, D.; Bouchara, J.P. Aspergillus terreus in a cystic fibrosis clinic: Environmental distribution and patient colonization pattern. J. Hosp. Infect. 2003, 53, 81–82. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsube, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Balajee, S.A.; Baddley, J.W.; Peterson, S.W.; Nickle, D.; Varga, J.; Boey, A.; Lass-Florl, C.; Frisvad, J.C.; Samson, R.A. ISHAM Working Group on A. terreus. Aspergillus alabamensis, a new clinically relevant species in the section Terrei. Eukaryot. Cell 2009, 8, 713–722. [Google Scholar] [CrossRef] [Green Version]
- Auberger, J.; Lass-Florl, C.; Clausen, J.; Bellmann, R.; Buzina, W.; Gastl, G.; Nachbaur, D. First case of breakthrough pulmonary Aspergillus niveus infection in a patient after allogeneic hematopoietic stem cell transplantation. Diagn. Microbiol. Infect. Dis. 2008, 62, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Lackner, M.; Obermair, J.; Naschberger, V.; Raschbichler, L.M.; Kandelbauer, C.; Pallua, J.; Metzlaff, J.; Furxer, S.; Lass-Florl, C.; Binder, U. Cryptic species of Aspergillus section Terrei display essential physiological features to cause infection and are similar in their virulence potential in Galleria mellonella. Virulence 2019, 10, 542–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, G.; Perkhofer, S.; Haas, H.; Schrettl, M.; Wurzner, R.; Dierich, M.P.; Lass-Florl, C. Potential basis for amphotericin B resistance in Aspergillus terreus. Antimicrob. Agents Chemother. 2008, 52, 1553–1555. [Google Scholar] [CrossRef] [Green Version]
- Lass-Florl, C.; Alastruey-Izquierdo, A.; Cuenca-Estrella, M.; Perkhofer, S.; Rodriguez-Tudela, J.L. In vitro activities of various antifungal drugs against Aspergillus terreus: Global assessment using the methodology of the European Committee on Antimicrobial Susceptibility Testing. Antimicrob. Agents Chemother. 2009, 53, 794–795. [Google Scholar] [CrossRef] [Green Version]
- Zoran, T.; Sartori, B.; Sappl, L.; Aigner, M.; Sanchez-Reus, F.; Rezusta, A.; Chowdhary, A.; Taj-Aldeen, S.J.; Arendrup, M.C.; Oliveri, S.; et al. Azole-resistance in Aspergillus terreus and related species: An emerging problem or a rare phenomenon? Front. Microbiol. 2018, 9, 516. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Jensen, R.H.; Grif, K.; Skov, M.; Pressler, T.; Johansen, H.K.; Lass-Florl, C. In vivo emergence of Aspergillus terreus with reduced azole susceptibility and a Cyp51a M217I alteration. J. Infect. Dis. 2012, 206, 981–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor, F.J.; Guarro, J. Treatment of Aspergillus terreus infections: A clinical problem not yet resolved. Int. J. Antimicrob. Agents 2014, 44, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, W.J.; Benjamin, D.K., Jr.; Kontoyiannis, D.P.; Perfect, J.R.; Lutsar, I.; Marr, K.A.; Lionakis, M.S.; Torres, H.A.; Jafri, H.; Walsh, T.J. Infections due to Aspergillus terreus: A multicenter retrospective analysis of 83 cases. Clin. Infect. Dis. 2004, 39, 192–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, T.J.; Petraitis, V.; Petraitiene, R.; Field-Ridley, A.; Sutton, D.; Ghannoum, M.; Sein, T.; Schaufele, R.; Peter, J.; Bacher, J.; et al. Experimental pulmonary aspergillosis due to Aspergillus terreus: Pathogenesis and treatment of an emerging fungal pathogen resistant to amphotericin B. J. Infect. Dis 2003, 188, 305–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lass-Flörl, C.; Grif, K.; Kontoyiannis, D.P. Molecular typing of Aspergillus terreus isolates collected in Houston, Texas, and Innsbruck, Austria: Evidence of great genetic diversity. J. Clin. Microbiol. 2007, 45, 2686–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwen, P.C.; Davis, J.C.; Reed, E.C.; Winfield, B.A.; Hinrichs, S.H. Airborne fungal spore monitoring in a protective environment during hospital construction, and correlation with an outbreak of invasive aspergillosis. Infect. Control. Hosp. Epidemiol. 1994, 15, 303–306. [Google Scholar] [CrossRef]
- Oren, I.; Haddad, N.; Finkelstein, R.; Rowe, J.M. Invasive pulmonary aspergillosis in neutropenic patients during hospital construction: Before and after chemoprophylaxis and institution of HEPA filters. Am. J. Hematol. 2001, 66, 257–262. [Google Scholar] [CrossRef]
- Lass-Flörl, C.; Rath, P.; Niederwieser, D.; Kofler, G.; Wurzner, R.; Krezy, A.; Dierich, M.P. Aspergillus terreus infections in haematological malignancies: Molecular epidemiology suggests association with in-hospital plants. J. Hosp. Infect. 2000, 46, 31–35. [Google Scholar] [CrossRef]
- Blum, G.; Perkhofer, S.; Grif, K.; Mayr, A.; Kropshofer, G.; Nachbaur, D.; Kafka-Ritsch, R.; Dierich, M.P.; Lass-Florl, C. A 1-year Aspergillus terreus surveillance study at the University Hospital of Innsbruck: Molecular typing of environmental and clinical isolates. Clin. Microbiol. Infect. 2008, 14, 1146–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lackner, M.; Coassin, S.; Haun, M.; Binder, U.; Kronenberg, F.; Haas, H.; Jank, M.; Maurer, E.; Meis, J.F.; Hagen, F.; et al. Geographically predominant genotypes of Aspergillus terreus species complex in Austria: S microsatellite typing study. Clin. Microbiol. Infect. 2016, 22, 270–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.B.; Cho, H.S.; Shin, H.D.; Frisvad, J.C.; Samson, R.A. Novel Neosartorya species isolated from soil in Korea. Int. J. Syst. Evol. Microbiol. 2006, 56, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Meletiadis, J.; Mouton, J.W.; Lagrou, K.; Hamal, P.; Guinea, J.; The Subcommittee on Antifungal Susceptibility Testing (AFST) of the ESCMID European Committee for Antimicrobial Susceptibility Testing (EUCAST). Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for conidia forming moulds. EUCAST E.Def. 9.3.2 2020, 1–23. Available online: https://www.eucast.org/astoffungi/methodsinantifungalsusceptibilitytesting/ast_of_moulds/ (accessed on 5 March 2021).
- Rougeron, A.; Giraud, S.; Razafimandimby, B.; Meis, J.F.; Bouchara, J.P.; Klaassen, C.H. Different colonization patterns of Aspergillus terreus in patients with cystic fibrosis. Clin. Microbiol. Infect. 2014, 20, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Rüping, M.J.; Gerlach, S.; Fischer, G.; Lass-Florl, C.; Hellmich, M.; Vehreschild, J.J.; Cornely, O.A. Environmental and clinical epidemiology of Aspergillus terreus: Data from a prospective surveillance study. J. Hosp. Infect. 2011, 78, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Guinea, J.; Pelaez, T.; Alcala, L.; Bouza, E. Outdoor environmental levels of Aspergillus spp. conidia over a wide geographical area. Med. Mycol. 2006, 44, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Jaronski, S.T. Ecological factors in the inundative use of fungal entomopathogens. Biocontrol 2010, 55, 159–185. [Google Scholar] [CrossRef]
- Manstretta, V.; Rossi, V. Effects of weather variables on Ascospore discharge from Fusarium graminearum perithecia. PLoS ONE 2015, 10, e0138860. [Google Scholar] [CrossRef] [Green Version]
- Burch, M.; Levetin, E. Effects of meteorological conditions on spore plumes. Int. J. Biometeorol. 2002, 46, 107–117. [Google Scholar] [CrossRef]
- Levetin, E.; Dorsey, K. Contribution of leaf surface fungi to the air spora. Aerobiologia 2006, 22, 3–12. [Google Scholar] [CrossRef]
- Hasnain, S.M. Influence of meteorological factors on the air spora. Grana 1993, 32, 184–188. [Google Scholar] [CrossRef]
- Crandall, S.G.; Gilbert, G.S. Meteorological factors associated with abundance of airborne fungal spores over natural vegetation. Atmos. Environ. 2017, 162, 87–99. [Google Scholar] [CrossRef]
- Langenberg, W.J.; Sutton, J.C.; Gillespie, T.J. Relation of weather variables and periodicities of airborne spores of Alternaria dauci. Phytopathology 1977, 67, 879–883. [Google Scholar] [CrossRef] [Green Version]
- Leyronas, C.; Nicot, P.C. Monitoring viable airborne inoculum of Botrytis cinerea in the South-East of France over 3 years: Relation with climatic parameters and the origin of air masses. Aerobiologia 2013, 29, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Hadi, A.; Magan, N. Influence of physiological factors on growth, sporulation and ochratoxin A/B production of the new Aspergillus ochraceus grouping. World Mycotoxin J. 2009, 2, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Akilandeswari, P.; Paradeep, B.V. Aspergillus terreus KMBF1501 a potential pigment producer under submerged fermentation. Int. J. Pharm. Pharm. Sci. 2017, 9, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Hachem, R.; Gomes, M.Z.; El Helou, G.; El Zakhem, A.; Kassis, C.; Ramos, E.; Jiang, Y.; Chaftari, A.M.; Raad, I.I. Invasive aspergillosis caused by Aspergillus terreus: An emerging opportunistic infection with poor outcome independent of azole therapy. J. Antimicrob. Chemother. 2014, 69, 3148–3155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lass-Florl, C.; Kofler, G.; Kropshofer, G.; Hermans, J.; Kreczy, A.; Dierich, M.P.; Niederwieser, D. In-vitro testing of susceptibility to amphotericin B is a reliable predictor of clinical outcome in invasive aspergillosis. J. Antimicrob. Chemother. 1998, 42, 497–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwen, P.C.; Rupp, M.E.; Langnas, A.N.; Reed, E.C.; Hinrichs, S.H. Invasive pulmonary aspergillosis due to Aspergillus terreus: 12-year experience and review of the literature. Clin. Infect. Dis. 1998, 26, 1092–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendrup, M.C.; Cuenca-Estrella, M.; Lass-Florl, C.; Hope, W.W.; Suscep, E.C.A. EUCAST Technical Note on Aspergillus and amphotericin B, itraconazole, and posaconazole. Clin. Microbiol. Infect. 2012, 18, E248–E250. [Google Scholar] [CrossRef] [Green Version]
- Espinel-Ingroff, A.; Fothergill, A.; Fuller, J.; Johnson, E.; Pelaez, T.; Turnidge, J. Wild-type MIC distributions and epidemiological cutoff values for caspofungin and Aspergillus spp. for the CLSI broth microdilution method (M38-A2 Document). Antimicrob. Agents Chemother. 2011, 55, 2855–2859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A.; Messer, S.A.; Hollis, R.J.; Jones, R.N.; Grp, S.P. Antifungal activities of posaconazole, ravuconazole, and voriconazole compared to those of itraconazole and amphotericin B against 239 clinical isolates of Aspergillus spp. and other filamentous fungi: Report from SENTRY Antimicrobial Surveillance Program, 2000. Antimicrob. Agents Chemother. 2002, 46, 1032–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutton, D.A.; Sanche, S.E.; Revankar, S.G.; Fothergill, A.W.; Rinaldi, M.G. In vitro amphotericin B resistance in clinical isolates of Aspergillus terreus, with a head-to-head comparison to voriconazole. J. Clin. Microbiol. 1999, 37, 2343–2345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, J.; Rimek, D.; Kappe, R. Intrinsic in vitro susceptibility of primary clinical isolates of Aspergillus fumigatus, Aspergillus terreus, Aspergillus nidulans, Candida albicans and Candida lusitaniae against amphotericin B. Mycoses 2006, 49, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Reichert-Lima, F.; Lyra, L.; Pontes, L.; Moretti, M.L.; Pham, C.D.; Lockhart, S.R.; Schreiber, A.Z. Surveillance for azoles resistance in Aspergillus spp. highlights a high number of amphotericin B-resistant isolates. Mycoses 2018, 61, 360–365. [Google Scholar] [CrossRef]
- Kathuria, S.; Sharma, C.; Singh, P.K.; Agarwal, P.; Agarwal, K.; Hagen, F.; Meis, J.F.; Chowdhary, A. Molecular epidemiology and in-vitro antifungal susceptibility of Aspergillus terreus species complex isolates in Delhi, India: Evidence of genetic diversity by amplified fragment length polymorphism and microsatellite typing. PLoS ONE 2015, 10, e0118997. [Google Scholar] [CrossRef]
- Neal, C.O.; Richardson, A.O.; Hurst, S.F.; Tortorano, A.M.; Viviani, M.A.; Stevens, D.A.; Balajee, S.A. Global population structure of Aspergillus terreus inferred by ISSR typing reveals geographical subclustering. BMC Microbiol. 2011, 11, 203. [Google Scholar] [CrossRef] [Green Version]
- Balajee, S.A.; Kano, R.; Baddley, J.W.; Moser, S.A.; Marr, K.A.; Alexander, B.D.; Andes, D.; Kontoyiannis, D.P.; Perrone, G.; Peterson, S.; et al. Molecular identification of Aspergillus species collected for the Transplant-Associated Infection Surveillance Network. J. Clin. Microbiol. 2009, 47, 3138–3141. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Aspergillus Spp. | No. (%) of Positive Samples | ||
|---|---|---|---|
| All (n = 3485) | Summer (n = 1714) | Winter (n = 1771) | |
| Aspergillus fumigatus | 2398 (68.8) | 1138 (66.4) | 1260 (71.1) |
| Aspergillus flavus | 889 (25.5) | 531 (30.1) | 358 (20.2) |
| Aspergillus niger | 887 (25.4) | 453 (26.4) | 434 (24.5) |
| Aspergillus terreus | 189 (5.4) | 68 (3.9) | 121 (6.8) |
| Sample Material | No. (%) of Identified A. Terreus Species Complex | ||
|---|---|---|---|
| All | Summer | Winter | |
| Soil | 127 (67.2) | 55 (80.9) | 72 (59.5) |
| Plant material | 22 (11.6) | 6 (8.8) | 16 (13.2) |
| Air | 20 (10.6) | 4 (5.9) | 16 (13.2) |
| Decaying material | 17 (9) | 3 (4.4) | 14 (11.6) |
| Water | 2 (1.1) | 0 (0) | 2 (1.7) |
| Other | 1 (0.5) | 0 (0) | 1 (0.8) |
| Total | 189/3485 (5.4) | 68/1714 (3.9) | 121/1771 (6.8) |
| District | No. (%) of Identified A. terreus Isolates | ||
|---|---|---|---|
| All (n = 189) | Summer (n = 68) | Winter (n = 121) | |
| RE (Reutte) 1 | 8 (4.2) | 4 (5.9) | 4 (3.3) |
| LA (Landeck) 1 | 7 (3.7) | 3 (4.4) | 4 (3.3) |
| IM (Imst) 1 | 7 (3.7) | 2 (2.9) | 5 (4.1) |
| IL (Innsbruck Land) | 28 (14.8) | 12 (17.6) | 16 (13.2) |
| I (Innsbruck) | 21 (11.1) | 6 (8.8) | 15 (12.4) |
| SZ (Schwaz) 2 | 56 (29.6) | 15 (22.1) | 41 (33.9) |
| KU (Kufstein) 2 | 40 (21.2) | 16 (23.5) | 24 (19.8) |
| KB (Kitzbühel) 2 | 21 (11.1) | 9 (13.2) | 12 (9.9) |
| LZ (Lienz) | 1 (0.5) | 1 (1.5) | 0 (0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dietl, A.-M.; Vahedi-Shahandashti, R.; Kandelbauer, C.; Kraak, B.; Lackner, M.; Houbraken, J.; Lass-Flörl, C. The Environmental Spread of Aspergillus terreus in Tyrol, Austria. Microorganisms 2021, 9, 539. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030539
Dietl A-M, Vahedi-Shahandashti R, Kandelbauer C, Kraak B, Lackner M, Houbraken J, Lass-Flörl C. The Environmental Spread of Aspergillus terreus in Tyrol, Austria. Microorganisms. 2021; 9(3):539. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030539
Chicago/Turabian StyleDietl, Anna-Maria, Roya Vahedi-Shahandashti, Carmen Kandelbauer, Bart Kraak, Michaela Lackner, Jos Houbraken, and Cornelia Lass-Flörl. 2021. "The Environmental Spread of Aspergillus terreus in Tyrol, Austria" Microorganisms 9, no. 3: 539. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030539