Insertional Inactivation of Prevotella intermedia OxyR Results in Reduced Survival with Oxidative Stress and in the Presence of Host Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
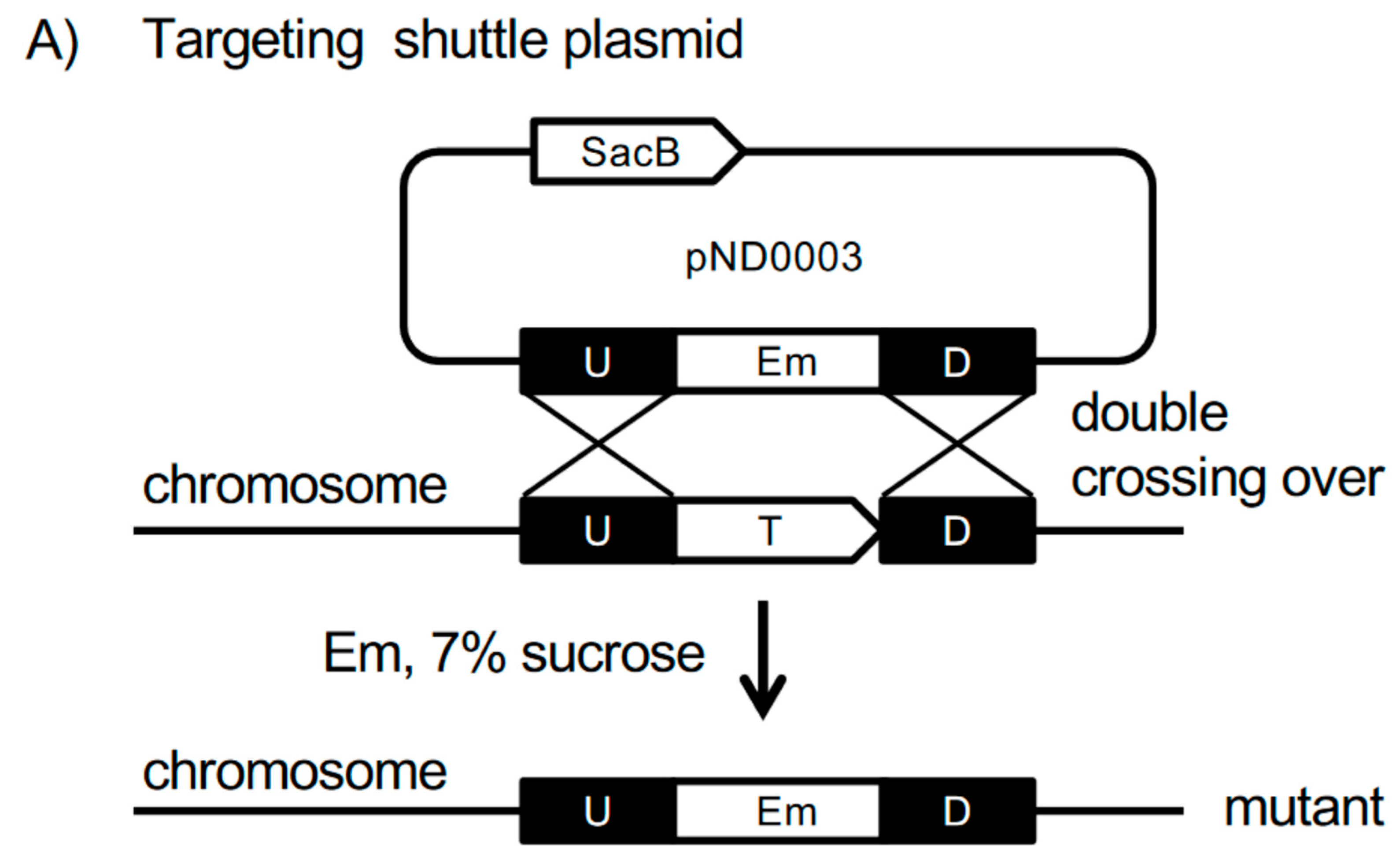
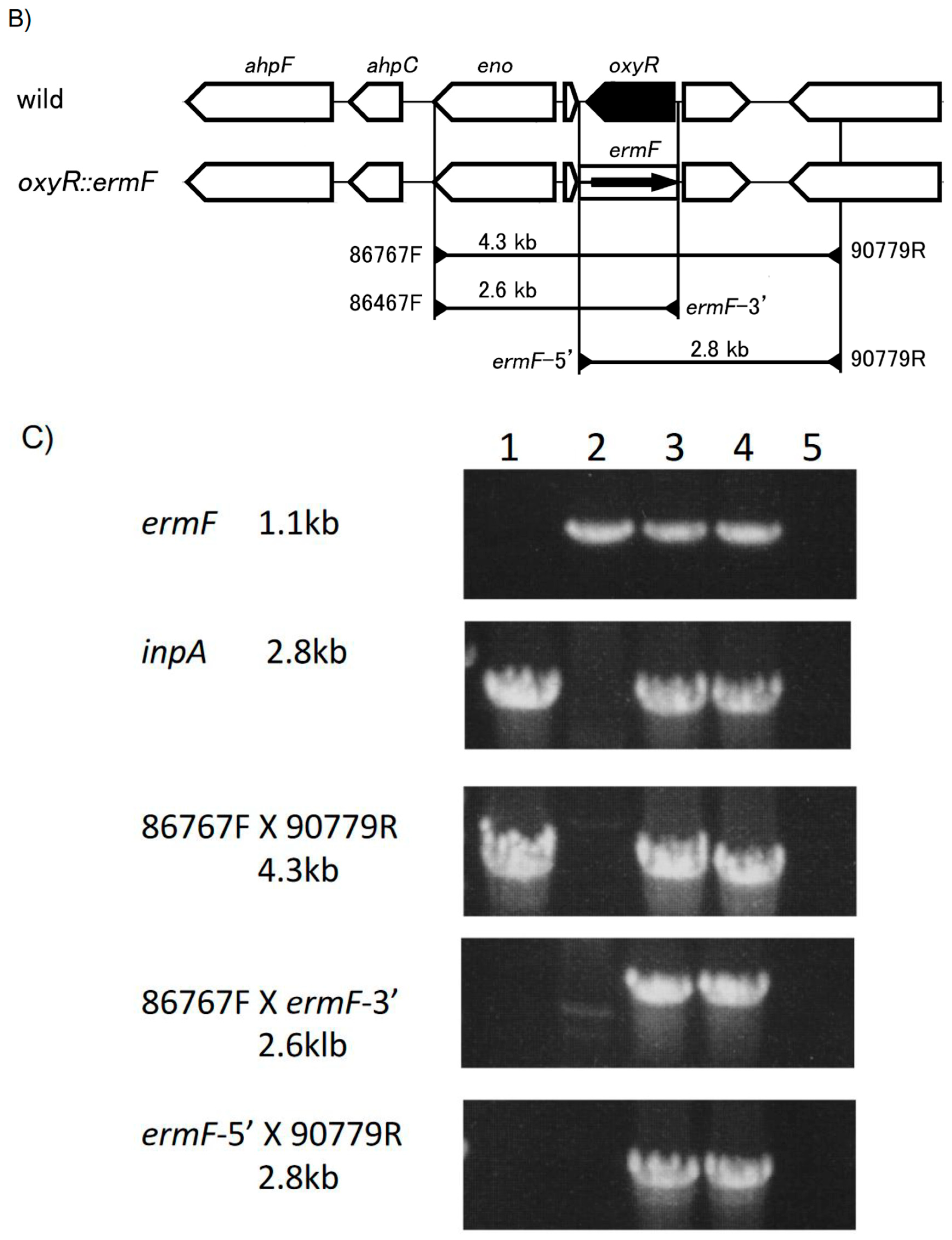
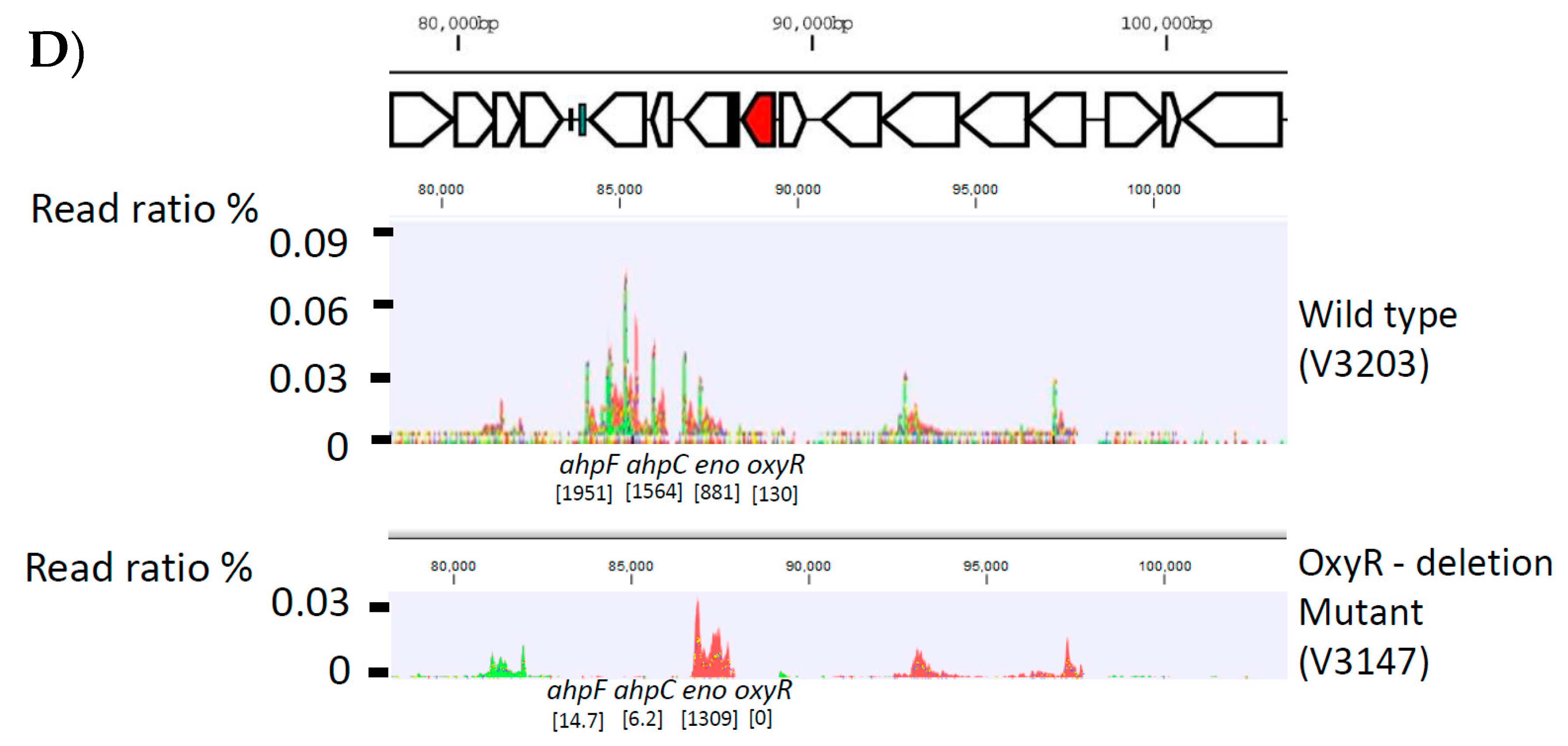
2.2. Generation of P. intermedia Mutant Strain
2.3. RNAseq Analysis of P. intermedia
2.4. Quantitative RT-PCR Analysis
2.5. Disc Diffusion Assay
2.6. Growth under Micro-Aerobic Conditions
2.7. Outgrowth after Aerobic Exposure
2.8. Interaction of P. intermedia with Host Cells
2.9. Statistical Analysis.
3. Results
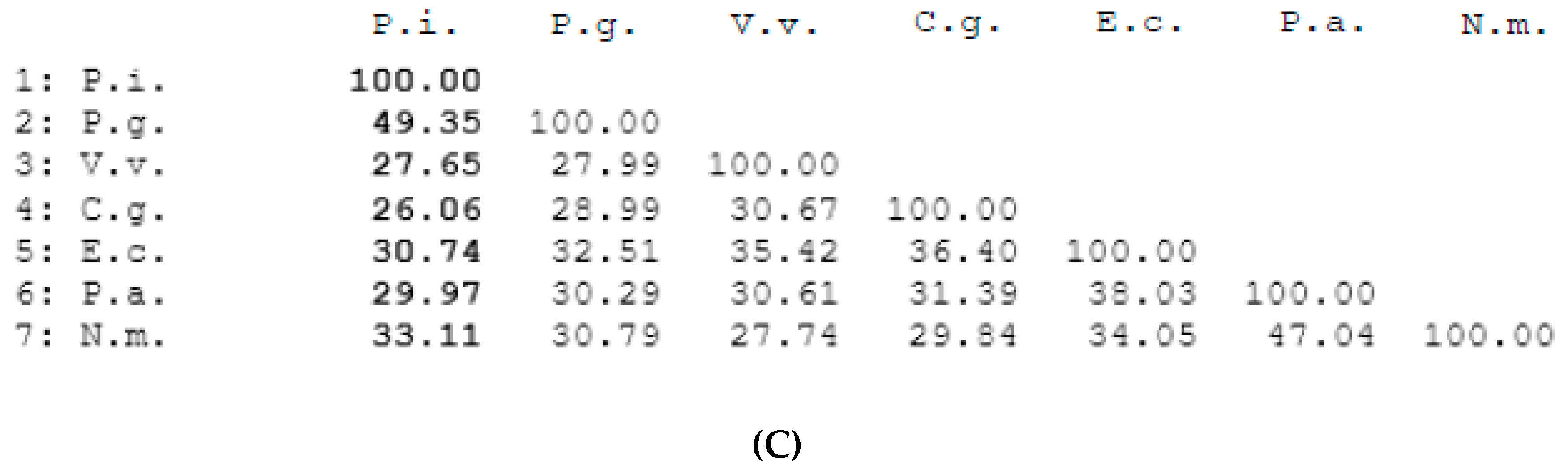
3.1. Bioinformatics Analysis of P. intermedia’s OxyR Ortholog
3.2. Generation of P. intermedia Insertional Deletion Mutant Strain
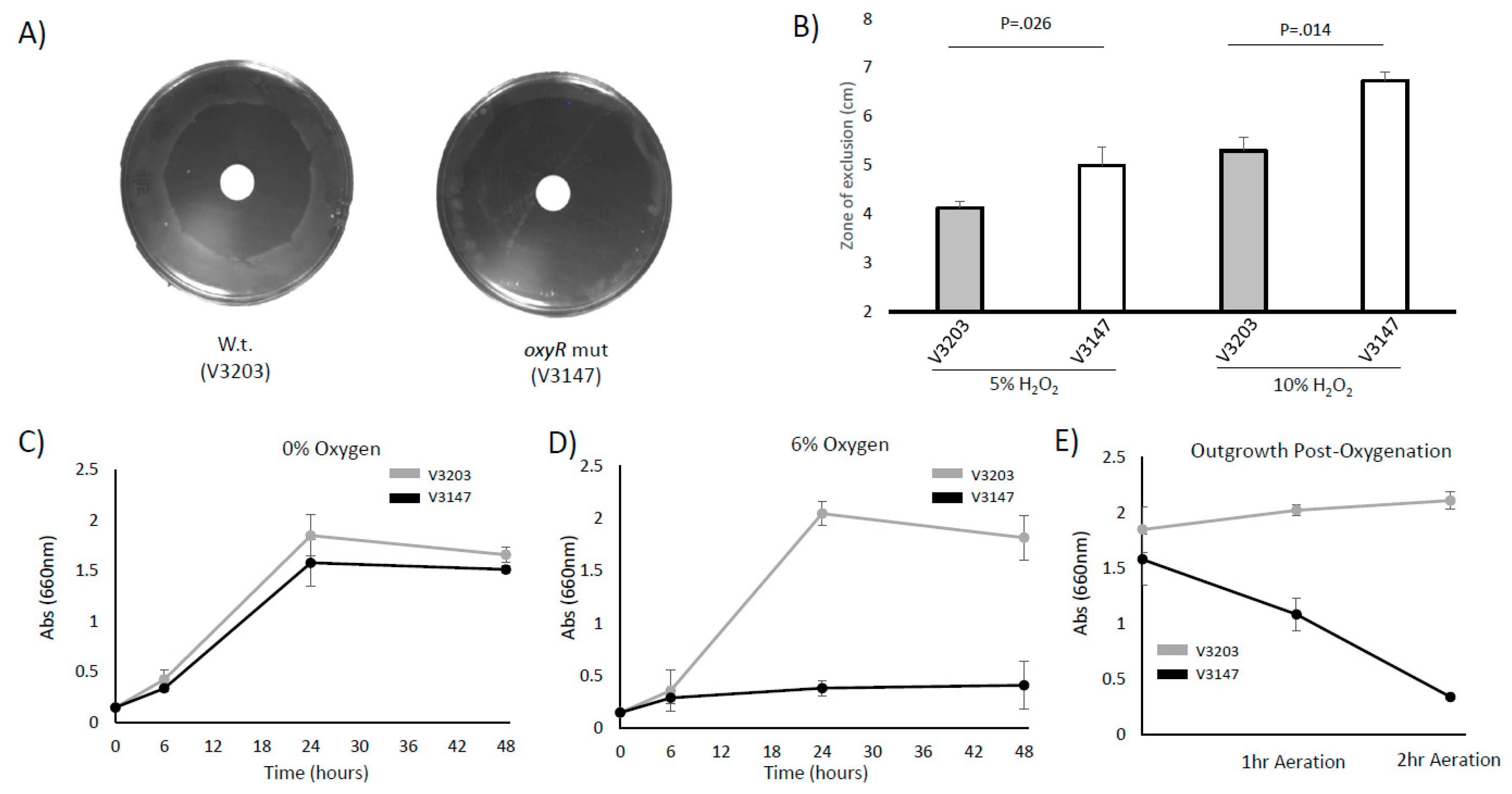
3.3. OxyR Plays a Role in Oxidative Stress Homeostasis in P. intermedia
3.4. P. intermedia Requires OxyR to Grow and Survive with Oxidative Stress and Oxygenated Conditions
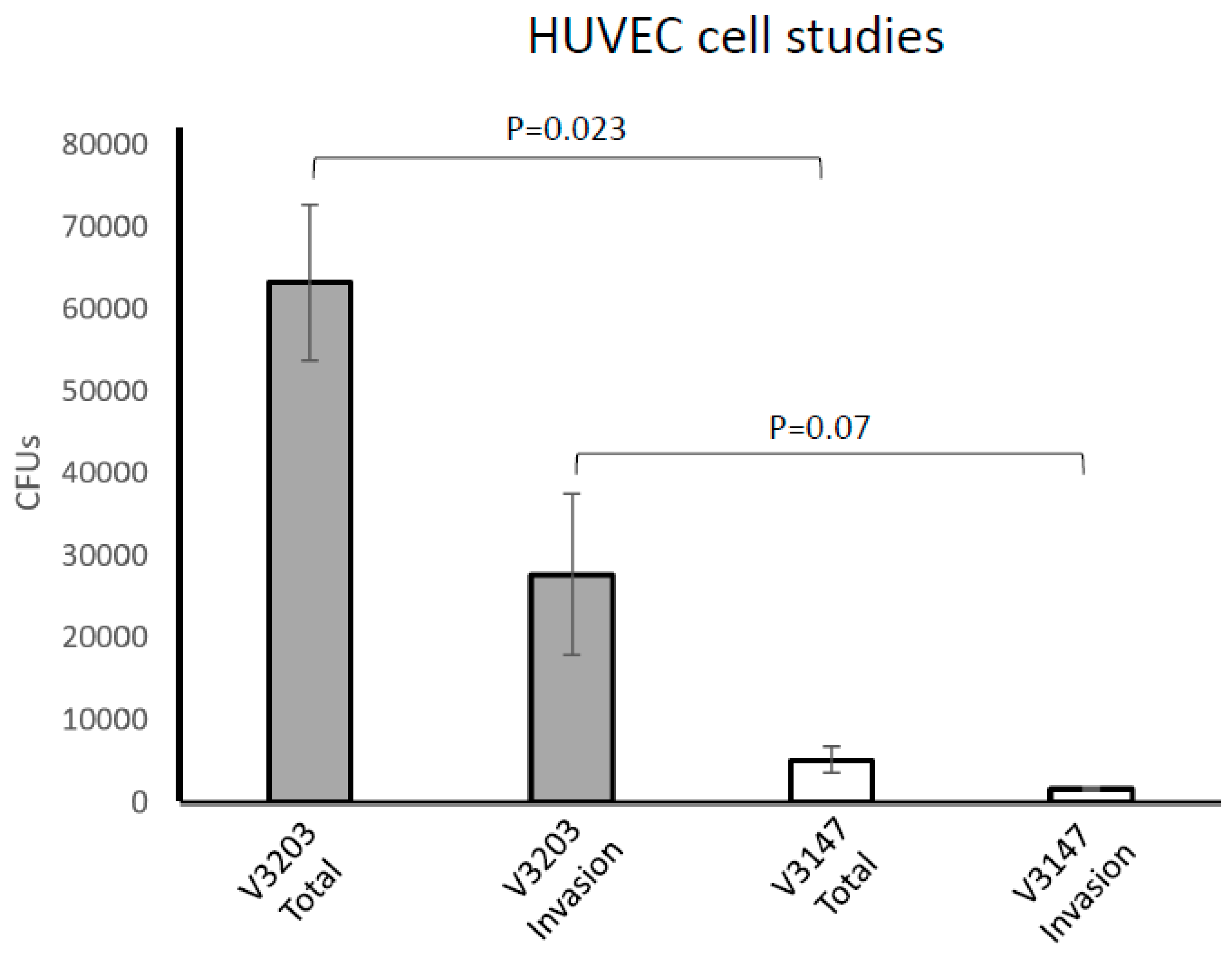
3.5. OxyR Is Required for P. intermedia Survival with Host Cells
4. Discussion
RNAseq Accession Number
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gui, Q.; Ramsey, K.; Hoffman, P.S.; Lewis, J.P. Amixicile depletes the ex vivo periodontal microbiome of anaerobic bacteria. J. Oral Biosci. 2020, 62, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Gui, Q.; Hoffman, P.S.; Lewis, J.P. Amixicile targets anaerobic bacteria within the oral microbiome. J. Oral Biosci. 2019, 61, 226–235. [Google Scholar] [CrossRef]
- López, N.J. Occurrence of Actinobacillus actinomycetemcomitans, Porphyromonas gingivalis, and Prevotella intermedia in progressive adult periodontitis. J. Periodontol. 2000, 71, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Lo Bue, A.M.L.; Nicoletti, G.; Toscano, A.M.; Rossetti, B.; Calì, G.; Condorelli, F. Porphyromonas gingivalis prevalence related to other micro-organisms in adult refractory periodontitis. New Microbiol. 1999, 22, 209–218. [Google Scholar] [PubMed]
- Teanpaisan, R.; Douglas, C.W.; Walsh, T.F. Characterisation of black-pigmented anaerobes isolated from diseased and healthy periodontal sites. J. Periodont. Res. 1995, 30, 245–251. [Google Scholar] [CrossRef]
- Maeda, N.; Okamoto, M.; Kondo, K.; Ishikawa, H.; Osada, R.; Tsurumoto, A.; Fujita, H. Incidence of Prevotella intermedia and Prevotella nigrescens in periodontal health and disease. Microbiol. Immunol. 1998, 42, 583–589. [Google Scholar] [CrossRef]
- Baumgartner, J.C.; Watkins, B.J.; Bae, K.S.; Xia, T. Association of black-pigmented bacteria with endodontic infections. J. Endod. 1999, 25, 413–415. [Google Scholar] [CrossRef]
- Falkler, W.A.; Enwonwu, C.O.; Idigbe, E.O. Microbiological understandings and mysteries of noma (cancrum oris). Oral Dis. 1999, 5, 150–155. [Google Scholar] [CrossRef]
- Enwonwu, C.O.; Falkler, W.A.; Idigbe, E.O. Oro-facial gangrene (noma/cancrum oris): Pathogenetic mechanisms. Crit. Rev. Oral Biol. Med. 2000, 11, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Usin, M.M.; Menso, J.; Rodríguez, V.I.; Gonzalez, A.; Tabares, S.; Parodi, R.; Sembaj, A. Association between maternal periodontitis and preterm and/or low birth weight infants in normal pregnancies. J. Matern. Fetal Neonatal. Med. 2016, 29, 115–119. [Google Scholar] [CrossRef]
- Haraszthy, V.I.; Zambon, J.J.; Trevisan, M.; Zeid, M.; Genco, R.J. Identification of periodontal pathogens in atheromatous plaques. J. Periodontol. 2000, 71, 1554–1560. [Google Scholar] [CrossRef]
- Madianos, P.; Lieff, S.; Murtha, A.; Boggess, K.; Auten, R.; Beck, J.; Offenbacher, S. Maternal periodontitis and prematurity. Part II: Maternal infection and fetal exposure. Ann. Periodontol. 2001, 6, 175–182. [Google Scholar] [CrossRef]
- Nagaoka, K.; Yanagihara, K.; Morinaga, Y.; Nakamura, S.; Harada, T.; Hasegawa, H.; Izumikawa, K.; Ishimatsu, Y.; Kakeya, H.; Nishimura, M.; et al. Prevotella intermedia induces severe bacteremic pneumococcal pneumonia in mice with upregulated platelet-activating factor receptor expression. Infect. Immun. 2014, 82, 587–593. [Google Scholar] [CrossRef] [Green Version]
- Mombelli, A.; Schmid, B.; Rutar, A.; Lang, N.P. Persistence patterns of Porphyromonas gingivalis, Prevotella intermedia/nigrescens, and Actinobacillus actinomyetemcomitans after mechanical therapy of periodontal disease. J. Periodontol. 2000, 71, 14–21. [Google Scholar] [CrossRef]
- Imlay, J.A. Where in the world do bacteria experience oxidative stress? Environ. Microbiol. 2019, 21, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Anaya, C.; Lewis, J.P. Outer membrane proteome of Prevotella intermedia 17: Identification of thioredoxin and iron-repressible hemin uptake loci. Proteomics 2007, 7, 403–412. [Google Scholar] [CrossRef]
- Lewis, J.P.; Plata, K.; Yu, F.; Rosato, A.; Anaya, C. Transcriptional organization, regulation and role of the Porphyromonas gingivalis W83 hmu haemin-uptake locus. Microbiology 2006, 152, 3367–3382. [Google Scholar] [CrossRef] [Green Version]
- Olczak, T.; Sroka, A.; Potempa, J.; Olczak, M. Porphyromonas gingivalis HmuY and HmuR: Further characterization of a novel mechanism of heme utilization. Arch. Microbiol. 2008, 189, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Bielecki, M.; Antonyuk, S.; Strange, R.W.; Siemińska, K.; Smalley, J.W.; Mackiewicz, P.; Śmiga, M.; Cowan, M.; Capper, M.J.; Ślęzak, P.; et al. Prevotella intermedia produces two proteins homologous to Porphyromonas gingivalis HmuY but with different heme coordination mode. Biochem. J. 2020, 477, 381–405. [Google Scholar] [CrossRef] [PubMed]
- Storz, G.; Tartaglia, L.A.; Ames, B.N. The OxyR regulon. Antonie Van Leeuwenhoek 1990, 58, 157–161. [Google Scholar] [CrossRef]
- Toledano, M.B.; Kullik, I.; Trinh, F.; Baird, P.T.; Schneider, T.D.; Storz, G. Redox-dependent shift of OxyR-DNA contacts along an extended DNA-binding site: A mechanism for differential promoter selection. Cell 1994, 78, 897–909. [Google Scholar] [CrossRef]
- Rocha, E.R.; Herren, C.D.; Smalley, D.J.; Smith, C.J. The complex oxidative stress response of Bacteroides fragilis: The role of OxyR in control of gene expression. Anaerobe 2003, 9, 165–173. [Google Scholar] [CrossRef]
- Tseng, H.-J.; McEwan, A.G.; Apicella, M.A.; Jennings, M.P. OxyR acts as a repressor of catalase expression in Neisseria gonorrhoeae. Infect. Immun. 2003, 71, 550–556. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-S.; Holmes, R.K. Characterization of OxyR as a negative transcriptional regulator that represses catalase production in Corynebacterium diphtheriae. PLoS ONE 2012, 7, e31709. [Google Scholar] [CrossRef] [Green Version]
- Si, M.; Zhao, C.; Burkinshaw, B.; Zhang, B.; Wei, D.; Wang, Y.; Dong, T.G.; Shen, X. Manganese scavenging and oxidative stress response mediated by type VI secretion system in Burkholderia thailandensis. Proc. Natl. Acad. Sci. USA 2017, 114, E2233–E2242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svintradze, D.V.; Peterson, D.L.; Collazo-Santiago, E.A.; Lewis, J.P.; Wright, H.T. Structures of the Porphyromonas gingivalis OxyR regulatory domain explain differences in expression of the OxyR regulon in Escherichia coli and P. gingivalis. Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 2091–2103. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Imlay, J.A. An anaerobic bacterium, Bacteroides thetaiotaomicron, uses a consortium of enzymes to scavenge hydrogen peroxide. Mol. Microbiol. 2013, 90, 1356–1371. [Google Scholar] [CrossRef] [Green Version]
- Honma, K.; Mishima, E.; Inagaki, S.; Sharma, A. The OxyR homologue in Tannerella forsythia regulates expression of oxidative stress responses and biofilm formation. Microbiology 2009, 155, 1912–1922. [Google Scholar] [CrossRef] [Green Version]
- Diaz, P.I.; Slakeski, N.; Reynolds, E.C.; Morona, R.; Rogers, A.H.; Kolenbrander, P.E. Role of oxyR in the oral anaerobe Porphyromonas gingivalis. J. Bacteriol. 2006, 188, 2454–2462. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.P.; Dawson, J.A.; Hannis, J.C.; Muddiman, D.; Macrina, F.L. Hemoglobinase activity of the lysine gingipain protease (Kgp) of Porphyromonas gingivalis W83. J. Bacteriol. 1999, 181, 4905–4913. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Ratnayake, D.B.; Okamoto, K.; Abe, N.; Yamamoto, K.; Nakayama, K. Genetic analyses of proteolysis, hemoglobin binding, and hemagglutination of Porphyromonas gingivalis. Construction of mutants with a combination of rgpA, rgpB, kgp, and hagA. J. Biol. Chem. 1999, 274, 17955–17960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueshima, J.; Shoji, M.; Ratnayake, D.B.; Abe, K.; Yoshida, S.; Yamamoto, K.; Nakayama, K. Purification, gene cloning, gene expression, and mutants of Dps from the obligate anaerobe Porphyromonas gingivalis. Infect. Immun. 2003, 71, 1170–1178. [Google Scholar] [CrossRef] [Green Version]
- Ohara, N.; Kikuchi, Y.; Shoji, M.; Naito, M.; Nakayama, K. Superoxide dismutase-encoding gene of the obligate anaerobe Porphyromonas gingivalis is regulated by the redox-sensing transcription activator OxyR. Microbiology 2006, 152, 955–966. [Google Scholar] [CrossRef] [Green Version]
- Naito, M.; Ogura, Y.; Itoh, T.; Shoji, M.; Okamoto, M.; Hayashi, T.; Nakayama, K. The complete genome sequencing of Prevotella intermedia strain OMA14 and a subsequent fine-scale, intra-species genomic comparison reveal an unusual amplification of conjugative and mobile transposons and identify a novel Prevotella-lineage-specific repeat. DNA Res. 2016, 23, 11–19. [Google Scholar]
- Yoshimura, M.; Ohara, N.; Kondo, Y.; Shoji, M.; Okano, S.; Nakano, Y.; Abiko, Y.; Nakayama, K. Proteome analysis of Porphyromonas gingivalis cells placed in a subcutaneous chamber of mice. Oral Microbiol. Immunol. 2008, 23, 413–418. [Google Scholar] [CrossRef]
- Nishikawa, K.; Duncan, M.J. Histidine kinase-mediated production and autoassembly of Porphyromonas gingivalis fimbriae. J. Bacteriol. 2010, 192, 1975–1987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanamandra, S.S.; Sarrafee, S.S.; Anaya-Bergman, C.; Jones, K.; Lewis, J.P. Role of the Porphyromonas gingivalis extracytoplasmic function sigma factor, SigH. Mol. Oral Microbiol. 2012, 27, 202–219. [Google Scholar] [CrossRef] [PubMed]
- Belvin, B.R.; Gui, Q.; Hutcherson, J.A.; Lewis, J.P. The Porphyromonas gingivalis Hybrid Cluster Protein Hcp Is Required for Growth with Nitrite and Survival with Host Cells. Infect. Immun. 2019, 87, e00572-18. [Google Scholar] [CrossRef] [Green Version]
- Shoemaker, N.B.; Guthrie, E.P.; Salyers, A.A.; Gardner, J.F. Evidence that the clindamycin-erythromycin resistance gene of Bacteroides plasmid pBF4 is on a transposable element. J. Bacteriol. 1985, 162, 626–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Miyazaki, H.; Anaya, C.; Yu, F.; Yeudall, W.A.; Lewis, J.P. Role of Porphyromonas gingivalis FeoB2 in metal uptake and oxidative stress protection. Infect. Immun. 2006, 74, 4214–4223. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, J.L.; Odelson, D.A.; Macrina, F.L. Complete nucleotide sequence and transcription of ermF, a macrolide-lincosamide-streptogramin B resistance determinant from Bacteroides fragilis. J. Bacteriol. 1986, 168, 523–533. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.D.; Chen, R.; Lenton, P.A.; Zhang, G.; Hinrichs, J.E.; Rudney, J.D. Persistence of extracrevicular bacterial reservoirs after treatment of aggressive periodontitis. J. Periodontol. 2008, 79, 2305–2312. [Google Scholar] [CrossRef]
- Wayakanon, K.; Thornhill, M.H.; Douglas, C.W.I.; Lewis, A.L.; Warren, N.J.; Pinnock, A.; Armes, S.P.; Battaglia, G.; Murdoch, C. Polymersome-mediated intracellular delivery of antibiotics to treat Porphyromonas gingivalis-infected oral epithelial cells. FASEB J. 2013, 27, 4455–4465. [Google Scholar] [CrossRef]
- Seaver, L.C.; Imlay, J.A. Alkyl hydroperoxide reductase is the primary scavenger of endogenous hydrogen peroxide in Escherichia coli. J. Bacteriol. 2001, 183, 7173–7181. [Google Scholar] [CrossRef] [Green Version]
- Poole, L.B. Flavin-dependent alkyl hydroperoxide reductase from Salmonella typhimurium. 2. Cystine disulfides involved in catalysis of peroxide reduction. Biochemistry 1996, 35, 65–75. [Google Scholar] [CrossRef]
- Poole, L.B.; Ellis, H.R. Flavin-dependent alkyl hydroperoxide reductase from Salmonella typhimurium. 1. Purification and enzymatic activities of overexpressed AhpF and AhpC proteins. Biochemistry 1996, 35, 56–64. [Google Scholar] [CrossRef]
- Rocha, E.R.; Owens, G.; Smith, C.J. The redox-sensitive transcriptional activator OxyR regulates the peroxide response regulon in the obligate anaerobe Bacteroides fragilis. J. Bacteriol. 2000, 182, 5059–5069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khademian, M.; Imlay, J.A. Escherichia coli cytochrome c peroxidase is a respiratory oxidase that enables the use of hydrogen peroxide as a terminal electron acceptor. Proc. Natl. Acad. Sci. USA 2017, 114, E6922–E6931. [Google Scholar] [CrossRef] [Green Version]
- Storz, G.; Altuvia, S. OxyR regulon. Methods Enzymol. 1994, 234, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Altuvia, S.; Storz, G. The novel oxyS RNA regulates expression of the sigma s subunit of Escherichia coli RNA polymerase. Nucleic Acids Symp. Ser. 1997, 36, 27–28. [Google Scholar]
- Zheng, M.; Wang, X.; Templeton, L.J.; Smulski, D.R.; LaRossa, R.A.; Storz, G. DNA microarray-mediated transcriptional profiling of the Escherichia coli response to hydrogen peroxide. J. Bacteriol. 2001, 183, 4562–4570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Gu, H.; Yang, Y.; Bai, H.; Zhao, C.; Si, M.; Su, T.; Shen, X. Molecular Mechanisms of AhpC in Resistance to Oxidative Stress in Burkholderia thailandensis. Front. Microbiol. 2019, 10, 1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimoto, H.; Takahasi, Y.; Kato, D.; Umemoto, T. Construction of a plasmid vector for transformation of Porphyromonas gingivalis. FEMS Microbiol. Lett. 1997, 152, 175–181. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naito, M.; Belvin, B.R.; Shoji, M.; Gui, Q.; Lewis, J.P. Insertional Inactivation of Prevotella intermedia OxyR Results in Reduced Survival with Oxidative Stress and in the Presence of Host Cells. Microorganisms 2021, 9, 551. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030551
Naito M, Belvin BR, Shoji M, Gui Q, Lewis JP. Insertional Inactivation of Prevotella intermedia OxyR Results in Reduced Survival with Oxidative Stress and in the Presence of Host Cells. Microorganisms. 2021; 9(3):551. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030551
Chicago/Turabian StyleNaito, Mariko, B. Ross Belvin, Mikio Shoji, Qin Gui, and Janina P. Lewis. 2021. "Insertional Inactivation of Prevotella intermedia OxyR Results in Reduced Survival with Oxidative Stress and in the Presence of Host Cells" Microorganisms 9, no. 3: 551. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030551