
Present and Future Perspectives on Therapeutic Options for Carbapenemase-Producing Enterobacterales Infections

 ,
,  , and
, and
Abstract
:1. Introduction
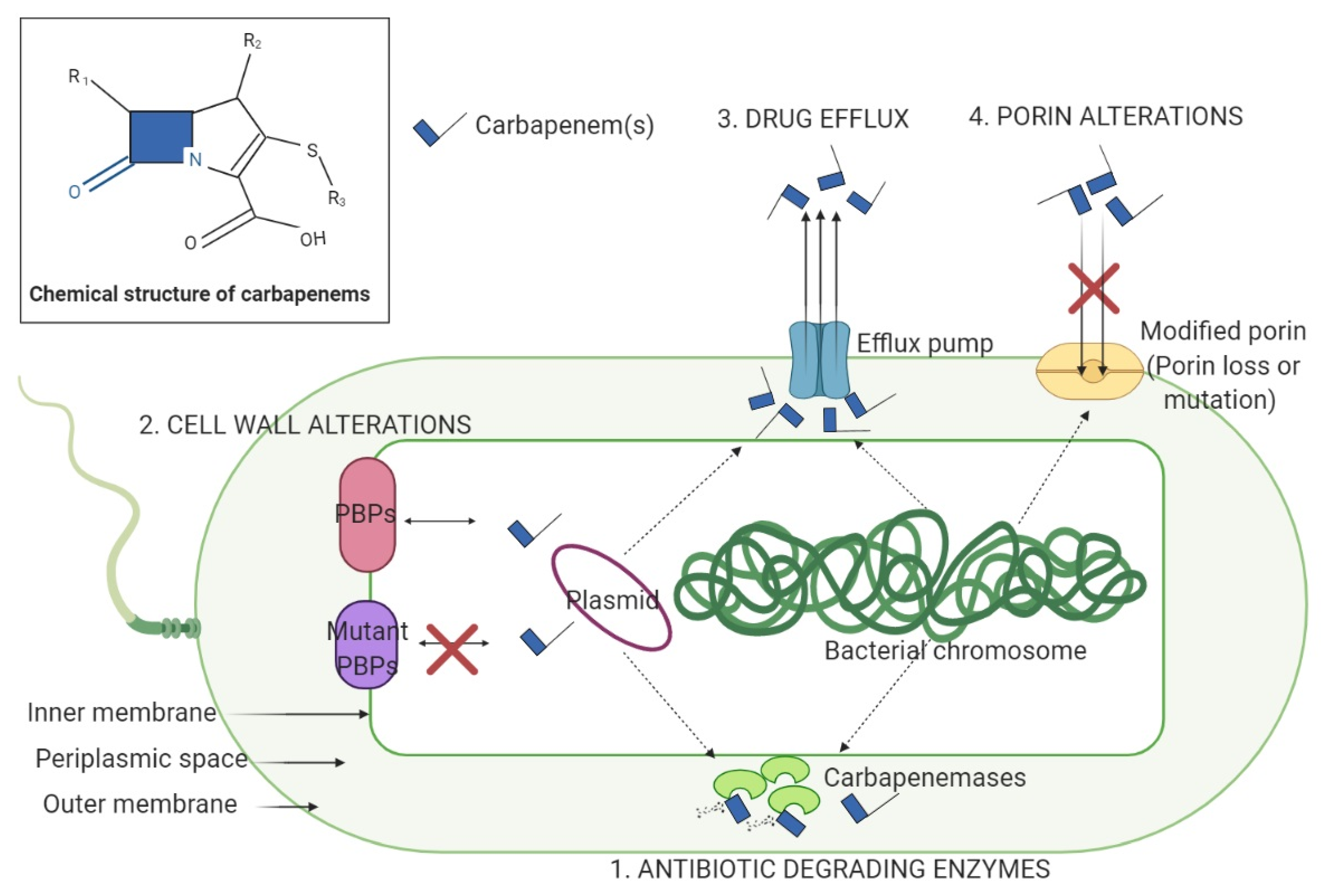
2. CRE Permeability and the Impact of Porin Deletions and Efflux Pumps
3. Old Antibiotics in Treating CRE Infections
4. Current Promising Antibiotics in Treating CRE Infections
5. Future Promising Strategies in CRE Treatment
6. Discussion
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wolrd Health Organization. WHO Priority Pathogens List for R&D of New Antibiotics; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Lee, C.-M.; Lai, C.-C.; Chiang, H.-T.; Lu, M.-C.; Wang, L.-F.; Tsai, T.-L.; Kang, M.-Y.; Jan, Y.-N.; Lo, Y.-T.; Ko, W.-C.; et al. Presence of multidrug-resistant organisms in the residents and environments of long-term care facilities in Taiwan. J. Microbiol. Immunol. Infect. 2017, 50, 133–144. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, R.G.; Johnson, J.K.; Bork, J.T.; Heil, E.L. Treatment options for extended-spectrum beta-lactamase (ESBL) and AmpC-producing bacteria. Expert Opin. Pharmacother. 2016, 17, 953–967. [Google Scholar] [CrossRef]
- Ambler, R.P. The structure of beta-lactamases. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1980, 289, 321–331. [Google Scholar] [CrossRef]
- Yigit, H.; Queenan, A.M.; Anderson, G.J.; Domenech-Sanchez, A.; Biddle, J.W.; Steward, C.D.; Alberti, S.; Bush, K.; Tenover, F.C. Novel carbapenem-hydrolyzing beta-lactamase, KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2001, 45, 1151–1161. [Google Scholar] [CrossRef] [Green Version]
- Codjoe, F.S.; Donkor, E.S. Carbapenem Resistance: A Review. Med. Sci. 2017, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Logan, L.K.; Weinstein, R.A. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J. Infect. Dis. 2017, 215, S28–S36. [Google Scholar] [CrossRef] [Green Version]
- Tzouvelekis, L.S.; Markogiannakis, A.; Psichogiou, M.; Tassios, P.T.; Daikos, G.L. Carbapenemases in Klebsiella pneumoniae and other Enterobacteriaceae: An evolving crisis of global dimensions. Clin. Microbiol. Rev. 2012, 25, 682–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention. National Action Plan for Combating Antibiotic-resistant Bacteria; CDC: Atlanta, GA, USA, 2015.
- Infectious Disease Society of America. The 10 × ’20 Initiative: Pursuing a global commitment to develop 10 new antibacterial drugs by 2020. Clin. Infect. Dis. 2010, 50, 1081–1083. [Google Scholar] [CrossRef] [Green Version]
- Nikaido, H.; Pagès, J.-M. Broad-specificity efflux pumps and their role in multidrug resistance of Gram-negative bacteria. FEMS Microbiol. Rev. 2012, 36, 340–363. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Réfregiers, M.; Pos, K.M.; Pagès, J.-M. Mechanisms of envelope permeability and antibiotic influx and efflux in Gram-negative bacteria. Nat. Microbiol. 2017, 2, 17001. [Google Scholar] [CrossRef] [PubMed]
- Vergalli, J.; Bodrenko, I.V.; Masi, M.; Moynié, L.; Acosta-Gutiérrez, S.; Naismith, J.H.; Davin-Regli, A.; Ceccarelli, M.; van den Berg, B.; Winterhalter, M.; et al. Porins and small-molecule translocation across the outer membrane of Gram-negative bacteria. Nat. Rev. Microbiol. 2020, 18, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Hong, Y.-K.; Lee, H.; Ko, K.S. High prevalence of non-clonal imipenem-nonsusceptible Enterobacter spp. isolates in Korea and their association with porin down-regulation. Diagn. Microbiol. Infect. Dis. 2017, 87, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Xu, L.; Han, Y.; Chen, Z.; Liu, C.; Ming, L. Mechanism for carbapenem resistance of clinical Enterobacteriaceae isolates. Exp. Ther. Med. 2018, 15, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Lomovskaya, O.; Sun, D.; Rubio-Aparicio, D.; Nelson, K.; Tsivkovski, R.; Griffith, D.C.; Dudley, M.N. Vaborbactam: Spectrum of Beta-Lactamase Inhibition and Impact of Resistance Mechanisms on Activity in Enterobacteriaceae. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, W.R.; Kline, E.G.; Jones, C.E.; Morder, K.T.; Mettus, R.T.; Doi, Y.; Nguyen, M.H.; Clancy, C.J.; Shields, R.K. Effects of KPC Variant and Porin Genotype on the In Vitro Activity of Meropenem-Vaborbactam against Carbapenem-Resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2019, 63, e02048-18. [Google Scholar] [CrossRef] [Green Version]
- Miao, M.; Wen, H.; Xu, P.; Niu, S.; Lv, J.; Xie, X.; Mediavilla, J.R.; Tang, Y.-W.; Kreiswirth, B.N.; Zhang, X.; et al. Genetic Diversity of Carbapenem-Resistant Enterobacteriaceae (CRE) Clinical Isolates from a Tertiary Hospital in Eastern China. Front. Microbiol. 2018, 9, 3341. [Google Scholar] [CrossRef] [Green Version]
- D’Onofrio, V.; Conzemius, R.; Varda-Brkić, D.; Bogdan, M.; Grisold, A.; Gyssens, I.C.; Bedenić, B.; Barišić, I. Epidemiology of colistin-resistant, carbapenemase-producing Enterobacteriaceae and Acinetobacter baumannii in Croatia. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2020, 81, 104263. [Google Scholar] [CrossRef] [PubMed]
- Nang, S.C.; Li, J.; Velkov, T. The rise and spread of mcr plasmid-mediated polymyxin resistance. Crit. Rev. Microbiol. 2019, 45, 131–161. [Google Scholar] [CrossRef] [PubMed]
- Macesic, N.; Nelson, B.; Mcconville, T.H.; Giddins, M.J.; Green, D.A.; Stump, S.; Gomez-Simmonds, A.; Annavajhala, M.K.; Uhlemann, A.-C. Emergence of Polymyxin Resistance in Clinical Klebsiella pneumoniae Through Diverse Genetic Adaptations: A Genomic, Retrospective Cohort Study. Clin. Infect. Dis. 2020, 70, 2084–2091. [Google Scholar] [CrossRef]
- De Araújo Longo, L.G.; Fontana, H.; Santos de Sousa, V.; Chilinque Zambão da Silva, N.; Souto Martins, I.; Meurer Moreira, B. Emergence of mgrB locus deletion mediating polymyxin resistance in pandemic KPC-producing Klebsiella pneumoniae ST15 lineage. J. Med. Microbiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Chen, Y.; Liu, D.; Yang, D.; Liu, Z.; Wang, Y.; Wang, J.; Wang, X.; Xu, X.; Li, X.; et al. Abundance of tigecycline resistance genes and association with antibiotic residues in Chinese livestock farms. J. Hazard. Mater. 2021, 409, 124921. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhu, Z.; Chen, Y.; Wang, W.; He, F. The Plasmid-Borne tet(A) Gene Is an Important Factor Causing Tigecycline Resistance in ST11 Carbapenem-Resistant Klebsiella pneumoniae Under Selective Pressure. Front. Microbiol. 2021, 12, 644949. [Google Scholar] [CrossRef] [PubMed]
- Ugah, U.I.; Udeani, T.K. High Prevalence of Phenotypic Resistance to Colistin, Tigecycline and Netilmicin in a Region with no History of Colistin Administration in Nigeria. Clin. Lab. 2021, 67. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Baño, J.; Gutiérrez-Gutiérrez, B.; Machuca, I.; Pascual, A. Treatment of Infections Caused by Extended-Spectrum-Beta-Lactamase-, AmpC-, and Carbapenemase-Producing Enterobacteriaceae. Clin. Microbiol. Rev. 2018, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falagas, M.E.; Karageorgopoulos, D.E.; Nordmann, P. Therapeutic options for infections with Enterobacteriaceae producing carbapenem-hydrolyzing enzymes. Future Microbiol. 2011, 6, 653–666. [Google Scholar] [CrossRef]
- Livermore, D.M.; Warner, M.; Mushtaq, S.; Doumith, M.; Zhang, J.; Woodford, N. What remains against carbapenem-resistant Enterobacteriaceae? Evaluation of chloramphenicol, ciprofloxacin, colistin, fosfomycin, minocycline, nitrofurantoin, temocillin and tigecycline. Int. J. Antimicrob. Agents 2011, 37, 415–419. [Google Scholar] [CrossRef] [Green Version]
- Karaiskos, I.; Lagou, S.; Pontikis, K.; Rapti, V.; Poulakou, G. The “Old” and the “New” Antibiotics for MDR Gram-Negative Pathogens: For Whom, When, and How. Front. Public Health 2019, 7, 151. [Google Scholar] [CrossRef] [Green Version]
- Falagas, M.E.; Rafailidis, P.I.; Kasiakou, S.K.; Hatzopoulou, P.; Michalopoulos, A. Effectiveness and nephrotoxicity of colistin monotherapy vs. colistin-meropenem combination therapy for multidrug-resistant Gram-negative bacterial infections. Clin. Microbiol. Infect. Dis. 2006, 12, 1227–1230. [Google Scholar] [CrossRef]
- Pogue, J.M.; Lee, J.; Marchaim, D.; Yee, V.; Zhao, J.J.; Chopra, T.; Lephart, P.; Kaye, K.S. Incidence of and risk factors for colistin-associated nephrotoxicity in a large academic health system. Clin. Infect. Dis. 2011, 53, 879–884. [Google Scholar] [CrossRef] [Green Version]
- Martis, N.; Leroy, S.; Blanc, V. Colistin in multi-drug resistant Pseudomonas aeruginosa blood-stream infections: A narrative review for the clinician. J. Infect. 2014, 69, 1–12. [Google Scholar] [CrossRef]
- Grégoire, N.; Aranzana-Climent, V.; Magréault, S.; Marchand, S.; Couet, W. Clinical Pharmacokinetics and Pharmacodynamics of Colistin. Clin. Pharmacokinet. 2017, 56, 1441–1460. [Google Scholar] [CrossRef]
- Kazmierczak, K.M.; de Jonge, B.L.M.; Stone, G.G.; Sahm, D.F. In vitro activity of ceftazidime/avibactam against isolates of Enterobacteriaceae collected in European countries: INFORM global surveillance 2012–15. J. Antimicrob. Chemother. 2018, 73, 2782–2788. [Google Scholar] [CrossRef] [PubMed]
- Karaiskos, I.; Souli, M.; Galani, I.; Giamarellou, H. Colistin: Still a lifesaver for the 21st century? Expert Opin. Drug Metab. Toxicol. 2017, 13, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Karaiskos, I.; Antoniadou, A.; Giamarellou, H. Combination therapy for extensively-drug resistant gram-negative bacteria. Expert Rev. Anti. Infect. Ther. 2017, 15, 1123–1140. [Google Scholar] [CrossRef] [PubMed]
- Daikos, G.L.; Tsaousi, S.; Tzouvelekis, L.S.; Anyfantis, I.; Psichogiou, M.; Argyropoulou, A.; Stefanou, I.; Sypsa, V.; Miriagou, V.; Nepka, M.; et al. Carbapenemase-producing Klebsiella pneumoniae bloodstream infections: Lowering mortality by antibiotic combination schemes and the role of carbapenems. Antimicrob. Agents Chemother. 2014, 58, 2322–2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, B.T.; Pogue, J.M.; Zavascki, A.P.; Paul, M.; Daikos, G.L.; Forrest, A.; Giacobbe, D.R.; Viscoli, C.; Giamarellou, H.; Karaiskos, I.; et al. International Consensus Guidelines for the Optimal Use of the Polymyxins: Endorsed by the American College of Clinical Pharmacy (ACCP), European Society of Clinical Microbiology and Infectious Diseases (ESCMID), Infectious Diseases Society of America (IDSA), International Society for Anti‐infective Pharmacology (ISAP), Society of Critical Care Medicine (SCCM), and Society of Infectious Diseases Pharmacists (SIDP). Pharmacotherapy 2019, 39, 10–39. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Izdebski, R.; Baraniak, A.; Bojarska, K.; Urbanowicz, P.; Fiett, J.; Pomorska-Wesołowska, M.; Hryniewicz, W.; Gniadkowski, M.; Żabicka, D. Mobile MCR-1-associated resistance to colistin in Poland. J. Antimicrob. Chemother. 2016, 71, 2331–2333. [Google Scholar] [CrossRef]
- Baron, S.; Hadjadj, L.; Rolain, J.-M.; Olaitan, A.O. Molecular mechanisms of polymyxin resistance: Knowns and unknowns. Int. J. Antimicrob. Agents 2016, 48, 583–591. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, D.; Quan, J.; Hua, X.; Yu, Y. mcr-1 facilitated selection of high-level colistin-resistant mutants in Escherichia coli. Clin. Microbiol. Infect. 2019, 25, 517.e1–517.e4. [Google Scholar] [CrossRef] [Green Version]
- Xavier, B.B.; Lammens, C.; Ruhal, R.; Kumar-Singh, S.; Butaye, P.; Goossens, H.; Malhotra-Kumar, S. Identification of a novel plasmid-mediated colistin-resistance gene, mcr-2, in Escherichia coli, Belgium, June 2016. Euro Surveill. 2016, 21, 30280. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Li, H.; Shen, Y.; Liu, Z.; Wang, S.; Shen, Z.; Zhang, R.; Walsh, T.R.; Shen, J.; Wang, Y. Novel Plasmid-Mediated Colistin Resistance Gene mcr-3 in Escherichia coli. MBio 2017, 8, e00543-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A.; Villa, L.; Feudi, C.; Curcio, L.; Orsini, S.; Luppi, A.; Pezzotti, G.; Magistrali, C.F. Novel plasmid-mediated colistin resistance mcr-4 gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. Euro Surveill. 2017, 22, 30589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borowiak, M.; Fischer, J.; Hammerl, J.A.; Hendriksen, R.S.; Szabo, I.; Malorny, B. Identification of a novel transposon-associated phosphoethanolamine transferase gene, mcr-5, conferring colistin resistance in d-tartrate fermenting Salmonella enterica subsp. enterica serovar Paratyphi B. J. Antimicrob. Chemother. 2017, 72, 3317–3324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of Novel Mobilized Colistin Resistance Gene mcr-9 in a Multidrug-Resistant, Colistin-Susceptible Salmonella enterica Serotype Typhimurium Isolate. MBio 2019, 10, e00853-19. [Google Scholar] [CrossRef] [Green Version]
- Vrancianu, C.O.; Popa, L.I.; Bleotu, C.; Chifiriuc, M.C. Targeting Plasmids to Limit Acquisition and Transmission of Antimicrobial Resistance. Front. Microbiol. 2020, 11, 761. [Google Scholar] [CrossRef]
- Berglund, B.; Hoang, N.T.B.; Tärnberg, M.; Le, N.K.; Svartström, O.; Khu, D.T.K.; Nilsson, M.; Le, H.T.; Welander, J.; Olson, L.; et al. Insertion sequence transpositions and point mutations in mgrB causing colistin resistance in a clinical strain of carbapenem-resistant Klebsiella pneumoniae from Vietnam. Int. J. Antimicrob. Agents 2018, 51, 789–793. [Google Scholar] [CrossRef]
- Uz Zaman, T.; Albladi, M.; Siddique, M.I.; Aljohani, S.M.; Balkhy, H.H. Insertion element mediated mgrB disruption and presence of ISKpn28 in colistin-resistant Klebsiella pneumoniae isolates from Saudi Arabia. Infect. Drug Resist. 2018, 11, 1183–1187. [Google Scholar] [CrossRef] [Green Version]
- Snyman, Y.; Whitelaw, A.C.; Reuter, S.; Maloba, M.R.B.; Newton-Foot, M. Colistin Resistance Mechanisms in Clinical Escherichia coli and Klebsiella spp. Isolates from the Western Cape of South Africa. Microb. Drug Resist. 2021. ahead of print. [Google Scholar] [CrossRef]
- He, Y.-Z.; Long, T.-F.; He, B.; Li, X.-P.; Li, G.; Chen, L.; Liao, X.-P.; Liu, Y.-H.; Sun, J. ISEc69-Mediated Mobilization of the Colistin Resistance Gene mcr-2 in Escherichia coli. Front. Microbiol. 2020, 11, 564973. [Google Scholar] [CrossRef]
- Falagas, M.E.; Vouloumanou, E.K.; Samonis, G.; Vardakas, K.Z. Fosfomycin. Clin. Microbiol. Rev. 2016, 29, 321–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontikis, K.; Karaiskos, I.; Bastani, S.; Dimopoulos, G.; Kalogirou, M.; Katsiari, M.; Oikonomou, A.; Poulakou, G.; Roilides, E.; Giamarellou, H. Outcomes of critically ill intensive care unit patients treated with fosfomycin for infections due to pandrug-resistant and extensively drug-resistant carbapenemase-producing Gram-negative bacteria. Int. J. Antimicrob. Agents 2014, 43, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Monden, K.; Ando, E.; Iida, M.; Kumon, H. Role of fosfomycin in a synergistic combination with ofloxacin against Pseudomonas aeruginosa growing in a biofilm. J. Infect. Chemother. 2002, 8, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Steinort, D.; Pillai, S.; Joukhadar, C. Fosfomycin: An old, new friend? Clin. Microbiol. Infect. Dis. 2010, 29, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Michalopoulos, A.S.; Livaditis, I.G.; Gougoutas, V. The revival of fosfomycin. Int. J. Infect. Dis. 2011, 15, e732–e739. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Candel, F.J.; Cantón, R. Current approach to fosfomycin: From bench to bedside. Enferm. Infecc. Microbiol. Clin. 2019, 37, 1–3. [Google Scholar] [CrossRef]
- Rodríguez-Baño, J.; Cisneros, J.M.; Cobos-Trigueros, N.; Fresco, G.; Navarro-San Francisco, C.; Gudiol, C.; Horcajada, J.P.; López-Cerero, L.; Martínez, J.A.; Molina, J.; et al. Executive summary of the diagnosis and antimicrobial treatment of invasive infections due to multidrug-resistant Enterobacteriaceae. Guidelines of the Spanish Society of Infectious Diseases and Clinical Microbiology (SEIMC). Enferm. Infecc. Microbiol. Clin. 2015, 33, 338–341. [Google Scholar] [CrossRef]
- López-Cerero, L.; de Cueto, M.; Díaz-Guerrero, M.A.; Morillo, C.; Pascual, A. Evaluation of the Etest method for fosfomycin susceptibility of ESBL-producing Klebsiella pneumoniae. J. Antimicrob. Chemother. 2007, 59, 810–812. [Google Scholar] [CrossRef] [Green Version]
- De Cueto, M.; López, L.; Hernández, J.R.; Morillo, C.; Pascual, A. In vitro activity of fosfomycin against extended-spectrum-beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae: Comparison of susceptibility testing procedures. Antimicrob. Agents Chemother. 2006, 50, 368–370. [Google Scholar] [CrossRef] [Green Version]
- Kaase, M.; Szabados, F.; Anders, A.; Gatermann, S.G. Fosfomycin susceptibility in carbapenem-resistant Enterobacteriaceae from Germany. J. Clin. Microbiol. 2014, 52, 1893–1897. [Google Scholar] [CrossRef] [Green Version]
- Ballestero-Téllez, M.; Docobo-Pérez, F.; Rodríguez-Martínez, J.M.; Conejo, M.C.; Ramos-Guelfo, M.S.; Blázquez, J.; Rodríguez-Baño, J.; Pascual, A. Role of inoculum and mutant frequency on fosfomycin MIC discrepancies by agar dilution and broth microdilution methods in Enterobacteriaceae. Clin. Microbiol. Infect. 2017, 23, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Aprile, A.; Scalia, G.; Stefani, S.; Mezzatesta, M.L. In vitro fosfomycin study on concordance of susceptibility testing methods against ESBL and carbapenem-resistant Enterobacteriaceae. J. Glob. Antimicrob. Resist. 2020, 23, 286–289. [Google Scholar] [CrossRef]
- Campanile, F.; Wootton, M.; Davies, L.; Aprile, A.; Mirabile, A.; Pomponio, S.; Demetrio, F.; Bongiorno, D.; Walsh, T.R.; Stefani, S.; et al. Gold standard susceptibility testing of fosfomycin in Staphylococcus aureus and Enterobacterales using a new agar dilution panel®. J. Glob. Antimicrob. Resist. 2020, 23, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Cottell, J.L.; Webber, M.A. Experiences in fosfomycin susceptibility testing and resistance mechanism determination in Escherichia coli from urinary tract infections in the UK. J. Med. Microbiol. 2019, 68, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.C.; Brigman, H.V.; Anderson, J.C.; Emery, C.L.; Bias, T.E.; Bergen, P.J.; Landersdorfer, C.B.; Hirsch, E.B. Performance of Four Fosfomycin Susceptibility Testing Methods against an International Collection of Clinical Pseudomonas aeruginosa Isolates. J. Clin. Microbiol. 2020, 58, e01121-20. [Google Scholar] [CrossRef] [PubMed]
- Mojica, M.F.; De La Cadena, E.; Hernández-Gómez, C.; Correa, A.; Appel, T.M.; Pallares, C.J.; Villegas, M.V. Performance of disk diffusion and broth microdilution for fosfomycin susceptibility testing of multidrug-resistant clinical isolates of Enterobacterales and Pseudomonas aeruginosa. J. Glob. Antimicrob. Resist. 2020, 21, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Albur, M.S.; Noel, A.; Bowker, K.; MacGowan, A. The combination of colistin and fosfomycin is synergistic against NDM-1-producing Enterobacteriaceae in in vitro pharmacokinetic/pharmacodynamic model experiments. Int. J. Antimicrob. Agents 2015, 46, 560–567. [Google Scholar] [CrossRef]
- Vardakas, K.Z.; Legakis, N.J.; Triarides, N.; Falagas, M.E. Susceptibility of contemporary isolates to fosfomycin: A systematic review of the literature. Int. J. Antimicrob. Agents 2016, 47, 269–285. [Google Scholar] [CrossRef] [PubMed]
- Galani, I.; Karaiskos, I.; Karantani, I.; Papoutsaki, V.; Maraki, S.; Papaioannou, V.; Kazila, P.; Tsorlini, H.; Charalampaki, N.; Toutouza, M.; et al. Epidemiology and resistance phenotypes of carbapenemase-producing Klebsiella pneumoniae in Greece, 2014 to 2016. Euro Surveill. 2018, 23, 1700775. [Google Scholar] [CrossRef] [Green Version]
- Flamm, R.K.; Rhomberg, P.R.; Watters, A.A.; Sweeney, K.; Ellis-Grosse, E.J.; Shortridge, D. Activity of fosfomycin when tested against US contemporary bacterial isolates. Diagn. Microbiol. Infect. Dis. 2019, 93, 143–146. [Google Scholar] [CrossRef]
- Dinh, A.; Salomon, J.; Bru, J.P.; Bernard, L. Fosfomycin: Efficacy against infections caused by multidrug-resistant bacteria. Scand. J. Infect. Dis. 2012, 44, 182–189. [Google Scholar] [CrossRef]
- Bassetti, M.; Giacobbe, D.R.; Giamarellou, H.; Viscoli, C.; Daikos, G.L.; Dimopoulos, G.; De Rosa, F.G.; Giamarellos-Bourboulis, E.J.; Rossolini, G.M.; Righi, E.; et al. Management of KPC-producing Klebsiella pneumoniae infections. Clin. Microbiol. Infect. 2018, 24, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Karageorgopoulos, D.E.; Miriagou, V.; Tzouvelekis, L.S.; Spyridopoulou, K.; Daikos, G.L. Emergence of resistance to fosfomycin used as adjunct therapy in KPC Klebsiella pneumoniae bacteraemia: Report of three cases. J. Antimicrob. Chemother. 2012, 67, 2777–2779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, W.; Luo, Q.; Shi, Q.; Huang, C.; Yu, X.; Niu, T.; Zhou, K.; Zhang, J.; Xiao, Y. In vitro antibacterial effect of fosfomycin combination therapy against colistin-resistant Klebsiella pneumoniae. Infect. Drug Resist. 2018, 11, 577–585. [Google Scholar] [CrossRef] [Green Version]
- Kollef, M.H.; Ricard, J.-D.; Roux, D.; Francois, B.; Ischaki, E.; Rozgonyi, Z.; Boulain, T.; Ivanyi, Z.; János, G.; Garot, D.; et al. A Randomized Trial of the Amikacin Fosfomycin Inhalation System for the Adjunctive Therapy of Gram-Negative Ventilator-Associated Pneumonia: IASIS Trial. Chest 2017, 151, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Perdigão Neto, L.V.; Oliveira, M.S.; Martins, R.C.R.; Marchi, A.P.; Gaudereto, J.J.; da Costa, L.A.T.J.; de Lima, L.F.A.; Takeda, C.F.V.; Costa, S.F.; Levin, A.S. Fosfomycin in severe infections due to genetically distinct pan-drug-resistant Gram-negative microorganisms: Synergy with meropenem. J. Antimicrob. Chemother. 2019, 74, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Garnacho-Montero, J.; Dimopoulos, G.; Poulakou, G.; Akova, M.; Cisneros, J.M.; De Waele, J.; Petrosillo, N.; Seifert, H.; Timsit, J.F.; Vila, J.; et al. Task force on management and prevention of Acinetobacter baumannii infections in the ICU. Intensive Care Med. 2015, 41, 2057–2075. [Google Scholar] [CrossRef]
- Giamarellou, H.; Poulakou, G. Pharmacokinetic and pharmacodynamic evaluation of tigecycline. Expert Opin. Drug Metab. Toxicol. 2011, 7, 1459–1470. [Google Scholar] [CrossRef] [PubMed]
- Koomanachai, P.; Kim, A.; Nicolau, D.P. Pharmacodynamic evaluation of tigecycline against Acinetobacter baumannii in a murine pneumonia model. J. Antimicrob. Chemother. 2009, 63, 982–987. [Google Scholar] [CrossRef] [Green Version]
- Routsi, C.; Kokkoris, S.; Douka, E.; Ekonomidou, F.; Karaiskos, I.; Giamarellou, H. High-dose tigecycline-associated alterations in coagulation parameters in critically ill patients with severe infections. Int. J. Antimicrob. Agents 2015, 45, 90–93. [Google Scholar] [CrossRef]
- Falagas, M.E.; Vardakas, K.Z.; Tsiveriotis, K.P.; Triarides, N.A.; Tansarli, G.S. Effectiveness and safety of high-dose tigecycline-containing regimens for the treatment of severe bacterial infections. Int. J. Antimicrob. Agents 2014, 44, 1–7. [Google Scholar] [CrossRef]
- Karakonstantis, S. A systematic review of implications, mechanisms, and stability of in vivo emergent resistance to colistin and tigecycline in Acinetobacter baumannii. J. Chemother. 2021, 33, 1–11. [Google Scholar] [CrossRef]
- Zavascki, A.P.; Klee, B.O.; Bulitta, J.B. Aminoglycosides against carbapenem-resistant Enterobacteriaceae in the critically ill: The pitfalls of aminoglycoside susceptibility. Exp. Rev. Anti. Infect. Ther. 2017, 15, 519–526. [Google Scholar] [CrossRef]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Update 2010, 13, 151–171. [Google Scholar] [CrossRef] [Green Version]
- Tada, T.; Miyoshi-Akiyama, T.; Kato, Y.; Ohmagari, N.; Takeshita, N.; Hung, N.V.; Phuong, D.M.; Thu, T.A.; Binh, N.G.; Anh, N.Q.; et al. Emergence of 16S rRNA methylase-producing Acinetobacter baumannii and Pseudomonas aeruginosa isolates in hospitals in Vietnam. BMC Infect. Dis. 2013, 13, 251. [Google Scholar] [CrossRef] [Green Version]
- Heidary, M.; Salimi Chirani, A.; Khoshnood, S.; Eslami, G.; Atyabi, S.M.; Nazem, H.; Fazilati, M.; Hashemi, A.; Soleimani, S. Molecular detection of aminoglycoside-modifying enzyme genes in Acinetobacter baumannii clinical isolates. Acta Microbiol. Immunol. Hung. 2017, 64, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Sheikhalizadeh, V.; Hasani, A.; Ahangarzadeh Rezaee, M.; Rahmati-Yamchi, M.; Hasani, A.; Ghotaslou, R.; Goli, H.R. Comprehensive study to investigate the role of various aminoglycoside resistance mechanisms in clinical isolates of Acinetobacter baumannii. J. Infect. Chemother. 2017, 23, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Garneau-Tsodikova, S.; Labby, K.J. Mechanisms of Resistance to Aminoglycoside Antibiotics: Overview and Perspectives. Medchemcomm 2016, 7, 11–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doi, Y.; Arakawa, Y. 16S ribosomal RNA methylation: Emerging resistance mechanism against aminoglycosides. Clin. Infect. Dis. 2007, 45, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Berçot, B.; Poirel, L.; Nordmann, P. Updated multiplex polymerase chain reaction for detection of 16S rRNA methylases: High prevalence among NDM-1 producers. Diagn. Microbiol. Infect. Dis. 2011, 71, 442–445. [Google Scholar] [CrossRef]
- Zhou, Y.; Yu, H.; Guo, Q.; Xu, X.; Ye, X.; Wu, S.; Guo, Y.; Wang, M. Distribution of 16S rRNA methylases among different species of Gram-negative bacilli with high-level resistance to aminoglycosides. Clin. Microbiol. Infect. Dis. 2010, 29, 1349–1353. [Google Scholar] [CrossRef] [PubMed]
- Alexander, B.T.; Marschall, J.; Tibbetts, R.J.; Neuner, E.A.; Dunne, W.M.J.; Ritchie, D.J. Treatment and clinical outcomes of urinary tract infections caused by KPC-producing Enterobacteriaceae in a retrospective cohort. Clin. Ther. 2012, 34, 1314–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roger, C.; Nucci, B.; Louart, B.; Friggeri, A.; Knani, H.; Evrard, A.; Lavigne, J.-P.; Allaouchiche, B.; Lefrant, J.-Y.; Roberts, J.A.; et al. Impact of 30 mg/kg amikacin and 8 mg/kg gentamicin on serum concentrations in critically ill patients with severe sepsis. J. Antimicrob. Chemother. 2016, 71, 208–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Duin, D.; Cober, E.; Richter, S.S.; Perez, F.; Kalayjian, R.C.; Salata, R.A.; Evans, S.; Fowler, V.G.J.; Kaye, K.S.; Bonomo, R.A. Impact of therapy and strain type on outcomes in urinary tract infections caused by carbapenem-resistant Klebsiella pneumoniae. J. Antimicrob. Chemother. 2015, 70, 1203–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paquette, F.; Bernier-Jean, A.; Brunette, V.; Ammann, H.; Lavergne, V.; Pichette, V.; Troyanov, S.; Bouchard, J. Acute Kidney Injury and Renal Recovery with the Use of Aminoglycosides: A Large Retrospective Study. Nephron 2015, 131, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.-S.; Gould, I.M.; Lee, W.-S.; Hsueh, P.-R. New Drugs for Multidrug-Resistant Gram-Negative Organisms: Time for Stewardship. Drugs 2019, 79, 705–714. [Google Scholar] [CrossRef]
- Wilson, G.M.; Fitzpatrick, M.; Walding, K.; Gonzalez, B.; Schweizer, M.L.; Suda, K.J.; Evans, C.T. Meta-analysis of Clinical Outcomes Using Ceftazidime/Avibactam, Ceftolozane/Tazobactam, and Meropenem/Vaborbactam for the Treatment of Multidrug-Resistant Gram-Negative Infections. Open Forum Infect. Dis. 2021, 8, ofaa651. [Google Scholar] [CrossRef]
- Karaiskos, I.; Galani, I.; Souli, M.; Giamarellou, H. Novel β-lactam-β-lactamase inhibitor combinations: Expectations for the treatment of carbapenem-resistant Gram-negative pathogens. Expert Opin. Drug Metab. Toxicol. 2019, 15, 133–149. [Google Scholar] [CrossRef]
- De Jonge, B.L.M.; Karlowsky, J.A.; Kazmierczak, K.M.; Biedenbach, D.J.; Sahm, D.F.; Nichols, W.W. In Vitro Susceptibility to Ceftazidime-Avibactam of Carbapenem-Nonsusceptible Enterobacteriaceae Isolates Collected during the INFORM Global Surveillance Study (2012 to 2014). Antimicrob. Agents Chemother. 2016, 60, 3163–3169. [Google Scholar] [CrossRef] [Green Version]
- Lahiri, S.D.; Johnstone, M.R.; Ross, P.L.; McLaughlin, R.E.; Olivier, N.B.; Alm, R.A. Avibactam and class C β-lactamases: Mechanism of inhibition, conservation of the binding pocket, and implications for resistance. Antimicrob. Agents Chemother. 2014, 58, 5704–5713. [Google Scholar] [CrossRef] [Green Version]
- Karlowsky, J.A.; Biedenbach, D.J.; Kazmierczak, K.M.; Stone, G.G.; Sahm, D.F. Activity of Ceftazidime-Avibactam against Extended-Spectrum- and AmpC β-Lactamase-Producing Enterobacteriaceae Collected in the INFORM Global Surveillance Study from 2012 to 2014. Antimicrob. Agents Chemother. 2016, 60, 2849–2857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shields, R.K.; Nguyen, M.H.; Chen, L.; Press, E.G.; Potoski, B.A.; Marini, R.V.; Doi, Y.; Kreiswirth, B.N.; Clancy, C.J. Ceftazidime-Avibactam Is Superior to Other Treatment Regimens against Carbapenem-Resistant Klebsiella pneumoniae Bacteremia. Antimicrob. Agents Chemother. 2017, 61, e00883-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Duin, D.; Lok, J.J.; Earley, M.; Cober, E.; Richter, S.S.; Perez, F.; Salata, R.A.; Kalayjian, R.C.; Watkins, R.R.; Doi, Y.; et al. Colistin Versus Ceftazidime-Avibactam in the Treatment of Infections Due to Carbapenem-Resistant Enterobacteriaceae. Clin. Infect. Dis. 2018, 66, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Van Asten, S.A.V.; Boattini, M.; Kraakman, M.E.M.; Bianco, G.; Iannaccone, M.; Costa, C.; Cavallo, R.; Bernards, A.T. Ceftazidime-avibactam resistance and restoration of carbapenem susceptibility in KPC-producing Klebsiella pneumoniae infections: A case series. J. Infect. Chemother. 2021, 27, 778–780. [Google Scholar] [CrossRef]
- Shapiro, A.B.; Moussa, S.H.; Carter, N.M.; Gao, N.; Miller, A.A. Ceftazidime–Avibactam Resistance Mutations V240G, D179Y, and D179Y/T243M in KPC-3 β-Lactamase Do Not Alter Cefpodoxime–ETX1317 Susceptibility. ACS Infect. Dis. 2021, 7, 79–87. [Google Scholar] [CrossRef]
- Pagès, J.-M.; Peslier, S.; Keating, T.A.; Lavigne, J.-P.; Nichols, W.W. Role of the Outer Membrane and Porins in Susceptibility of β-Lactamase-Producing Enterobacteriaceae to Ceftazidime-Avibactam. Antimicrob. Agents Chemother. 2015, 60, 1349–1359. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wu, T.; Perez, O.; Rana, A.P.; Chen, L.; Kreiswirth, B.N.; Satlin, M.J.; Bulman, Z.P. In vitro Optimization of Ceftazidime/Avibactam for KPC-Producing Klebsiella pneumoniae. Front. Microbiol. 2021, 12, 414. [Google Scholar] [CrossRef]
- Yin, D.; Wu, S.; Yang, Y.; Shi, Q.; Dong, D.; Zhu, D.; Hu, F. Results from the China Antimicrobial Surveillance Network (CHINET) in 2017 of the In Vitro Activities of Ceftazidime-Avibactam and Ceftolozane-Tazobactam against Clinical Isolates of Enterobacteriaceae and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2019, 63, e02431-18. [Google Scholar] [CrossRef] [Green Version]
- Cluck, D.; Lewis, P.; Stayer, B.; Spivey, J.; Moorman, J. Ceftolozane-tazobactam: A new-generation cephalosporin. Am. J. Health Pharm. AJHP Off. J. Am. Soc. Health Pharm. 2015, 72, 2135–2146. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Bassetti, M.; Duncan, L.R.; Castanheira, M. Ceftolozane/tazobactam activity against drug-resistant Enterobacteriaceae and Pseudomonas aeruginosa causing urinary tract and intraabdominal infections in Europe: Report from an antimicrobial surveillance programme (2012–2015). J. Antimicrob. Chemother. 2017, 72, 1386–1395. [Google Scholar] [CrossRef]
- Saran, O.; Sulik-Tyszka, B.; Basak, G.W.; Wróblewska, M.M. Activity of Ceftolozane/Tazobactam Against Gram-Negative Rods of the Family Enterobacteriaceae and Pseudomonas Spp. Isolated from Onco-Hematological Patients Hospitalized in a Clinical Hospital in Poland. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.; Duncan, L.R.; Mendes, R.E.; Sader, H.S.; Shortridge, D. Activity of Ceftolozane-Tazobactam against Pseudomonas aeruginosa and Enterobacteriaceae Isolates Collected from Respiratory Tract Specimens of Hospitalized Patients in the United States during 2013 to 2015. Antimicrob. Agents Chemother. 2018, 62, e02125-17. [Google Scholar] [CrossRef] [Green Version]
- Castanheira, M.; Doyle, T.B.; Mendes, R.E.; Sader, H.S. Comparative Activities of Ceftazidime-Avibactam and Ceftolozane-Tazobactam against Enterobacteriaceae Isolates Producing Extended-Spectrum β-Lactamases from U.S. Hospitals. Antimicrob. Agents Chemother. 2019, 63, e00160-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popejoy, M.W.; Paterson, D.L.; Cloutier, D.; Huntington, J.A.; Miller, B.; Bliss, C.A.; Steenbergen, J.N.; Hershberger, E.; Umeh, O.; Kaye, K.S. Efficacy of ceftolozane/tazobactam against urinary tract and intra-abdominal infections caused by ESBL-producing Escherichia coli and Klebsiella pneumoniae: A pooled analysis of Phase 3 clinical trials. J. Antimicrob. Chemother. 2017, 72, 268–272. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Kim, J.; Trinh, S. Meropenem-Vaborbactam (Vabomere(TM)): Another Option for Carbapenem-Resistant Enterobacteriaceae. P T 2019, 44, 110–113. [Google Scholar] [PubMed]
- Maraki, S.; Mavromanolaki, V.E.; Moraitis, P.; Stafylaki, D.; Kasimati, A.; Magkafouraki, E.; Scoulica, E. Ceftazidime-avibactam, meropenen-vaborbactam, and imipenem-relebactam in combination with aztreonam against multidrug-resistant, metallo-β-lactamase-producing Klebsiella pneumoniae. Eur. J. Clin. Microbiol. Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Haidar, G.; Clancy, C.J.; Chen, L.; Samanta, P.; Shields, R.K.; Kreiswirth, B.N.; Nguyen, M.H. Identifying Spectra of Activity and Therapeutic Niches for Ceftazidime-Avibactam and Imipenem-Relebactam against Carbapenem-Resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2017, 61, e00642-17. [Google Scholar] [CrossRef] [Green Version]
- Karlowsky, J.A.; Lob, S.H.; Kazmierczak, K.M.; Hawser, S.P.; Magnet, S.; Young, K.; Motyl, M.R.; Sahm, D.F. In vitro activity of imipenem/relebactam against Gram-negative ESKAPE pathogens isolated in 17 European countries: 2015 SMART surveillance programme. J. Antimicrob. Chemother. 2018, 73, 1872–1879. [Google Scholar] [CrossRef]
- Mushtaq, S.; Meunier, D.; Vickers, A.; Woodford, N.; Livermore, D.M. Activity of imipenem/relebactam against Pseudomonas aeruginosa producing ESBLs and carbapenemases. J. Antimicrob. Chemother. 2021, 76, 434–442. [Google Scholar] [CrossRef]
- Livermore, D.M.; Warner, M.; Mushtaq, S. Activity of MK-7655 combined with imipenem against Enterobacteriaceae and Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2013, 68, 2286–2290. [Google Scholar] [CrossRef] [Green Version]
- Canver, M.C.; Satlin, M.J.; Westblade, L.F.; Kreiswirth, B.N.; Chen, L.; Robertson, A.; Fauntleroy, K.; La Spina, M.; Callan, K.; Jenkins, S.G. Activity of Imipenem-Relebactam and Comparator Agents against Genetically Characterized Isolates of Carbapenem-Resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2019, 63, e00672-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Zhang, H.; Yu, Y.; Kong, H.; Duan, Q.; Wang, Y.; Zhang, S.; Sun, Z.; Liao, K.; Gu, L.; et al. In Vitro Activity of Imipenem/Relebactam Against Enterobacteriaceae Isolates Obtained from Intra-abdominal, Respiratory Tract, and Urinary Tract Infections in China: Study for Monitoring Antimicrobial Resistance Trends (SMART), 2015-2018. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2020, 71, S427–S435. [Google Scholar] [CrossRef] [PubMed]
- Lob, S.H.; Karlowsky, J.A.; Young, K.; Motyl, M.R.; Hawser, S.; Kothari, N.D.; Sahm, D.F. In vitro activity of imipenem-relebactam against resistant phenotypes of Enterobacteriaceae and Pseudomonas aeruginosa isolated from intraabdominal and urinary tract infection samples—SMART Surveillance Europe 2015–2017. J. Med. Microbiol. 2020, 69, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Ackley, R.; Roshdy, D.; Meredith, J.; Minor, S.; Anderson, W.E.; Capraro, G.A.; Polk, C. Meropenem-Vaborbactam versus Ceftazidime-Avibactam for Treatment of Carbapenem-Resistant Enterobacteriaceae Infections. Antimicrob. Agents Chemother. 2020, 64, e02313-19. [Google Scholar] [CrossRef]
- Kaye, K.S.; Vazquez, J.; Mathers, A.; Daikos, G.; Alexander, E.; Loutit, J.S.; Zhang, S.; Dudley, M.N.; Cornely, O. Clinical Outcomes of Serious Infections due to Carbapenem-Resistant Enterobacteriaceae (CRE) in TANGO II, a Phase 3, Randomized, Multi-National, Open-Label Trial of Meropenem-Vaborbactam (M-V) Vs. Best Available Therapy (BAT). Open Forum Infect. Dis. 2017, 4, S534–S535. [Google Scholar] [CrossRef] [Green Version]
- Kaye, K.S.; Bhowmick, T.; Metallidis, S.; Bleasdale, S.C.; Sagan, O.S.; Stus, V.; Vazquez, J.; Zaitsev, V.; Bidair, M.; Chorvat, E.; et al. Effect of Meropenem-Vaborbactam vs Piperacillin-Tazobactam on Clinical Cure or Improvement and Microbial Eradication in Complicated Urinary Tract Infection: The TANGO I Randomized Clinical Trial. JAMA 2018, 319, 788–799. [Google Scholar] [CrossRef]
- Carvalhaes, C.G.; Shortridge, D.; Sader, H.S.; Castanheira, M. Activity of Meropenem-Vaborbactam against Bacterial Isolates Causing Pneumonia in Patients in U.S. Hospitals during 2014 to 2018. Antimicrob. Agents Chemother. 2020, 64, e02177-19. [Google Scholar] [CrossRef] [Green Version]
- Moya, B.; Barcelo, I.M.; Bhagwat, S.; Patel, M.; Bou, G.; Papp-Wallace, K.M.; Bonomo, R.A.; Oliver, A. WCK 5107 (Zidebactam) and WCK 5153 Are Novel Inhibitors of PBP2 Showing Potent “β-Lactam Enhancer” Activity against Pseudomonas aeruginosa, Including Multidrug-Resistant Metallo-β-Lactamase-Producing High-Risk Clones. Antimicrob. Agents Chemother. 2017, 61, e02529-16. [Google Scholar] [CrossRef] [Green Version]
- Moya, B.; Barcelo, I.M.; Bhagwat, S.; Patel, M.; Bou, G.; Papp-Wallace, K.M.; Bonomo, R.A.; Oliver, A. Potent β-Lactam Enhancer Activity of Zidebactam and WCK 5153 against Acinetobacter baumannii, Including Carbapenemase-Producing Clinical Isolates. Antimicrob. Agents Chemother. 2017, 61, e01238-17. [Google Scholar] [CrossRef] [Green Version]
- Karlowsky, J.A.; Hackel, M.A.; Bouchillon, S.K.; Sahm, D.F. In Vitro Activity of WCK 5222 (Cefepime-Zidebactam) against Worldwide Collected Gram-Negative Bacilli Not Susceptible to Carbapenems. Antimicrob. Agents Chemother. 2020, 64, e01432-20. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y.; Yin, D.; Zheng, Y.; Wu, S.; Zhu, D.; Hu, F. In Vitro Activity of Cefepime-Zidebactam, Ceftazidime-Avibactam, and Other Comparators against Clinical Isolates of Enterobacterales, Pseudomonas aeruginosa, and Acinetobacter baumannii: Results from China Antimicrobial Surveillance Network (CHINET) in 2018. Antimicrob. Agents Chemother. 2020, 65, e01726-20. [Google Scholar] [CrossRef] [PubMed]
- Rodvold, K.A.; Gotfried, M.H.; Chugh, R.; Gupta, M.; Patel, A.; Chavan, R.; Yeole, R.; Friedland, H.D.; Bhatia, A. Plasma and Intrapulmonary Concentrations of Cefepime and Zidebactam following Intravenous Administration of WCK 5222 to Healthy Adult Subjects. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papp-Wallace, K.M. The latest advances in β-lactam/β-lactamase inhibitor combinations for the treatment of Gram-negative bacterial infections. Expert Opin. Pharmacother. 2019, 20, 2169–2184. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, Y.; Jia, P.; Zhu, Y.; Zhang, G.; Zhang, J.; Duan, S.; Kang, W.; Wang, T.; Jing, R.; et al. Global trends of antimicrobial susceptibility to ceftaroline and ceftazidime–avibactam: A surveillance study from the ATLAS program (2012–2016). Antimicrob. Resist. Infect. Control 2020, 9, 166. [Google Scholar] [CrossRef]
- Justo, J.A.; Mayer, S.M.; Pai, M.P.; Soriano, M.M.; Danziger, L.H.; Novak, R.M.; Rodvold, K.A. Pharmacokinetics of ceftaroline in normal body weight and obese (classes I, II, and III) healthy adult subjects. Antimicrob. Agents Chemother. 2015, 59, 3956–3965. [Google Scholar] [CrossRef] [Green Version]
- Vrancianu, C.O.; Gheorghe, I.; Dobre, E.-G.; Barbu, I.C.; Cristian, R.E.; Popa, M.; Lee, S.H.; Limban, C.; Vlad, I.M.; Chifiriuc, M.C. Emerging Strategies to Combat β-Lactamase Producing ESKAPE Pathogens. Int. J. Mol. Sci. 2020, 21, 8527. [Google Scholar] [CrossRef]
- Castanheira, M.; Sader, H.S.; Farrell, D.J.; Mendes, R.E.; Jones, R.N. Activity of ceftaroline-avibactam tested against Gram-negative organism populations, including strains expressing one or more β-lactamases and methicillin-resistant Staphylococcus aureus carrying various staphylococcal cassette chromosome mec types. Antimicrob. Agents Chemother. 2012, 56, 4779–4785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanheira, M.; Williams, G.; Jones, R.N.; Sader, H.S. Activity of ceftaroline-avibactam tested against contemporary Enterobacteriaceae isolates carrying β-lactamases prevalent in the United States. Microb. Drug Resist. 2014, 20, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Khalid, A.; Lubián, A.F.; Ma, L.; Lin, R.C.Y.; Iredell, J.R. Characterizing the role of porin mutations in susceptibility of beta lactamase producing Klebsiella pneumoniae isolates to ceftaroline and ceftaroline-avibactam. Int. J. Infect. Dis. 2020, 93, 252–257. [Google Scholar] [CrossRef]
- Wu, J.Y.; Srinivas, P.; Pogue, J.M. Cefiderocol: A Novel Agent for the Management of Multidrug-Resistant Gram-Negative Organisms. Infect. Dis. Ther. 2020, 9, 17–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.R.; Yeo, S. Cefiderocol, a New Siderophore Cephalosporin for the Treatment of Complicated Urinary Tract Infections Caused by Multidrug-Resistant Pathogens: Preclinical and Clinical Pharmacokinetics, Pharmacodynamics, Efficacy and Safety. Clin. Drug Investig. 2020, 40, 901–913. [Google Scholar] [CrossRef]
- Ito, A.; Sato, T.; Ota, M.; Takemura, M.; Nishikawa, T.; Toba, S.; Kohira, N.; Miyagawa, S.; Ishibashi, N.; Matsumoto, S.; et al. In Vitro Antibacterial Properties of Cefiderocol, a Novel Siderophore Cephalosporin, against Gram-Negative Bacteria. Antimicrob. Agents Chemother. 2018, 62, e01454-17. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, M.R.; Abdelhamed, A.M.; Good, C.E.; Rhoads, D.D.; Hujer, K.M.; Hujer, A.M.; Domitrovic, T.N.; Rudin, S.D.; Richter, S.S.; van Duin, D.; et al. ARGONAUT-I: Activity of Cefiderocol (S-649266), a Siderophore Cephalosporin, against Gram-Negative Bacteria, Including Carbapenem-Resistant Nonfermenters and Enterobacteriaceae with Defined Extended-Spectrum β-Lactamases and Carbapenemases. Antimicrob. Agents Chemother. 2019, 63, e01801-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlowsky, J.A.; Hackel, M.A.; Tsuji, M.; Yamano, Y.; Echols, R.; Sahm, D.F. In Vitro Activity of Cefiderocol, a Siderophore Cephalosporin, Against Gram-Negative Bacilli Isolated by Clinical Laboratories in North America and Europe in 2015-2016: SIDERO-WT-2015. Int. J. Antimicrob. Agents 2019, 53, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Monogue, M.L.; Tsuji, M.; Yamano, Y.; Echols, R.; Nicolau, D.P. Efficacy of Humanized Exposures of Cefiderocol (S-649266) against a Diverse Population of Gram-Negative Bacteria in a Murine Thigh Infection Model. Antimicrob. Agents Chemother. 2017, 61, e01022-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, S.; Singley, C.M.; Hoover, J.; Nakamura, R.; Echols, R.; Rittenhouse, S.; Tsuji, M.; Yamano, Y. Efficacy of Cefiderocol against Carbapenem-Resistant Gram-Negative Bacilli in Immunocompetent-Rat Respiratory Tract Infection Models Recreating Human Plasma Pharmacokinetics. Antimicrob. Agents Chemother. 2017, 61, e00700-17. [Google Scholar] [CrossRef] [Green Version]
- Bassetti, M.; Echols, R.; Matsunaga, Y.; Ariyasu, M.; Doi, Y.; Ferrer, R.; Lodise, T.P.; Naas, T.; Niki, Y.; Paterson, D.L.; et al. Efficacy and safety of cefiderocol or best available therapy for the treatment of serious infections caused by carbapenem-resistant Gram-negative bacteria (CREDIBLE-CR): A randomised, open-label, multicentre, pathogen-focused, descriptive, phase 3 trial. Lancet Infect. Dis. 2021, 21, 226–240. [Google Scholar] [CrossRef]
- Wunderink, R.G.; Matsunaga, Y.; Ariyasu, M.; Clevenbergh, P.; Echols, R.; Kaye, K.S.; Kollef, M.; Menon, A.; Pogue, J.M.; Shorr, A.F.; et al. Cefiderocol versus high-dose, extended-infusion meropenem for the treatment of Gram-negative nosocomial pneumonia (APEKS-NP): A randomised, double-blind, phase 3, non-inferiority trial. Lancet Infect. Dis. 2021, 21, 213–225. [Google Scholar] [CrossRef]
- Chew, K.L.; Tay, M.K.L.; Cheng, B.; Lin, R.T.P.; Octavia, S.; Teo, J.W.P. Aztreonam-Avibactam Combination Restores Susceptibility of Aztreonam in Dual-Carbapenemase-Producing Enterobacteriaceae. Antimicrob. Agents Chemother. 2018, 62, e00414-18. [Google Scholar] [CrossRef] [Green Version]
- Ransom, E.; Bhatnagar, A.; Patel, J.B.; Machado, M.-J.; Boyd, S.; Reese, N.; Lutgring, J.D.; Lonsway, D.; Anderson, K.; Brown, A.C.; et al. Validation of Aztreonam-Avibactam Susceptibility Testing Using Digitally Dispensed Custom Panels. J. Clin. Microbiol. 2020, 58, e01944-19. [Google Scholar] [CrossRef]
- Bhatnagar, A.; Ransom, E.M.; Machado, M.-J.; Boyd, S.; Reese, N.; Anderson, K.; Lonsway, D.; Elkins, C.A.; Rasheed, J.K.; Patel, J.B.; et al. Assessing the in vitro impact of ceftazidime on aztreonam/avibactam susceptibility testing for highly resistant MBL-producing Enterobacterales. J. Antimicrob. Chemother. 2021, 76, 979–983. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Lee, S.C.; Bae, M.; Sung, H.; Kim, M.-N.; Jung, J.; Kim, M.J.; Kim, S.-H.; Lee, S.-O.; Choi, S.-H.; et al. In Vitro Activities and Inoculum Effects of Ceftazidime-Avibactam and Aztreonam-Avibactam against Carbapenem-Resistant Enterobacterales Isolates from South Korea. Antibiotics 2020, 9, 912. [Google Scholar] [CrossRef]
- Monogue, M.L.; Giovagnoli, S.; Bissantz, C.; Zampaloni, C.; Nicolau, D.P. In Vivo Efficacy of Meropenem with a Novel Non-β-Lactam-β-Lactamase Inhibitor, Nacubactam, against Gram-Negative Organisms Exhibiting Various Resistance Mechanisms in a Murine Complicated Urinary Tract Infection Model. Antimicrob. Agents Chemother. 2018, 62, e02596-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morinaka, A.; Tsutsumi, Y.; Yamada, M.; Suzuki, K.; Watanabe, T.; Abe, T.; Furuuchi, T.; Inamura, S.; Sakamaki, Y.; Mitsuhashi, N.; et al. OP0595, a new diazabicyclooctane: Mode of action as a serine β-lactamase inhibitor, antibiotic and β-lactam “enhancer”. J. Antimicrob. Chemother. 2015, 70, 2779–2786. [Google Scholar] [CrossRef] [Green Version]
- Barnes, M.D.; Taracila, M.A.; Good, C.E.; Bajaksouzian, S.; Rojas, L.J.; van Duin, D.; Kreiswirth, B.N.; Jacobs, M.R.; Haldimann, A.; Papp-Wallace, K.M.; et al. Nacubactam Enhances Meropenem Activity against Carbapenem-Resistant Klebsiella pneumoniae Producing KPC. Antimicrob. Agents Chemother. 2019, 63, e00432-19. [Google Scholar] [CrossRef] [Green Version]
- Papp-Wallace, K.M.; Bonomo, R.A. New β-Lactamase Inhibitors in the Clinic. Infect. Dis. Clin. North Am. 2016, 30, 441–464. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, S.; Vickers, A.; Woodford, N.; Haldimann, A.; Livermore, D.M. Activity of nacubactam (RG6080/OP0595) combinations against MBL-producing Enterobacteriaceae. J. Antimicrob. Chemother. 2019, 74, 953–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landman, D.; Kelly, P.; Bäcker, M.; Babu, E.; Shah, N.; Bratu, S.; Quale, J. Antimicrobial activity of a novel aminoglycoside, ACHN-490, against Acinetobacter baumannii and Pseudomonas aeruginosa from New York City. J. Antimicrob. Chemother. 2011, 66, 332–334. [Google Scholar] [CrossRef] [Green Version]
- Karpiuk, I.; Tyski, S. Looking for new preparations for antibacterial therapy. IV. New antimicrobial agents from the aminoglycoside, macrolide and tetracycline groups in clinical trials. Przegl. Epidemiol. 2015, 69, 723–729. [Google Scholar]
- Syue, L.-S.; Chen, Y.-H.; Ko, W.-C.; Hsueh, P.-R. New drugs for the treatment of complicated intra-abdominal infections in the era of increasing antimicrobial resistance. Int. J. Antimicrob. Agents 2016, 47, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Mingeot-Leclercq, M.P.; Glupczynski, Y.; Tulkens, P.M. Aminoglycosides: Activity and resistance. Antimicrob. Agents Chemother. 1999, 43, 727–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, M.R.; Good, C.E.; Hujer, A.M.; Abdelhamed, A.M.; Rhoads, D.D.; Hujer, K.M.; Rudin, S.D.; Domitrovic, T.N.; Connolly, L.E.; Krause, K.M.; et al. ARGONAUT II Study of the In Vitro Activity of Plazomicin against Carbapenemase-Producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2020, 64, e00012-20. [Google Scholar] [CrossRef]
- Fleischmann, W.A.; Greenwood-Quaintance, K.E.; Patel, R. In Vitro Activity of Plazomicin Compared to Amikacin, Gentamicin, and Tobramycin against Multidrug-Resistant Aerobic Gram-Negative Bacilli. Antimicrob. Agents Chemother. 2020, 64, e01711-19. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.A.; Kulengowski, B.; Burgess, D.S. In vitro activity of plazomicin compared to other clinically relevant aminoglycosides in carbapenem-resistant Enterobacteriaceae. Diagn. Microbiol. Infect. Dis. 2020, 98, 115117. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.; Sader, H.S.; Mendes, R.E.; Jones, R.N. Activity of Plazomicin Tested against Enterobacterales Isolates Collected from U.S. Hospitals in 2016-2017: Effect of Different Breakpoint Criteria on Susceptibility Rates among Aminoglycosides. Antimicrob. Agents Chemother. 2020, 64, e02418-19. [Google Scholar] [CrossRef]
- Walkty, A.; Adam, H.; Baxter, M.; Denisuik, A.; Lagacé-Wiens, P.; Karlowsky, J.A.; Hoban, D.J.; Zhanel, G.G. In vitro activity of plazomicin against 5,015 gram-negative and gram-positive clinical isolates obtained from patients in canadian hospitals as part of the CANWARD study, 2011-2012. Antimicrob. Agents Chemother. 2014, 58, 2554–2563. [Google Scholar] [CrossRef] [Green Version]
- Denervaud-Tendon, V.; Poirel, L.; Connolly, L.E.; Krause, K.M.; Nordmann, P. Plazomicin activity against polymyxin-resistant Enterobacteriaceae, including MCR-1-producing isolates. J. Antimicrob. Chemother. 2017, 72, 2787–2791. [Google Scholar] [CrossRef]
- Shaeer, K.M.; Zmarlicka, M.T.; Chahine, E.B.; Piccicacco, N.; Cho, J.C. Plazomicin: A Next-Generation Aminoglycoside. Pharmacotherapy 2019, 39, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Doi, Y.; Wachino, J.-I.; Arakawa, Y. Aminoglycoside Resistance: The Emergence of Acquired 16S Ribosomal RNA Methyltransferases. Infect. Dis. Clin. North Am. 2016, 30, 523–537. [Google Scholar] [CrossRef] [Green Version]
- Castanheira, M.; Streit, J.M.; Serio, A.W.; Krause, K.M.; Flamm, R.K. 1345. Comparative Activity of Plazomicin and Other Aminoglycosides Against Enterobacteriaceae Isolates from Various Infection Sources from Hospitalized Patients in the United States. Open Forum Infect. Dis. 2018, 5, S411. [Google Scholar] [CrossRef]
- Cox, G.; Ejim, L.; Stogios, P.J.; Koteva, K.; Bordeleau, E.; Evdokimova, E.; Sieron, A.O.; Savchenko, A.; Serio, A.W.; Krause, K.M.; et al. Plazomicin Retains Antibiotic Activity against Most Aminoglycoside Modifying Enzymes. ACS Infect. Dis. 2018, 4, 980–987. [Google Scholar] [CrossRef]
- Reyes, N.; Aggen, J.B.; Kostrub, C.F. In vivo efficacy of the novel aminoglycoside ACHN-490 in murine infection models. Antimicrob. Agents Chemother. 2011, 55, 1728–1733. [Google Scholar] [CrossRef] [Green Version]
- Drusano, G.L.; Liu, W.; Fikes, S.; Cirz, R.; Robbins, N.; Kurhanewicz, S.; Rodriquez, J.; Brown, D.; Baluya, D.; Louie, A. Interaction of drug- and granulocyte-mediated killing of Pseudomonas aeruginosa in a murine pneumonia model. J. Infect. Dis. 2014, 210, 1319–1324. [Google Scholar] [CrossRef]
- Layton, R.C.; Brasel, T.; Gigliotti, A.; Barr, E.; Storch, S.; Myers, L.; Hobbs, C.; Koster, F. Primary pneumonic plague in the African Green monkey as a model for treatment efficacy evaluation. J. Med. Primatol. 2011, 40, 6–17. [Google Scholar] [CrossRef]
- Mega, W.; Overheim, K.; Cirz, R.; Cass, R.; Valderas, M.; Sherwood, R. Telemetry used to determine efficacy of intravenous plazomicin against inhaled tularemia in cynomolgus macaques (CM). J. Pharmacol. Toxicol. Methods 2014, 70, 334. [Google Scholar] [CrossRef]
- Gall, J.; Choi, T.; Riddle, V.; Van Wart, S.; Gibbons, J.A.; Seroogy, J. A Phase 1 Study of Intravenous Plazomicin in Healthy Adults to Assess Potential Effects on the QT/QTc Interval, Safety, and Pharmacokinetics. Clin. Pharmacol. Drug. Dev. 2019, 8, 1032–1041. [Google Scholar] [CrossRef]
- Connolly, L.E.; Riddle, V.; Cebrik, D.; Armstrong, E.S.; Miller, L.G. A Multicenter, Randomized, Double-Blind, Phase 2 Study of the Efficacy and Safety of Plazomicin Compared with Levofloxacin in the Treatment of Complicated Urinary Tract Infection and Acute Pyelonephritis. Antimicrob. Agents Chemother. 2018, 62, e01989-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuti, J.L.; Kim, A.; Cloutier, D.J.; Nicolau, D.P. Evaluation of Plazomicin, Tigecycline, and Meropenem Pharmacodynamic Exposure against Carbapenem-Resistant Enterobacteriaceae in Patients with Bloodstream Infection or Hospital-Acquired/Ventilator-Associated Pneumonia from the CARE Study (ACHN-490-007). Infect. Dis. Ther. 2019, 8, 383–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagenlehner, F.M.E.; Cloutier, D.J.; Komirenko, A.S.; Cebrik, D.S.; Krause, K.M.; Keepers, T.R.; Connolly, L.E.; Miller, L.G.; Friedland, I.; Dwyer, J.P. Once-Daily Plazomicin for Complicated Urinary Tract Infections. N. Engl. J. Med. 2019, 380, 729–740. [Google Scholar] [CrossRef]
- Solomkin, J.; Evans, D.; Slepavicius, A.; Lee, P.; Marsh, A.; Tsai, L.; Sutcliffe, J.A.; Horn, P. Assessing the Efficacy and Safety of Eravacycline vs Ertapenem in Complicated Intra-abdominal Infections in the Investigating Gram-Negative Infections Treated With Eravacycline (IGNITE 1) Trial: A Randomized Clinical Trial. JAMA Surg. 2017, 152, 224–232. [Google Scholar] [CrossRef]
- Lee, Y.R.; Burton, C.E. Eravacycline, a newly approved fluorocycline. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1787–1794. [Google Scholar] [CrossRef]
- Connors, K.P.; Housman, S.T.; Pope, J.S.; Russomanno, J.; Salerno, E.; Shore, E.; Redican, S.; Nicolau, D.P. Phase I, Open-Label, Safety and Pharmacokinetic Study to Assess Bronchopulmonary Disposition of Intravenous Eravacycline in Healthy Men and Women. Antimicrob. Agents Chemother. 2014, 58, 2113–2118. [Google Scholar] [CrossRef] [Green Version]
- Solomkin, J.S.; Ramesh, M.K.; Cesnauskas, G.; Novikovs, N.; Stefanova, P.; Sutcliffe, J.A.; Walpole, S.M.; Horn, P.T. Phase 2, randomized, double-blind study of the efficacy and safety of two dose regimens of eravacycline versus ertapenem for adult community-acquired complicated intra-abdominal infections. Antimicrob. Agents Chemother. 2014, 58, 1847–1854. [Google Scholar] [CrossRef] [Green Version]
- Solomkin, J.S.; Gardovskis, J.; Lawrence, K.; Montravers, P.; Sway, A.; Evans, D.; Tsai, L. IGNITE4: Results of a Phase 3, Randomized, Multicenter, Prospective Trial of Eravacycline vs Meropenem in the Treatment of Complicated Intraabdominal Infections. Clin. Infect. Dis. 2019, 69, 921–929. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, J.C. Omadacycline: A Modernized Tetracycline. Clin. Infect. Dis. 2019, 69, S1–S5. [Google Scholar] [CrossRef]
- Watkins, R.R.; Deresinski, S. Omadacycline: A Novel Tetracycline Derivative with Oral and Intravenous Formulations. Clin. Infect. Dis. 2019, 69, 890–896. [Google Scholar] [CrossRef]
- Karlowsky, J.A.; Steenbergen, J.; Zhanel, G.G. Microbiology and Preclinical Review of Omadacycline. Clin. Infect. Dis. 2019, 69, S6–S15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhanel, G.G.; Esquivel, J.; Zelenitsky, S.; Lawrence, C.K.; Adam, H.J.; Golden, A.; Hink, R.; Berry, L.; Schweizer, F.; Zhanel, M.A.; et al. Omadacycline: A Novel Oral and Intravenous Aminomethylcycline Antibiotic Agent. Drugs 2020, 80, 285–313. [Google Scholar] [CrossRef]
- Huband, M.D.; Pfaller, M.A.; Shortridge, D.; Flamm, R.K. Surveillance of omadacycline activity tested against clinical isolates from the United States and Europe: Results from the SENTRY Antimicrobial Surveillance Programme, 2017. J. Glob. Antimicrob. Resist. 2019, 19, 56–63. [Google Scholar] [CrossRef]
- Lin, W.; Flarakos, J.; Du, Y.; Hu, W.; He, H.; Mangold, J.; Tanaka, S.K.; Villano, S. Pharmacokinetics, Distribution, Metabolism, and Excretion of Omadacycline following a Single Intravenous or Oral Dose of 14C-Omadacycline in Rats. Antimicrob. Agents Chemother. 2017, 61, e01784-16. [Google Scholar] [CrossRef] [Green Version]
- Lepak, A.J.; Zhao, M.; Marchillo, K.; VanHecker, J.; Andes, D.R. In Vivo Pharmacodynamic Evaluation of Omadacycline (PTK 0796) against Streptococcus pneumoniae in the Murine Pneumonia Model. Antimicrob. Agents Chemother. 2017, 61, e02368-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepak, A.J.; Zhao, M.; Marchillo, K.; VanHecker, J.; Andes, D.R. In Vivo Pharmacodynamics of Omadacycline against Staphylococcus aureus in the Neutropenic Murine Thigh Infection Model. Antimicrob. Agents Chemother. 2019, 63, e00624-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noel, G.J.; Draper, M.P.; Hait, H.; Tanaka, S.K.; Arbeit, R.D. A randomized, evaluator-blind, phase 2 study comparing the safety and efficacy of omadacycline to those of linezolid for treatment of complicated skin and skin structure infections. Antimicrob. Agents Chemother. 2012, 56, 5650–5654. [Google Scholar] [CrossRef] [Green Version]
- O’Riordan, W.; Green, S.; Overcash, J.S.; Puljiz, I.; Metallidis, S.; Gardovskis, J.; Garrity-Ryan, L.; Das, A.F.; Tzanis, E.; Eckburg, P.B.; et al. Omadacycline for Acute Bacterial Skin and Skin-Structure Infections. N. Engl. J. Med. 2019, 380, 528–538. [Google Scholar] [CrossRef] [PubMed]
- O’Riordan, W.; Cardenas, C.; Shin, E.; Sirbu, A.; Garrity-Ryan, L.; Das, A.F.; Eckburg, P.B.; Manley, A.; Steenbergen, J.N.; Tzanis, E.; et al. Once-daily oral omadacycline versus twice-daily oral linezolid for acute bacterial skin and skin structure infections (OASIS-2): A phase 3, double-blind, multicentre, randomised, controlled, non-inferiority trial. Lancet. Infect. Dis. 2019, 19, 1080–1090. [Google Scholar] [CrossRef]
- Stets, R.; Popescu, M.; Gonong, J.R.; Mitha, I.; Nseir, W.; Madej, A.; Kirsch, C.; Das, A.F.; Garrity-Ryan, L.; Steenbergen, J.N.; et al. Omadacycline for Community-Acquired Bacterial Pneumonia. N. Engl. J. Med. 2019, 380, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Mogle, B.T.; Steele, J.M.; Thomas, S.J.; Bohan, K.H.; Kufel, W.D. Clinical review of delafloxacin: A novel anionic fluoroquinolone. J. Antimicrob. Chemother. 2018, 73, 1439–1451. [Google Scholar] [CrossRef] [Green Version]
- Kocsis, B.; Domokos, J.; Szabo, D. Chemical structure and pharmacokinetics of novel quinolone agents represented by avarofloxacin, delafloxacin, finafloxacin, zabofloxacin and nemonoxacin. Ann. Clin. Microbiol. Antimicrob. 2016, 15, 34. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, S.C.J.; Mercuro, N.J.; Davis, S.L.; Rybak, M.J. Delafloxacin: Place in Therapy and Review of Microbiologic, Clinical and Pharmacologic Properties. Infect. Dis. Ther. 2018, 7, 197–217. [Google Scholar] [CrossRef] [Green Version]
- Scott, L.J. Delafloxacin: A Review in Acute Bacterial Skin and Skin Structure Infections. Drugs 2020, 80, 1247–1258. [Google Scholar] [CrossRef]
- McCurdy, S.; Lawrence, L.; Quintas, M.; Woosley, L.; Flamm, R.; Tseng, C.; Cammarata, S. In Vitro Activity of Delafloxacin and Microbiological Response against Fluoroquinolone-Susceptible and Nonsusceptible Staphylococcus aureus Isolates from Two Phase 3 Studies of Acute Bacterial Skin and Skin Structure Infections. Antimicrob. Agents Chemother. 2017, 61, e00772-17. [Google Scholar] [CrossRef] [Green Version]
- O’Riordan, W.; McManus, A.; Teras, J.; Poromanski, I.; Cruz-Saldariagga, M.; Quintas, M.; Lawrence, L.; Liang, S.; Cammarata, S. A Comparison of the Efficacy and Safety of Intravenous Followed by Oral Delafloxacin With Vancomycin Plus Aztreonam for the Treatment of Acute Bacterial Skin and Skin Structure Infections: A Phase 3, Multinational, Double-Blind, Randomized Study. Clin. Infect. Dis. 2018, 67, 657–666. [Google Scholar] [CrossRef]
- McCurdy, S.; Keedy, K.; Lawrence, L.; Nenninger, A.; Sheets, A.; Quintas, M.; Cammarata, S. Efficacy of Delafloxacin versus Moxifloxacin against Bacterial Respiratory Pathogens in Adults with Community-Acquired Bacterial Pneumonia (CABP): Microbiology Results from the Delafloxacin Phase 3 CABP Trial. Antimicrob. Agents Chemother. 2020, 64, e01949-19. [Google Scholar] [CrossRef] [Green Version]
- McCurdy, S.; Nenninger, A.; Sheets, A.; Keedy, K.; Lawrence, L.; Quintas, M.; Cammarata, S. Efficacy of delafloxacin versus moxifloxacin against atypical bacterial respiratory pathogens in adults with community-acquired bacterial pneumonia (CABP): Data from the Delafloxacin Phase 3 CABP Trial. Int. J. Infect. Dis. 2020, 97, 374–379. [Google Scholar] [CrossRef]
- Hook, E.W., 3rd; Golden, M.R.; Taylor, S.N.; Henry, E.; Tseng, C.; Workowski, K.A.; Swerdlow, J.; Nenninger, A.; Cammarata, S. Efficacy and Safety of Single-Dose Oral Delafloxacin Compared with Intramuscular Ceftriaxone for Uncomplicated Gonorrhea Treatment: An Open-Label, Noninferiority, Phase 3, Multicenter, Randomized Study. Sex. Transm. Dis. 2019, 46, 279–286. [Google Scholar] [CrossRef]
- Pullman, J.; Gardovskis, J.; Farley, B.; Sun, E.; Quintas, M.; Lawrence, L.; Ling, R.; Cammarata, S. Efficacy and safety of delafloxacin compared with vancomycin plus aztreonam for acute bacterial skin and skin structure infections: A Phase 3, double-blind, randomized study. J. Antimicrob. Chemother. 2017, 72, 3471–3480. [Google Scholar] [CrossRef] [Green Version]
- O’Riordan, W.; Mehra, P.; Manos, P.; Kingsley, J.; Lawrence, L.; Cammarata, S. A randomized phase 2 study comparing two doses of delafloxacin with tigecycline in adults with complicated skin and skin-structure infections. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2015, 30, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingsley, J.; Mehra, P.; Lawrence, L.E.; Henry, E.; Duffy, E.; Cammarata, S.K.; Pullman, J. A randomized, double-blind, Phase 2 study to evaluate subjective and objective outcomes in patients with acute bacterial skin and skin structure infections treated with delafloxacin, linezolid or vancomycin. J. Antimicrob. Chemother. 2016, 71, 821–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peyclit, L.; Baron, S.A.; Rolain, J.-M. Drug Repurposing to Fight Colistin and Carbapenem-Resistant Bacteria. Front. Cell. Infect. Microbiol. 2019, 9, 193. [Google Scholar] [CrossRef] [PubMed]
- Kaul, G.; Shukla, M.; Dasgupta, A.; Chopra, S. Update on drug-repurposing: Is it useful for tackling antimicrobial resistance? Future Microbiol. 2019, 14, 829–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miró-Canturri, A.; Ayerbe-Algaba, R.; Smani, Y. Drug Repurposing for the Treatment of Bacterial and Fungal Infections. Front. Microbiol. 2019, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liu, Y.; Coates, A. Azidothymidine Produces Synergistic Activity in Combination with Colistin against Antibiotic-Resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2019, 63, e01630-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, S.M.S.; Sioson, J.S.P.; Yap, J.M.; Ng, F.M.; Ching, H.S.V.; Teo, J.W.P.; Jureen, R.; Hill, J.; Chia, C.S.B. Repurposing Zidovudine in combination with Tigecycline for treating carbapenem-resistant Enterobacteriaceae infections. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2018, 37, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Carlson-Banning, K.M.; Chou, A.; Liu, Z.; Hamill, R.J.; Song, Y.; Zechiedrich, L. Toward repurposing ciclopirox as an antibiotic against drug-resistant Acinetobacter baumannii, Escherichia coli, and Klebsiella pneumoniae. PLoS ONE 2013, 8, e69646. [Google Scholar] [CrossRef] [Green Version]
- Gajdošová, M.; Vetchý, D.; Muselík, J.; Gajdziok, J.; Juřica, J.; Vetchá, M.; Hauptman, K.; Jekl, V. Bilayer mucoadhesive buccal films with prolonged release of ciclopirox olamine for the treatment of oral candidiasis: In vitro development, ex vivo permeation testing, pharmacokinetic and efficacy study in rabbits. Int. J. Pharm. 2021, 592, 120086. [Google Scholar] [CrossRef]
- Hijazi, S.; Visaggio, D.; Pirolo, M.; Frangipani, E.; Bernstein, L.; Visca, P. Antimicrobial Activity of Gallium Compounds on ESKAPE Pathogens. Front. Cell. Infect. Microbiol. 2018, 8, 316. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.B.; Wang, J.; Doi, Y.; Velkov, T.; Bergen, P.J.; Li, J. Novel Polymyxin Combination with Antineoplastic Mitotane Improved the Bacterial Killing Against Polymyxin-Resistant Multidrug-Resistant Gram-Negative Pathogens. Front. Microbiol. 2018, 9, 721. [Google Scholar] [CrossRef] [Green Version]
- Hussein, M.H.; Schneider, E.K.; Elliott, A.G.; Han, M.; Reyes-Ortega, F.; Morris, F.; Blastovich, M.A.T.; Jasim, R.; Currie, B.; Mayo, M.; et al. From Breast Cancer to Antimicrobial: Combating Extremely Resistant Gram-Negative “Superbugs” Using Novel Combinations of Polymyxin B with Selective Estrogen Receptor Modulators. Microb. Drug Resist. 2017, 23, 640–650. [Google Scholar] [CrossRef]
- Otto, R.G.; van Gorp, E.; Kloezen, W.; Meletiadis, J.; van den Berg, S.; Mouton, J.W. An alternative strategy for combination therapy: Interactions between polymyxin B and non-antibiotics. Int. J. Antimicrob. Agents 2019, 53, 34–39. [Google Scholar] [CrossRef]
- Welton, T. Ionic liquids: A brief history. Biophys. Rev. 2018, 10, 691–706. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, J.N.; Gilmore, B.F. The antimicrobial potential of ionic liquids: A source of chemical diversity for infection and biofilm control. Int. J. Antimicrob. Agents 2015, 46, 131–139. [Google Scholar] [CrossRef]
- Singh, S.K.; Savoy, A.W. Ionic liquids synthesis and applications: An overview. J. Mol. Liq. 2020, 297, 112038. [Google Scholar] [CrossRef]
- Agatemor, C.; Ibsen, K.N.; Tanner, E.E.L.; Mitragotri, S. Ionic liquids for addressing unmet needs in healthcare. Bioeng. Transl. Med. 2018, 3, 7–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, P.; Pillai, V.V.S.; Benedetto, A. Mechanisms of action of ionic liquids on living cells: The state of the art. Biophys. Rev. 2020, 12, 1187–1215. [Google Scholar] [CrossRef] [PubMed]
- Łuczak, J.; Jungnickel, C.; Łącka, I.; Stolte, S.; Hupka, J. Antimicrobial and surface activity of 1-alkyl-3-methylimidazolium derivatives. Green Chem. 2010, 12, 593–601. [Google Scholar] [CrossRef]
- Miskiewicz, A.; Ceranowicz, P.; Szymczak, M.; Bartuś, K.; Kowalczyk, P. The Use of Liquids Ionic Fluids as Pharmaceutically Active Substances Helpful in Combating Nosocomial Infections Induced by Klebsiella Pneumoniae New Delhi Strain, Acinetobacter Baumannii and Enterococcus Species. Int. J. Mol. Sci. 2018, 19, 2779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martini Garcia, I.; Jung Ferreira, C.; de Souza, V.S.; Castelo Branco Leitune, V.; Samuel, S.M.W.; de Souza Balbinot, G.; de Souza da Motta, A.; Visioli, F.; Damiani Scholten, J.; Mezzomo Collares, F. Ionic liquid as antibacterial agent for an experimental orthodontic adhesive. Dent. Mater. 2019, 35, 1155–1165. [Google Scholar] [CrossRef]
- Hodyna, D.; Bardeau, J.-F.; Metelytsia, L.; Riabov, S.; Kobrina, L.; Laptiy, S.; Kalashnikova, L.; Parkhomenko, V.; Tarasyuk, O.; Rogalsky, S. Efficient antimicrobial activity and reduced toxicity of 1-dodecyl-3-methylimidazolium tetrafluoroborate ionic liquid/β-cyclodextrin complex. Chem. Eng. J. 2016, 284, 1136–1145. [Google Scholar] [CrossRef]
- Benedetto, A.; Ballone, P. Room-Temperature Ionic Liquids and Biomembranes: Setting the Stage for Applications in Pharmacology, Biomedicine, and Bionanotechnology. Langmuir 2018, 34, 9579–9597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jha, G.; Sahu, P.K.; Panda, S.; Singh, D.V.; Patole, S.; Mohapatra, H.; Sarkar, M. Synthesis, Photophysical Studies on Some Anthracene–based Ionic Liquids and their Application as Biofilm Formation Inhibitor. ChemistrySelect 2017, 2, 2426–2432. [Google Scholar] [CrossRef]
- Egorova, K.S.; Gordeev, E.G.; Ananikov, V.P. Biological Activity of Ionic Liquids and Their Application in Pharmaceutics and Medicine. Chem. Rev. 2017, 117, 7132–7189. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Xu, Q.; Zheng, Z.; Zhou, S.; Mao, H.; Wang, B.; Yan, F. Intrinsically Antibacterial Poly(ionic liquid) Membranes: The Synergistic Effect of Anions. ACS Macro Lett. 2015, 4, 1094–1098. [Google Scholar] [CrossRef]
- Ferraz, R.; Silva, D.; Dias, A.R.; Dias, V.; Santos, M.M.; Pinheiro, L.; Prudêncio, C.; Noronha, J.P.; Petrovski, Ž.; Branco, L.C. Synthesis and Antibacterial Activity of Ionic Liquids and Organic Salts Based on Penicillin G and Amoxicillin hydrolysate Derivatives against Resistant Bacteria. Pharmaceutics 2020, 12, 221. [Google Scholar] [CrossRef] [Green Version]
- Cole, M.R.; Hobden, J.A.; Warner, I.M. Recycling antibiotics into GUMBOS: A new combination strategy to combat multi-drug-resistant bacteria. Molecules 2015, 20, 6466–6487. [Google Scholar] [CrossRef]
- Dang, T.; Nizamov, I.S.; Salikhov, R.Z.; Sabirzyanova, L.R.; Vorobev, V.V.; Burganova, T.I.; Shaidoullina, M.M.; Batyeva, E.S.; Cherkasov, R.A.; Abdullin, T.I. Synthesis and characterization of pyridoxine, nicotine and nicotinamide salts of dithiophosphoric acids as antibacterial agents against resistant wound infection. Bioorg. Med. Chem. 2019, 27, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, R.; Teixeira, V.; Rodrigues, D.; Fernandes, R.; Noronha, J.P.; Branco, L.C.; Petrovski, Ž.; Prudêncio, C. Antibacterial activity of Ionic Liquids based on ampicillin against resistant bacteria. RSC Adv. 2013, 4, 4301–4307. [Google Scholar] [CrossRef] [Green Version]
- Prudêncio, C.; Vieira, M.; Van der Auweraer, S.; Ferraz, R. Recycling Old Antibiotics with Ionic Liquids. Antibiotics 2020, 9, 578. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Gonzalez, M.; Kong, C.; Weir, S.; Socha, A.M. Synthesis, antibiotic structure-activity relationships, and cellulose dissolution studies of new room-temperature ionic liquids derived from lignin. Biotechnol. Biofuels 2021, 14, 47. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Cataldo, V.A.; Dimke, T.; Stephan, I.; Guterman, R. Antibacterial and Degradable Thioimidazolium Poly(ionic liquid). ACS Sustain. Chem. Eng. 2020, 8, 8419–8424. [Google Scholar] [CrossRef]
- Zheng, Z.; Xu, Q.; Guo, J.; Qin, J.; Mao, H.; Wang, B.; Yan, F. Structure-Antibacterial Activity Relationships of Imidazolium-Type Ionic Liquid Monomers, Poly(ionic liquids) and Poly(ionic liquid) Membranes: Effect of Alkyl Chain Length and Cations. ACS Appl. Mater. Interfaces 2016, 8, 12684–12692. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Cao, S.; Horn, A.L. Emerging Applications of Machine Learning in Food Safety. Annu. Rev. Food Sci. Technol. 2021, 12, 513–538. [Google Scholar] [CrossRef]
- Grear, T.; Avery, C.; Patterson, J.; Jacobs, D.J. Molecular function recognition by supervised projection pursuit machine learning. Sci. Rep. 2021, 11, 4247. [Google Scholar] [CrossRef] [PubMed]
- Stokes, J.M.; Yang, K.; Swanson, K.; Jin, W.; Cubillos-Ruiz, A.; Donghia, N.M.; MacNair, C.R.; French, S.; Carfrae, L.A.; Bloom-Ackermann, Z.; et al. A Deep Learning Approach to Antibiotic Discovery. Cell 2020, 180, 688–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, N.M.; Lenhard, J.R.; Boissonneault, K.R.; Landersdorfer, C.B.; Bulitta, J.B.; Holden, P.N.; Forrest, A.; Nation, R.L.; Li, J.; Tsuji, B.T. Using machine learning to optimize antibiotic combinations: Dosing strategies for meropenem and polymyxin B against carbapenem-resistant Acinetobacter baumannii. Clin. Microbiol. Infect. 2020, 26, 1207–1213. [Google Scholar] [CrossRef]
- Chapman, J.; Orrell-Trigg, R.; Kwoon, K.Y.; Truong, V.K.; Cozzolino, D. A high-throughput and machine learning resistance monitoring system to determine the point of resistance for Escherichia coli with tetracycline: Combining UV-visible spectrophotometry with principal component analysis. Biotechnol. Bioeng. 2021, 118, 1511–1519. [Google Scholar] [CrossRef]
- Parvaiz, N.; Ahmad, F.; Yu, W.; MacKerell, A.D.J.; Azam, S.S. Discovery of beta-lactamase CMY-10 inhibitors for combination therapy against multi-drug resistant Enterobacteriaceae. PLoS ONE 2021, 16, e0244967. [Google Scholar] [CrossRef] [PubMed]
- Mienda, B.S.; Dräger, A. Genome-Scale Metabolic Modeling of Escherichia coli and Its Chassis Design for Synthetic Biology Applications. Methods Mol. Biol. 2021, 2189, 217–229. [Google Scholar] [CrossRef]
- Torres, M.D.T.; Cao, J.; Franco, O.L.; Lu, T.K.; de la Fuente-Nunez, C. Synthetic Biology and Computer-Based Frameworks for Antimicrobial Peptide Discovery. ACS Nano 2021, 15, 2143–2164. [Google Scholar] [CrossRef]
- Li, Q.; Pellegrino, J.; Lee, D.J.; Tran, A.A.; Chaires, H.A.; Wang, R.; Park, J.E.; Ji, K.; Chow, D.; Zhang, N.; et al. Synthetic group A streptogramin antibiotics that overcome Vat resistance. Nature 2020, 586, 145–150. [Google Scholar] [CrossRef]
- Gerstmans, H.; Criel, B.; Briers, Y. Synthetic biology of modular endolysins. Biotechnol. Adv. 2018, 36, 624–640. [Google Scholar] [CrossRef]
- Doi, Y.; Paterson, D.L. Carbapenemase-producing Enterobacteriaceae. Semin. Respir. Crit. Care Med. 2015, 36, 74–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, S.A.; Gray, T.J.; Gottlieb, T. Healthcare-acquired infections: Prevention strategies. Intern. Med. J. 2017, 47, 1341–1351. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.W.K.; Buising, K.L.; Jeremiah, C.J.; Cronin, K.; Poy Lorenzo, Y.S.; Howden, B.P.; Kwong, J.; Cocks, J.; Blood, A.; Greenough, J.; et al. Managing a nosocomial outbreak of carbapenem-resistant Klebsiella pneumoniae: An early Australian hospital experience. Intern. Med. J. 2015, 45, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Guidelines on Hand Hygiene in Health Care: First Global Patient Safety Challenge Clean Care Is Safer Care; WHO: Geneva, Switzerland, 2009; ISBN 978-92-4-159790-6. [Google Scholar]
- Pittet, D.; Allegranzi, B.; Sax, H.; Dharan, S.; Pessoa-Silva, C.L.; Donaldson, L.; Boyce, J.M. Evidence-based model for hand transmission during patient care and the role of improved practices. Lancet Infect. Dis. 2006, 6, 641–652. [Google Scholar] [CrossRef]
- Allegranzi, B.; Pittet, D. Role of hand hygiene in healthcare-associated infection prevention. J. Hosp. Infect. 2009, 73, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Sax, H.; Allegranzi, B.; Uçkay, I.; Larson, E.; Boyce, J.; Pittet, D. “My five moments for hand hygiene”: A user-centred design approach to understand, train, monitor and report hand hygiene. J. Hosp. Infect. 2007, 67, 9–21. [Google Scholar] [CrossRef]
- Barlam, T.F.; Cosgrove, S.E.; Abbo, L.M.; MacDougall, C.; Schuetz, A.N.; Septimus, E.J.; Srinivasan, A.; Dellit, T.H.; Falck-Ytter, Y.T.; Fishman, N.O.; et al. Implementing an Antibiotic Stewardship Program: Guidelines by the Infectious Diseases Society of America and the Society for Healthcare Epidemiology of America. Clin. Infect. Dis. 2016, 62, e51–e77. [Google Scholar] [CrossRef]
- Moehring, R.W.; Anderson, D.J. Antimicrobial stewardship as part of the infection prevention effort. Curr. Infect. Dis. Rep. 2012, 14, 592–600. [Google Scholar] [CrossRef]
- Suay-García, B.; Pérez-Gracia, M.T. Present and Future of Carbapenem-resistant Enterobacteriaceae (CRE) Infections. Antibiotics 2019, 8, 122. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-R.; Eom, Y.-B. Synergistic Activity of Equol and Meropenem against Carbapenem-Resistant Escherichia coli. Antibiotics 2021, 10, 161. [Google Scholar] [CrossRef]
- Noval, M.; Banoub, M.; Claeys, K.C.; Heil, E. The Battle Is on: New Beta-Lactams for the Treatment of Multidrug-Resistant Gram-Negative Organisms. Curr. Infect. Dis. Rep. 2020, 22, 1. [Google Scholar] [CrossRef] [PubMed]
- Campanella, T.A.; Gallagher, J.C. A Clinical Review and Critical Evaluation of Imipenem-Relebactam: Evidence to Date. Infect. Drug Resist. 2020, 13, 4297–4308. [Google Scholar] [CrossRef] [PubMed]
- Endimiani, A.; Hujer, K.M.; Hujer, A.M.; Armstrong, E.S.; Choudhary, Y.; Aggen, J.B.; Bonomo, R.A. ACHN-490, a neoglycoside with potent in vitro activity against multidrug-resistant Klebsiella pneumoniae isolates. Antimicrob. Agents Chemother. 2009, 53, 4504–4507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papp-Wallace, K.M.; Nguyen, N.Q.; Jacobs, M.R.; Bethel, C.R.; Barnes, M.D.; Kumar, V.; Bajaksouzian, S.; Rudin, S.D.; Rather, P.N.; Bhavsar, S.; et al. Strategic Approaches to Overcome Resistance against Gram-Negative Pathogens Using β-Lactamase Inhibitors and β-Lactam Enhancers: Activity of Three Novel Diazabicyclooctanes WCK 5153, Zidebactam (WCK 5107), and WCK 4234. J. Med. Chem. 2018, 61, 4067–4086. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Drug | Mechanism of Action | Commercially Available | Indications | Limitations |
|---|---|---|---|---|
| Ceftazidime/avibactam | Cell wall synthesis inhibitor | Yes; FDA-approved in 2015 as Allergan | cUTI, cIAI, BSI, pneumonia | Occurrence of resistance |
| Ceftolozane/tazobactam | Inhibition of PBPs | Yes; FDA-approved in 2014 (Zerbaxa) | cUTI, cIAI | Occurrence of resistance |
| Meropenem/varbobactam | Cell wall synthesis inhibitor | Yes; FDA-approved in 2017 as Vabomere | cUTI, cIAI, BSI, pneumonia | Ocurrence of resistance |
| Ceftaroline/avibactam | Inhibition of PBPs | Under clinical investigation | cUTI | Occurrence of resistance due to mutations in KPC-producing Enterobacteriaceae |
| Cefepime/Zidebactam (WCK 5222) | Direct inhibition of β-lactamases or PBP2 inhibition | Under clinical development | cUTI, cIAI, SI, pneumonia | Ocurrence of resistance |
| Imipenem/cilastatin-relebactam | Renal dehydro-peptidase inhibitor/β-lactamase inhibitor | Yes; FDA-approved in 2019 as Recarbrio | cUTI, cIAI, pneumonia | Severe hypersensitivity reactions |
| Aztreonam/avibactam | Cell wall synthesis inhibitor | Under clinical development | cIAI | Likelihood of resistance among MBL- and AmpC-co-producing K. pneumoniae |
| Meropenem/nacubactam | Cell wall synthesis inhibitor | Under clinical investigation | cIAI | Occurrence of resistance; alterations of renal function |
| Plazomicin | Protein synthesis inhibitor | Yes, FDA-approved in 2018 as ZEMDRI | cUTI, BSI, pneumonia | Ineffective against MBL-producers |
| Eravacycline | Protein synthesis inhibitor | Yes, FDA-approved in 2018 as XERAVA | cIAI, pneumonia | Not indicated for the treatment of cUTI |
| Cefiderocol | Cell wall synthesis inhibitor | Yes, FDA-approved in 2019 as Fetroja | cUTI | Under investigation in clinical trials |
| Omadacycline | Protein synthesis inhibitor | Yes, FDA-approved in 2018 as NUZYRA | cUTI, pneumonia, acute SI | Limited action against ESBL-producing K. pneumoniae |
| Delafloxacin | Protein synthesis inhibitor (topoisomerase IV and DNA gyrase) | Yes, FDA-approved in 2017 as Baxdela (USA); Quofenix (EU). | Acute SI, pneumonia | Peripheral neuropathy and central nervous system effects |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vrancianu, C.O.; Dobre, E.G.; Gheorghe, I.; Barbu, I.; Cristian, R.E.; Chifiriuc, M.C. Present and Future Perspectives on Therapeutic Options for Carbapenemase-Producing Enterobacterales Infections. Microorganisms 2021, 9, 730. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040730
Vrancianu CO, Dobre EG, Gheorghe I, Barbu I, Cristian RE, Chifiriuc MC. Present and Future Perspectives on Therapeutic Options for Carbapenemase-Producing Enterobacterales Infections. Microorganisms. 2021; 9(4):730. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040730
Chicago/Turabian StyleVrancianu, Corneliu Ovidiu, Elena Georgiana Dobre, Irina Gheorghe, Ilda Barbu, Roxana Elena Cristian, and Mariana Carmen Chifiriuc. 2021. "Present and Future Perspectives on Therapeutic Options for Carbapenemase-Producing Enterobacterales Infections" Microorganisms 9, no. 4: 730. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040730