
Lake Ecosystem Robustness and Resilience Inferred from a Climate-Stressed Protistan Plankton Network

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
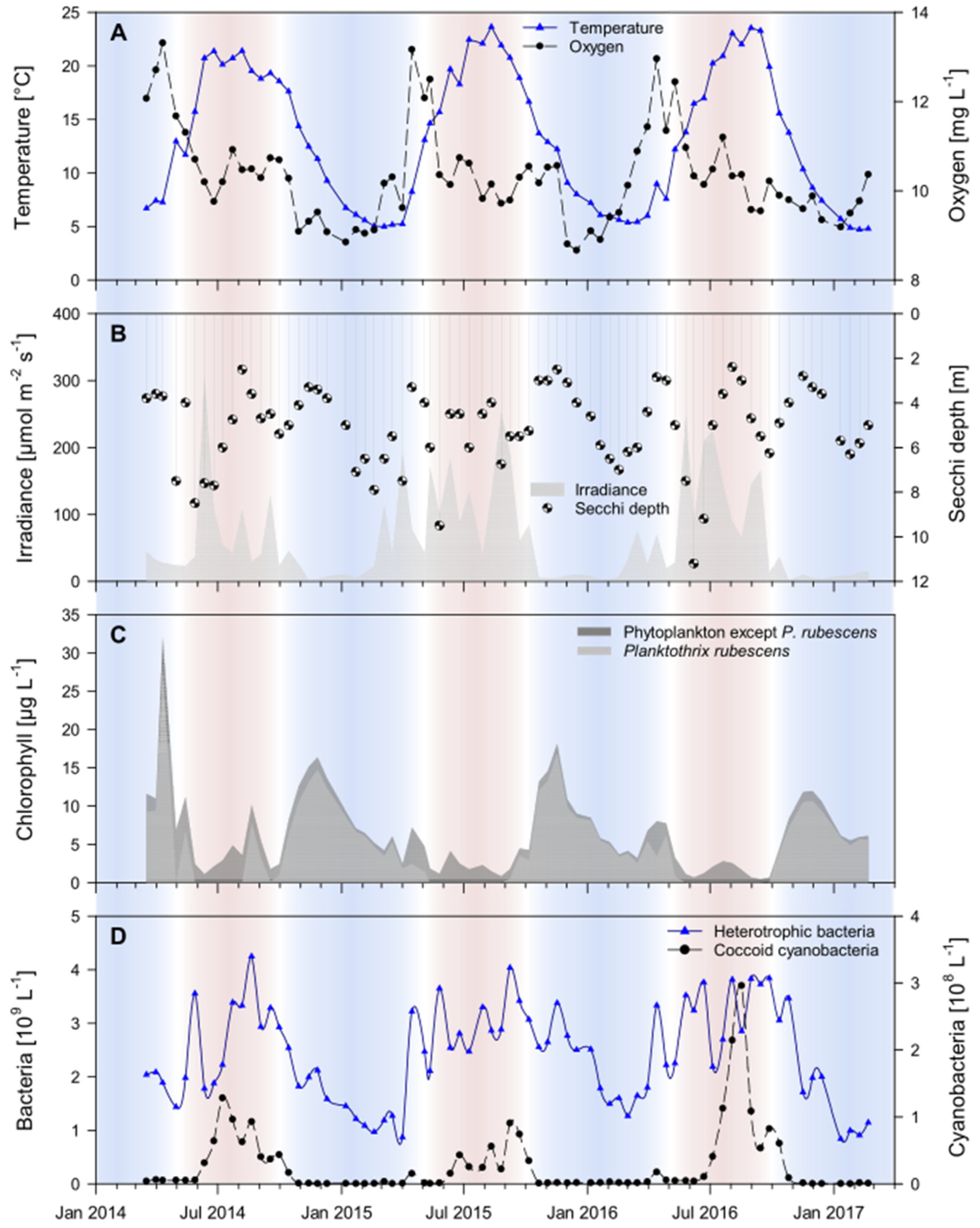
2.1. Sampling Site Information and Measurement of Environmental Parameters
2.2. Sample Processing and High-Throughput Sequencing
2.3. Sequence Quality Control, Clustering, and Taxonomic Assignment
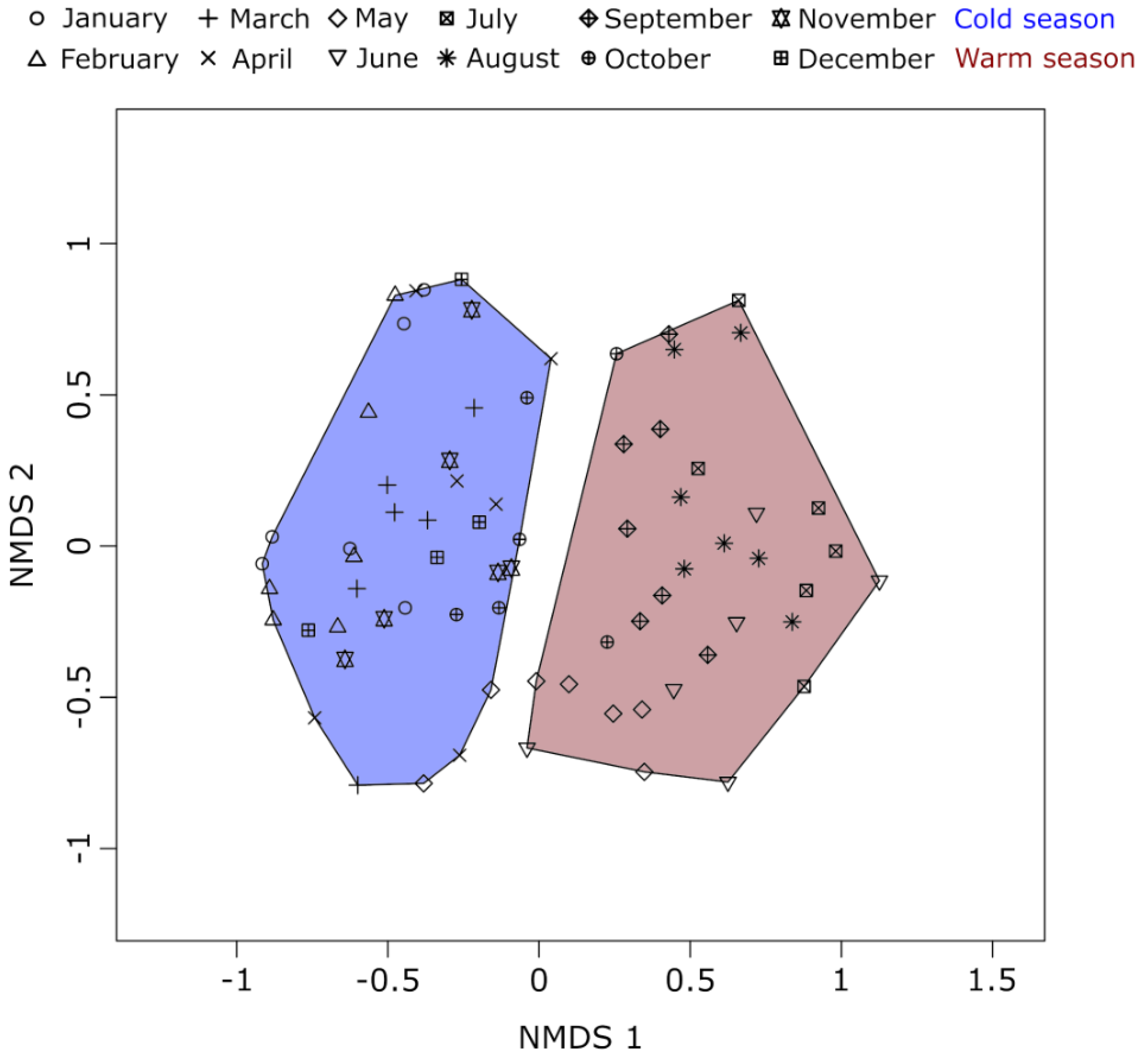
2.4. Compositional Variation Analyses
2.5. Construction of Co-Occurrence Networks
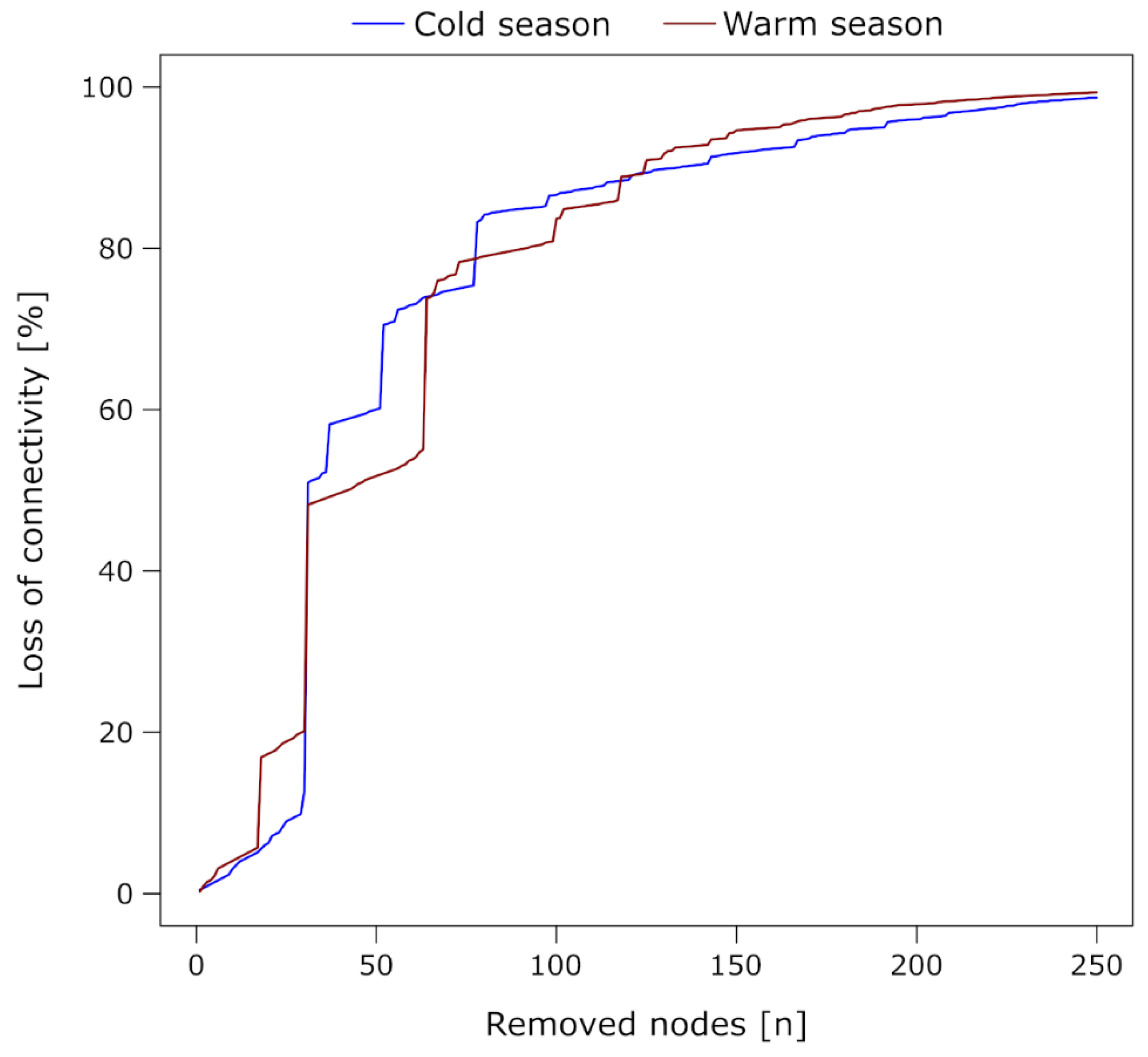
2.6. Evaluation of Patterns within Co-Occurrence Networks
3. Results
3.1. Seasonal Dynamics of Environmental Parameters
3.2. Co-Occurrence Network Properties
3.3. Impact of Environmental Parameters on the Co-Occurrence Networks
3.4. Taxonomic Composition of the Co-Occurrence Networks
3.5. Key Nodes of the Co-Occurrence Networks
4. Discussion
4.1. Placing Co-Occurrence Networks into Perspective
4.2. Succession in the Protistan Plankton Network of Lake Zurich Is Affected by Climate Change
4.3. Ecological Consequences for Different Protist Groups Inferred from Climate-Stressed Networks
4.4. Assessing Ecosystem Resilience of Lake Zurich with Protistan Community Networks
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sherr, B.F.; Sherr, E.B.; Hopkinson, C.S. Trophic Interactions within Pelagic Microbial Communities: Indications of feedback regulation of carbon flow. Hydrobiologia 1988, 159, 19–26. [Google Scholar] [CrossRef]
- Stockner, J.G.; Porter, K.G. Microbial food webs in freshwater planktonic ecosystems. In Complex Interactions in Lake Communities; Carpenter, S.R., Ed.; Springer: New York, NY, USA, 1988; pp. 69–83. ISBN 978-1-4612-3838-6. [Google Scholar]
- Sommer, U.; Gliwicz, Z.M.; Lampert, W.; Duncan, A. The PEG-model of seasonal succession of planktonic events in fresh waters. Arch. Hydrobiol. 1986, 106, 433–471. [Google Scholar]
- Sommer, U.; Adrian, R.; de Senerpont Domis, L.; Elser, J.J.; Gaedke, U.; Ibelings, B.; Jeppesen, E.; Lürling, M.; Molinero, J.C.; Mooij, W.M.; et al. Beyond the Plankton Ecology Group (PEG) Model: Mechanisms driving plankton succession. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 429–448. [Google Scholar] [CrossRef]
- Giner, C.R.; Balagué, V.; Krabberød, A.K.; Ferrera, I.; Reñé, A.; Garcés, E.; Gasol, J.M.; Logares, R.; Massana, R. Quantifying long-term recurrence in planktonic microbial eukaryotes. Mol. Ecol. 2019, 28, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Lallias, D.; Hiddink, J.G.; Fonseca, V.G.; Gaspar, J.M.; Sung, W.; Neill, S.P.; Barnes, N.; Ferrero, T.; Hall, N.; Lambshead, P.J.D.; et al. Environmental metabarcoding reveals heterogeneous drivers of microbial eukaryote diversity in contrasting estuarine ecosystems. ISME J. 2015, 9, 1208–1221. [Google Scholar] [CrossRef] [PubMed]
- Lemke, M.J.; Paver, S.F.; Dungey, K.E.; Velho, L.F.M.; Kent, A.D.; Rodrigues, L.C.; Kellerhals, D.M.; Randle, M.R. Diversity and succession of pelagic microorganism communities in a newly restored Illinois River Floodplain Lake. Hydrobiologia 2017, 804, 35–58. [Google Scholar] [CrossRef] [Green Version]
- Chonova, T.; Kurmayer, R.; Rimet, F.; Labanowski, J.; Vasselon, V.; Keck, F.; Illmer, P.; Bouchez, A. Benthic diatom communities in an alpine river impacted by waste water treatment effluents as revealed using DNA metabarcoding. Front. Microbiol. 2019, 10, 653. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.; López-García, P. Time series are critical to understand microbial plankton diversity and ecology. Mol. Ecol. 2019, 28, 920–922. [Google Scholar] [CrossRef] [PubMed]
- Nolte, V.; Pandey, R.V.; Jost, S.; Medinger, R.; Ottenwälder, B.; Boenigk, J.; Schlötterer, C. Contrasting seasonal niche separation between rare and abundant taxa conceals the extent of protist diversity. Mol. Ecol. 2010, 19, 2908–2915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, M.; López-García, P.; Deschamps, P.; Moreira, D.; Restoux, G.; Bertolino, P.; Jardillier, L. Marked seasonality and high spatial variability of protist communities in shallow freshwater systems. ISME J. 2015, 9, 1941–1953. [Google Scholar] [CrossRef] [Green Version]
- Karimi, B.; Maron, P.A.; Chemidlin-Prevost Boure, N.; Bernard, N.; Gilbert, D.; Ranjard, L. Microbial diversity and ecological networks as indicators of environmental quality. Environ. Chem. Lett. 2017, 15, 265–281. [Google Scholar] [CrossRef]
- Allison, S.D.; Martiny, J.B.H. Resistance, resilience, and redundancy in microbial communities. Proc. Natl. Acad. Sci. USA 2008, 105, 11512–11519. [Google Scholar] [CrossRef] [Green Version]
- Naylor, D.; Sadler, N.; Bhattacharjee, A.; Graham, E.B.; Anderton, C.R.; McClure, R.; Lipton, M.; Hofmockel, K.S.; Jansson, J.K. Soil microbiomes under climate change and implications for carbon cycling. Annu. Rev. Environ. Resour. 2020, 45, 29–59. [Google Scholar] [CrossRef]
- Shade, A.; Peter, H.; Allison, S.D.; Baho, D.; Berga, M.; Buergmann, H.; Huber, D.H.; Langenheder, S.; Lennon, J.T.; Martiny, J.B.; et al. Fundamentals of microbial community resistance and resilience. Front. Microbiol. 2012, 3, 417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connor, N.; Barberán, A.; Clauset, A. Using null models to infer microbial co-occurrence networks. PLoS ONE 2017, 12, e0176751. [Google Scholar] [CrossRef] [Green Version]
- Faust, K.; Lahti, L.; Gonze, D.; de Vos, W.M.; Raes, J. Metagenomics meets time series analysis: Unraveling microbial community dynamics. Curr. Opin. Microbiol. 2015, 25, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Faust, K.; Raes, J. Microbial interactions: From networks to models. Nat. Rev. Microbiol. 2012, 10, 538–550. [Google Scholar] [CrossRef]
- Röttjers, L.; Faust, K. From hairballs to hypotheses–biological insights from microbial networks. FEMS Microbiol. Rev. 2018, 42, 761–780. [Google Scholar] [CrossRef] [Green Version]
- Weiss, S.; Treuren, W.V.; Lozupone, C.; Faust, K.; Friedman, J.; Deng, Y.; Xia, L.C.; Xu, Z.Z.; Ursell, L.; Alm, E.J.; et al. Correlation detection strategies in microbial data sets vary widely in sensitivity and precision. ISME J. 2016, 10, 1669–1681. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, J.A.; Cram, J.A.; Needham, D.M. Marine microbial community dynamics and their ecological interpretation. Nat. Rev. Microbiol. 2015, 13, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Vacher, C.; Tamaddoni-Nezhad, A.; Kamenova, S.; Peyrard, N.; Moalic, Y.; Sabbadin, R.; Schwaller, L.; Chiquet, J.; Smith, M.A.; Vallance, J.; et al. Chapter one-learning ecological networks from next-generation sequencing data. In Ecosystem Services: From Biodiversity to Society, Part 2. Advances in Ecological Research; Woodward, G., Bohan, D.A., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 54, pp. 1–39. [Google Scholar]
- Mandakovic, D.; Rojas, C.; Maldonado, J.; Latorre, M.; Travisany, D.; Delage, E.; Bihouée, A.; Jean, G.; Díaz, F.P.; Fernández-Gómez, B.; et al. Structure and co-occurrence patterns in microbial communities under acute environmental stress reveal ecological factors fostering resilience. Sci. Rep. 2018, 8, 5875. [Google Scholar] [CrossRef]
- De Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.M.; Kretzschmar, M.; et al. Soil bacterial networks are less stable under drought than fungal networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.M.; Kitson, J.J.N.; Lunt, D.H.; Straw, N.A.; Pocock, M.J.O. Merging DNA metabarcoding and ecological network analysis to understand and build resilient terrestrial ecosystems. Funct. Ecol. 2016, 30, 1904–1916. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, R.; Zheng, Q.; Deng, Y.; van Nostrand, J.D.; Zhou, J.; Jiao, N. Bacterioplankton community resilience to ocean acidification: Evidence from microbial network analysis. ICES J. Mar. Sci. 2016, 73, 865–875. [Google Scholar] [CrossRef]
- Araújo, M.B.; Rozenfeld, A.; Rahbek, C.; Marquet, P.A. Using species co-occurrence networks to assess the impacts of climate change. Ecography 2011, 34, 897–908. [Google Scholar] [CrossRef]
- Berry, D.; Widder, S. Deciphering microbial interactions and detecting keystone species with co-occurrence networks. Front. Microbiol. 2014, 5, 219. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Schlaeppi, K.; van der Heijden, M.G.A. Keystone taxa as drivers of microbiome structure and functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Bashan, A.; Shi, D.-N.; Liu, Y.-Y. Articulation points in complex networks. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Clauset, A.; Newman, M.E.J.; Moore, C. Finding community structure in very large networks. Phys. Rev. E 2004, 70, 066111. [Google Scholar] [CrossRef] [Green Version]
- Blondel, V.D.; Guillaume, J.-L.; Lambiotte, R.; Lefebvre, E. Fast unfolding of communities in large networks. J. Stat. Mech. 2008, 10, P10008. [Google Scholar] [CrossRef] [Green Version]
- Chaffron, S.; Rehrauer, H.; Pernthaler, J.; von Mering, C. A Global network of coexisting microbes from environmental and whole-genome sequence data. Genome Res. 2010, 20, 947–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freilich, M.A.; Wieters, E.; Broitman, B.R.; Marquet, P.A.; Navarrete, S.A. Species co-occurrence networks: Can they reveal trophic and non-trophic interactions in ecological communities? Ecology 2018, 99, 690–699. [Google Scholar] [CrossRef] [PubMed]
- Dini-Andreote, F.; de Cássia Pereira e Silva, M.; Triadó-Margarit, X.; Casamayor, E.O.; van Elsas, J.D.; Salles, J.F. Dynamics of bacterial community succession in a salt marsh chronosequence: Evidences for Temporal niche partitioning. ISME J. 2014, 8, 1989–2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faust, K.; Lima-Mendez, G.; Lerat, J.-S.; Sathirapongsasuti, J.F.; Knight, R.; Huttenhower, C.; Lenaerts, T.; Raes, J. Cross-Biome comparison of microbial association networks. Front. Microbiol. 2015, 6, 1200. [Google Scholar] [CrossRef] [Green Version]
- Peura, S.; Bertilsson, S.; Jones, R.I.; Eiler, A. Resistant microbial cooccurrence patterns inferred by network topology. Appl. Environ. Microbiol. 2015, 81, 2090–2097. [Google Scholar] [CrossRef] [Green Version]
- Albert, R.; Jeong, H.; Barabási, A.-L. Error and attack tolerance of complex networks. Nature 2000, 406, 378–382. [Google Scholar] [CrossRef] [Green Version]
- Holme, P.; Kim, B.J.; Yoon, C.N.; Han, S.K. Attack vulnerability of complex networks. Phys. Rev. E 2002, 65, 056109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klau, G.W.; Weiskircher, R. Robustness and resilience. In Network Analysis; Brandes, U., Erlebach, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 417–437. ISBN 978-3-540-31955-9. [Google Scholar]
- Gao, J.; Barzel, B.; Barabási, A.-L. Universal resilience patterns in complex networks. Nature 2016, 530, 307–312. [Google Scholar] [CrossRef]
- Kéfi, S.; Miele, V.; Wieters, E.A.; Navarrete, S.A.; Berlow, E.L. How structured is the entangled bank? The surprisingly simple organization of multiplex ecological networks leads to increased persistence and resilience. PLoS Biol. 2016, 14, e1002527. [Google Scholar] [CrossRef]
- Needham, D.M.; Sachdeva, R.; Fuhrman, J.A. Ecological dynamics and co-occurrence among marine phytoplankton, bacteria and myoviruses shows microdiversity matters. ISME J. 2017, 11, 1614–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupatini, M.; Suleiman, A.K.A.; Jacques, R.J.S.; Antoniolli, Z.I.; de Siqueira Ferreira, A.; Kuramae, E.E.; Roesch, L.F.W. Network topology reveals high connectance levels and few key microbial genera within soils. Front. Environ. Sci. 2014, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Yankova, Y.; Neuenschwander, S.; Köster, O.; Posch, T. Abrupt stop of deep water turnover with lake warming: drastic consequences for algal primary producers. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yankova, Y.; Villiger, J.; Pernthaler, J.; Schanz, F.; Posch, T. Prolongation, deepening and warming of the metalimnion change habitat conditions of the harmful Filamentous Cyanobacterium Planktothrix Rubescens in a prealpine lake. Hydrobiologia 2016, 776, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Posch, T.; Köster, O.; Salcher, M.M.; Pernthaler, J. Harmful Filamentous Cyanobacteria favoured by reduced water turnover with lake warming. Nat. Clim. Chang. 2012, 2, 809–813. [Google Scholar] [CrossRef] [Green Version]
- Livingstone, D.M. Impact of secular climate change on the thermal structure of a large temperate central european lake. Clim. Chang. 2003, 57, 205–225. [Google Scholar] [CrossRef]
- North, R.P.; North, R.L.; Livingstone, D.M.; Köster, O.; Kipfer, R. Long-term changes in hypoxia and soluble reactive phosphorus in the hypolimnion of a large temperate lake: Consequences of a climate regime shift. Glob. Chang. Biol. 2014, 20, 811–823. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.; Forster, D.; Bruni, E.P.; Frantal, D.; Kammerlander, B.; Nachbaur, L.; Pitsch, G.; Posch, T.; Pröschold, T.; Teubner, K.; et al. Aquatic food webs in deep temperate lakes: key species establish through their autecological versatility. Mol. Ecol. 2021, 30, 1053–1071. [Google Scholar] [CrossRef] [PubMed]
- Petrou, K.; Nielsen, D.A. Uptake of Dimethylsulphoniopropionate (DMSP) by the diatom Thalassiosira Weissflogii: A model to investigate the cellular function of DMSP. Biogeochemistry 2018, 141, 265–271. [Google Scholar] [CrossRef]
- Gasol, J.M.; Zweifel, U.L.; Peters, F.; Fuhrman, J.A.; Hagström, Å. Significance of size and nucleic acid content heterogeneity as measured by flow cytometry in natural planktonic bacteria. Appl. Environ. Microbiol. 1999, 65, 4475–4483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoeck, T.; Bass, D.; Nebel, M.; Christen, R.; Jones, M.D.M.; Breiner, H.-W.; Richards, T.A. Multiple marker parallel tag environmental DNA sequencing reveals a highly complex eukaryotic community in marine anoxic water. Mol. Ecol. 2010, 19, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: Chichester, UK, 1991; pp. 115–175. ISBN 0-471-92906-9. [Google Scholar]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified Eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of Chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Forster, D.; Lentendu, G.; Filker, S.; Dubois, E.; Wilding, T.A.; Stoeck, T. Improving eDNA-based protist diversity assessments using networks of amplicon sequence variants. Environ. Microbiol. 2019, 21, 4109–4125. [Google Scholar] [CrossRef] [PubMed]
- Mahé, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm v2: Highly-scalable and high-resolution amplicon clustering. PeerJ 2015, 3, e1420. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the classification, nomenclature, and diversity of eukaryotes. J. Eukaryot. Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef] [Green Version]
- Guillou, L.; Bachar, D.; Audic, S.; Bass, D.; Berney, C.; Bittner, L.; Boutte, C.; Burgaud, G.; de Vargas, C.; Decelle, J.; et al. The Protist Ribosomal Reference Database (PR2): A catalog of unicellular eukaryote small sub-unit RRNA sequences with curated taxonomy. Nucleic Acids Res. 2012, 41, D597–D604. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R. A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 29 January 2021).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P. Vegan: Community Ecology Package. Available online: https://cran.r-project.org/web/packages/vegan/ (accessed on 29 January 2021).
- Lentendu, G.; Dunthorn, M. Relating network analyses to phylogenetic relatedness to infer protistan co-occurrences and co-exclusions in marine and terrestrial environments. bioRxiv 2020. [Google Scholar] [CrossRef]
- Gloor, G.B.; Macklaim, J.M.; Pawlowsky-Glahn, V.; Egozcue, J.J. Microbiome datasets are compositional: and this is not optional. Front. Microbiol. 2017, 8, 2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. In Proceedings of the Third International AAAI Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Csardi, G.; Nepusz, T. The Igraph Software package for complex network research. InterJ. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Canty, A.; Ripley, B. Boot: Bootstrap Functions. Available online: https://cran.r-project.org/web/packages/boot/ (accessed on 29 January 2021).
- Lhomme, S. NetSwan: Network Strengths and Weaknesses Analysis. Available online: https://cran.r-project.org/web/packages/NetSwan/ (accessed on 29 January 2021).
- Blanchet, F.G.; Cazelles, K.; Gravel, D. Co-Occurrence Is Not Evidence of Ecological Interactions. Ecol. Lett. 2020, 23, 1050–1063. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.; Diener, C.; Baliga, N.S.; Gibbons, S.M. Use and abuse of correlation analyses in microbial ecology. ISME J. 2019, 13, 2647–2655. [Google Scholar] [CrossRef] [Green Version]
- Cruaud, P.; Vigneron, A.; Fradette, M.-S.; Dorea, C.C.; Culley, A.I.; Rodriguez, M.J.; Charette, S.J. Annual protist community dynamics in a freshwater ecosystem undergoing contrasted climatic conditions: The Saint-Charles River (Canada). Front. Microbiol. 2019, 10, 2359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teubner, K. Synchronised changes of planktonic cyanobacterial and diatom assemblages in North German waters reduce seasonality to two principal periods. Arch. Hydrobiol. Spec. Issues Advanc. Limnol. 2000, 55, 565–580. [Google Scholar]
- Komárková, J.; Komárek, O.; Hejzlar, J. Evaluation of the long term monitoring of phytoplankton assemblages in a canyon-shape reservoir using multivariate statistical methods. Hydrobiologia 2003, 504, 143–157. [Google Scholar] [CrossRef]
- Eckert, E.M.; Salcher, M.M.; Posch, T.; Eugster, B.; Pernthaler, J. Rapid successions affect microbial N-Acetyl-Glucosamine uptake patterns during a lacustrine spring phytoplankton bloom. Environ. Microbiol. 2012, 14, 794–806. [Google Scholar] [CrossRef]
- van den Wyngaert, S.; Salcher, M.M.; Pernthaler, J.; Zeder, M.; Posch, T. Quantitative dominance of seasonally persistent Filamentous Cyanobacteria (Planktothrix Rubescens) in the microbial assemblages of a temperate lake. Limnol. Oceanogr. 2011, 56, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Sime-Ngando, T. Phytoplankton Chytridiomycosis: Fungal parasites of phytoplankton and their imprints on the food web dynamics. Front. Microbiol. 2012, 3, 361. [Google Scholar] [CrossRef] [Green Version]
- van den Wyngaert, S.; Vanholsbeeck, O.; Spaak, P.; Ibelings, B.W. Parasite fitness traits under environmental variation: Disentangling the roles of a Chytrid’s immediate host and external environment. Microb. Ecol. 2014, 68, 645–656. [Google Scholar] [CrossRef] [Green Version]
- Ibelings, B.W.; Gsell, A.S.; Mooij, W.M.; Donk, E.V.; van den Wyngaert, S.; Domis, L.N.D.S. Chytrid infections and diatom spring blooms: Paradoxical effects of climate warming on fungal epidemics in lakes. Freshw. Biol. 2011, 56, 754–766. [Google Scholar] [CrossRef]
- Posch, T.; Eugster, B.; Pomati, F.; Pernthaler, J.; Pitsch, G.; Eckert, E.M. Network of interactions between ciliates and phytoplankton during spring. Front. Microbiol. 2015, 6, 1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skogstad, A.; Granskog, L.; Klaveness, D. Growth of freshwater ciliates offered planktonic algae as food. J. Plankton Res. 1987, 9, 503–512. [Google Scholar] [CrossRef]
- Jürgens, K.; Wickham, S.A.; Rothhaupt, K.O.; Santer, B. Feeding rates of macro- and microzooplankton on heterotrophic nanoflagellates. Limnol. Oceanogr. 1996, 41, 1833–1839. [Google Scholar] [CrossRef]
- Šimek, K.; Grujčić, V.; Nedoma, J.; Jezberová, J.; Šorf, M.; Matoušů, A.; Pechar, L.; Posch, T.; Bruni, E.P.; Vrba, J. Microbial food webs in hypertrophic fishponds: Omnivorous ciliate taxa are major protistan bacterivores. Limnol. Oceanogr. 2019, 64, 2295–2309. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.J.; Jack, J.D. Rotifers as Predators on SMALL CILIATES. In Rotifer Symposium VI: Proceedings of the Sixth International Rotifer Symposium, Banyoles, Spain, 3–8 June 1991; Gilbert, J.J., Lubzens, E., Miracle, M.R., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 247–253. [Google Scholar]
- Wickham, S.A. Cyclops Predation on ciliates: Species. Specific differences and functional responses. J. Plankton Res. 1995, 17, 1633–1646. [Google Scholar] [CrossRef]
- Jürgens, K.; Skibbe, O.; Jeppesen, E. Impact of metazooplankton on the composition and population dynamics of planktonic ciliates in a shallow, hypertrophic lake. Aquat. Microb. Ecol. 1999, 17, 61–75. [Google Scholar] [CrossRef]
- Grossmann, L.; Beisser, D.; Bock, C.; Chatzinotas, A.; Jensen, M.; Preisfeld, A.; Psenner, R.; Rahmann, S.; Wodniok, S.; Boenigk, J. Trade-off between taxon diversity and functional diversity in european lake ecosystems. Mol. Ecol. 2016, 25, 5876–5888. [Google Scholar] [CrossRef] [PubMed]
- Gerea, M.; Queimaliños, C.; Unrein, F. Grazing impact and prey selectivity of picoplanktonic cells by mixotrophic flagellates in oligotrophic lakes. Hydrobiologia 2019, 831, 5–21. [Google Scholar] [CrossRef]
- Simons, A.L.; Mazor, R.; Theroux, S. Using co-occurrence network topology in assessing ecological stress in benthic macroinvertebrate communities. Ecol. Evol. 2019, 9, 12789–12801. [Google Scholar] [CrossRef] [PubMed]
- Dunne, J.A.; Williams, R.J.; Martinez, N.D. Network structure and biodiversity loss in food webs: Robustness Increases with connectance. Ecol. Lett. 2002, 5, 558–567. [Google Scholar] [CrossRef] [Green Version]
- Tylianakis, J.M.; Laliberté, E.; Nielsen, A.; Bascompte, J. Conservation of species interaction networks. Biol. Conserv. 2010, 143, 2270–2279. [Google Scholar] [CrossRef]
- Biggs, C.R.; Yeager, L.A.; Bolser, D.G.; Bonsell, C.; Dichiera, A.M.; Hou, Z.; Keyser, S.R.; Khursigara, A.J.; Lu, K.; Muth, A.F.; et al. Does functional redundancy affect ecological stability and resilience? A review and meta-analysis. Ecosphere 2020, 11, e03184. [Google Scholar] [CrossRef]
- Jeppesen, E.; Mehner, T.; Winfield, I.J.; Kangur, K.; Sarvala, J.; Gerdeaux, D.; Rask, M.; Malmquist, H.J.; Holmgren, K.; Volta, P.; et al. Impacts of climate warming on the long-term dynamics of key fish species in 24 european lakes. Hydrobiologia 2012, 694, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Grafton, R.Q.; Pittock, J.; Davis, R.; Williams, J.; Fu, G.; Warburton, M.; Udall, B.; McKenzie, R.; Yu, X.; Che, N.; et al. Global insights into water resources, climate change and governance. Nat. Clim. Change 2013, 3, 315–321. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Average (Minimum–Maximum) | Unit |
|---|---|---|
| Water temperature | 12.7 (4.7–23.7) | °C |
| Air temperature | 11.8 (−6.0–24.6) | °C |
| Secchi depth (water transparency) | 5.0 (2.4–11.2) | m |
| Conductivity | 266 (219–293) | µS cm−1 |
| Oxygen concentration | 10.6 (8.7–13.3) | mg O2 L−1 |
| Oxygen saturation | 100 (72–126) | % |
| Orthophosphate * | 1.6 (0.0–3.2) | µg P L−1 |
| Total phosphorus * | 12.3 (7.0–25) | µg P L−1 |
| Particulate phosphorus * | 8.5 (3.0–21) | µg P L−1 |
| Nitrate (NO3-N) * | 434 (113–610) | µg N L−1 |
| Ammonium (NH4-N) * | 6.0 (2.3–22.6) | µg N L−1 |
| Dissolved organic carbon (DOC) * | 1.4 (1.1–1.7) | mg C L−1 |
| Total chlorophyll a | 6.6 (0.5–32.1) | µg Chl a L−1 |
| Maximal depth | 136 | m |
| Total lake volume | 3.3 | km3 |
| Total lake area | 66.6 | km2 |
| Water retention time | 1.2 | years |
| Cold Season Network | Warm Season Network | |
|---|---|---|
| Input samples | 38 | 35 |
| Input NSCs | 21,667 | 23,904 |
| Spearman’s rho co-exclusion threshold | −0.59 | −0.61 |
| Spearman’s rho co-occurrence threshold | 0.6 | 0.62 |
| Edges (co-occurrences) | 6872 | 5252 |
| Nodes (NSCs) | 924 | 963 |
| Nodes of environmental parameters | 11 | 5 |
| Average degree | 14.53 | 10.79 |
| Average path length | 4.64 | 5.46 |
| Connected components (larger than 3 nodes) | 41 [6] | 68 [7] |
| Density | 0.015 | 0.011 |
| Diameter | 12 | 17 |
| Modularity | 0.02 | 0.03 |
| Transitivity | 0.49 | 0.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forster, D.; Qu, Z.; Pitsch, G.; Bruni, E.P.; Kammerlander, B.; Pröschold, T.; Sonntag, B.; Posch, T.; Stoeck, T. Lake Ecosystem Robustness and Resilience Inferred from a Climate-Stressed Protistan Plankton Network. Microorganisms 2021, 9, 549. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030549
Forster D, Qu Z, Pitsch G, Bruni EP, Kammerlander B, Pröschold T, Sonntag B, Posch T, Stoeck T. Lake Ecosystem Robustness and Resilience Inferred from a Climate-Stressed Protistan Plankton Network. Microorganisms. 2021; 9(3):549. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030549
Chicago/Turabian StyleForster, Dominik, Zhishuai Qu, Gianna Pitsch, Estelle P. Bruni, Barbara Kammerlander, Thomas Pröschold, Bettina Sonntag, Thomas Posch, and Thorsten Stoeck. 2021. "Lake Ecosystem Robustness and Resilience Inferred from a Climate-Stressed Protistan Plankton Network" Microorganisms 9, no. 3: 549. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030549