
Multi-Drug Resistant Plasmids with ESBL/AmpC and mcr-5.1 in Paraguayan Poultry Farms: The Linkage of Antibiotic Resistance and Hatcheries

 , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
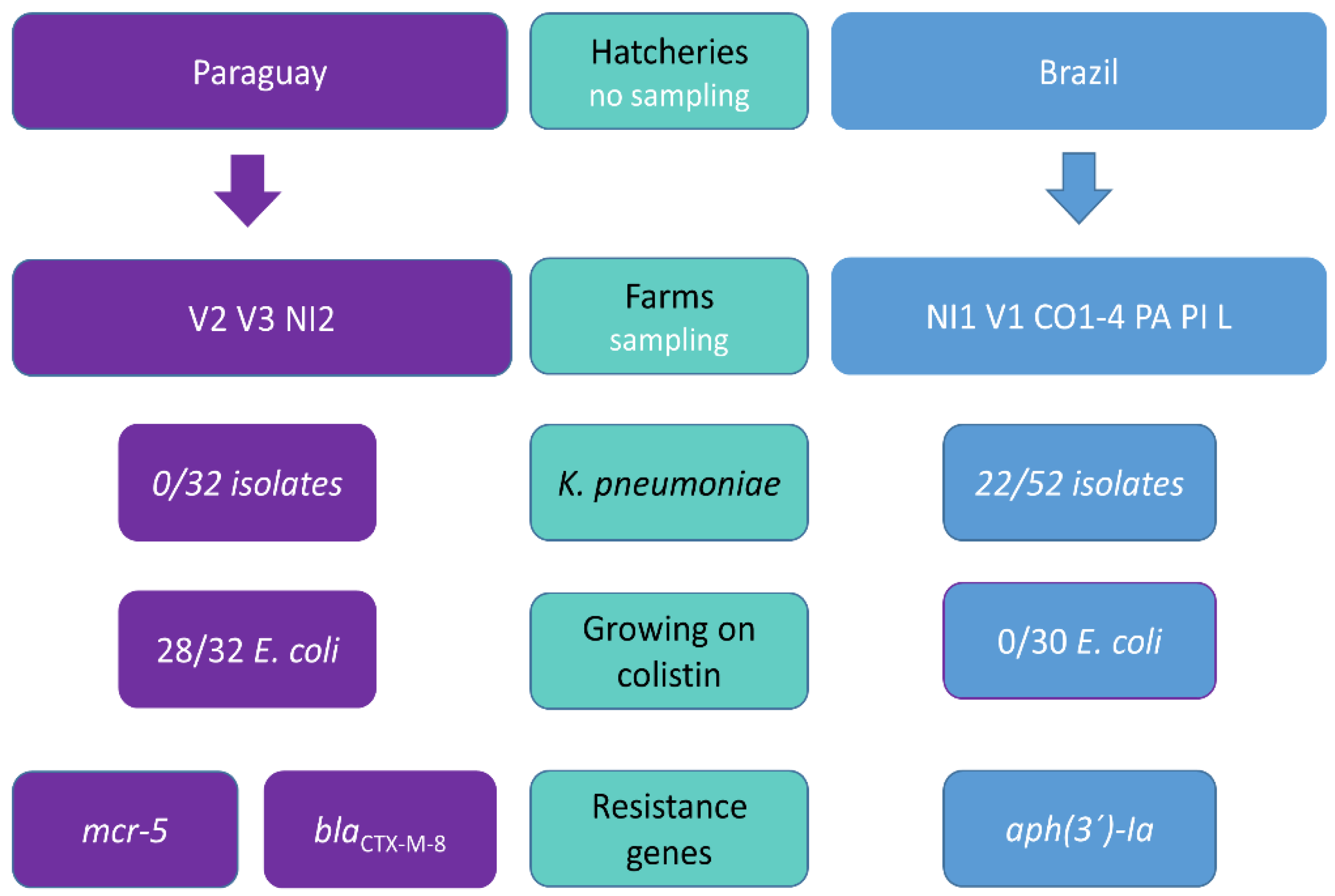
2.1. Samples Collection and Selective Cultivation
2.2. Antibiotic Susceptibility Testing
2.3. Clonality of K. pneumoniae Isolates and Testing of Selected Resistance Genes
2.4. Whole-Genome Sequencing and Data Analysis
2.5. Phylogenetic and Genomic Analysis
2.6. Transferability of ESBL/AmpC Genes
2.7. Typing of Plasmids Carrying ESBL/AmpC Genes
2.8. Plasmid Sequencing and Data Analysis
3. Results and Discussion
3.1. Trends in Antibiotic Resistance among Paraguayan Poultry
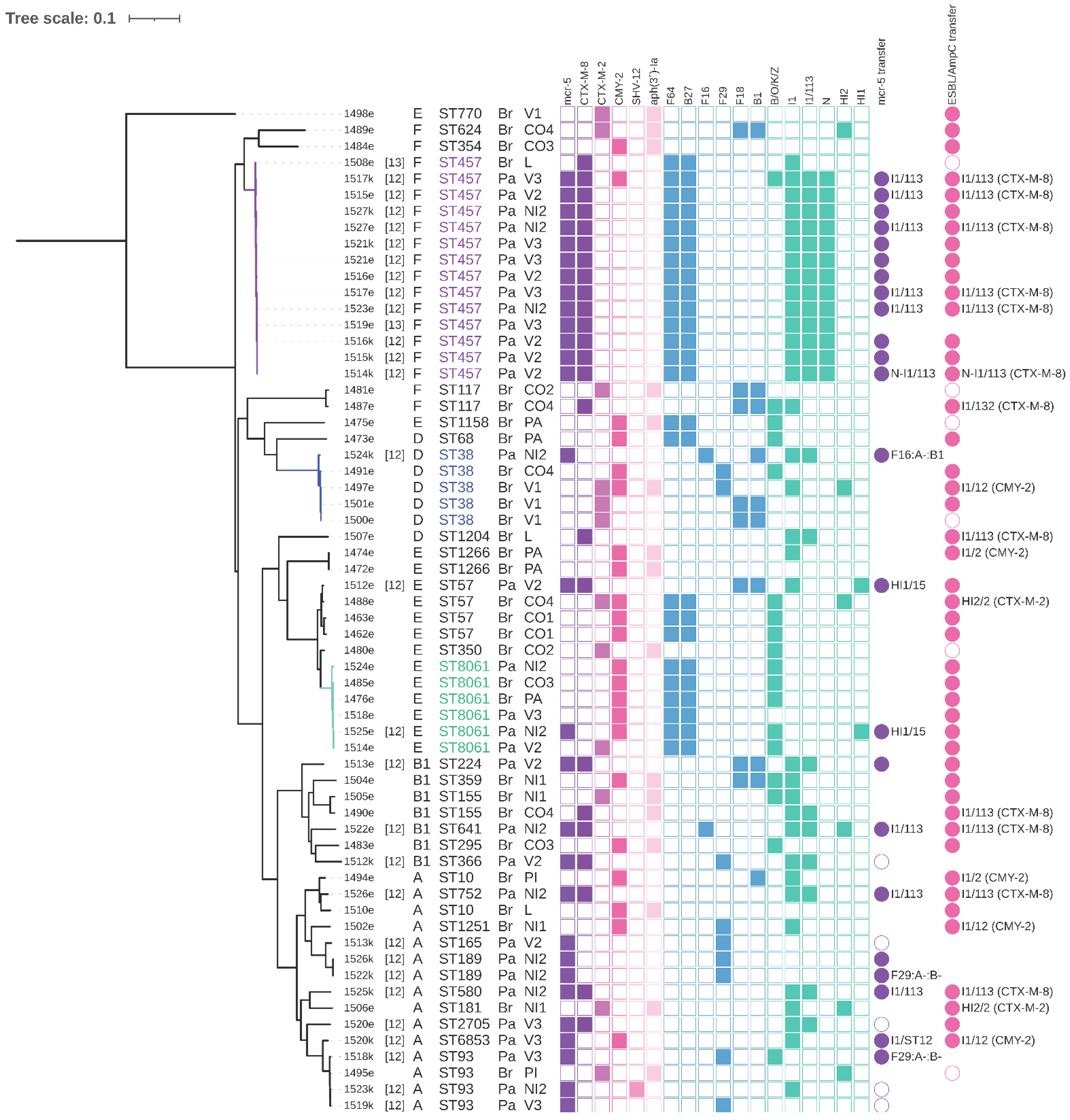
3.2. Phylogenetic Relationships among E. coli Isolates
3.3. E. coli ST457 from Paraguayan and Brazilian Hatcheries
3.4. E. coli ST38 Comparison
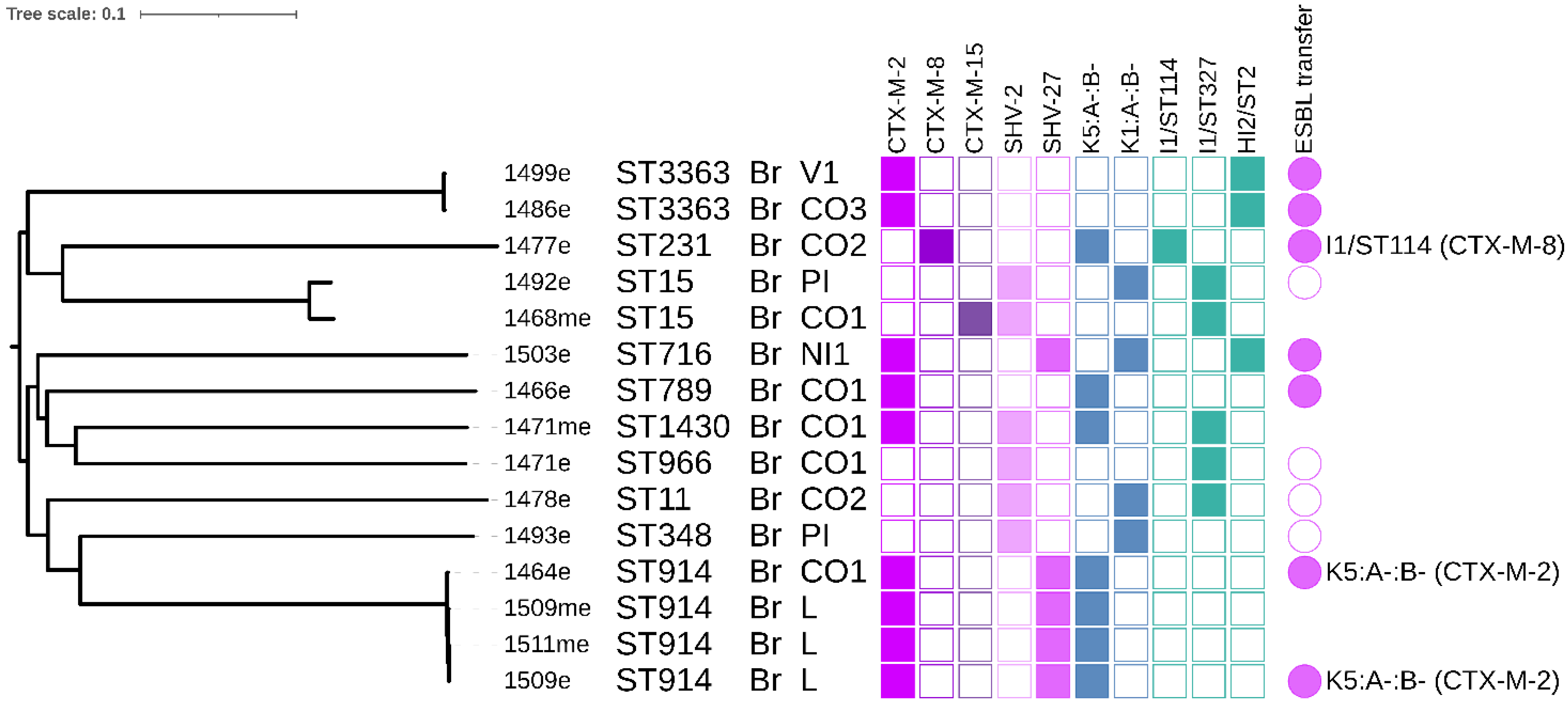
3.5. K. pneumoniae Phylogeny
3.6. Conjugation Experiments and Plasmid Typing
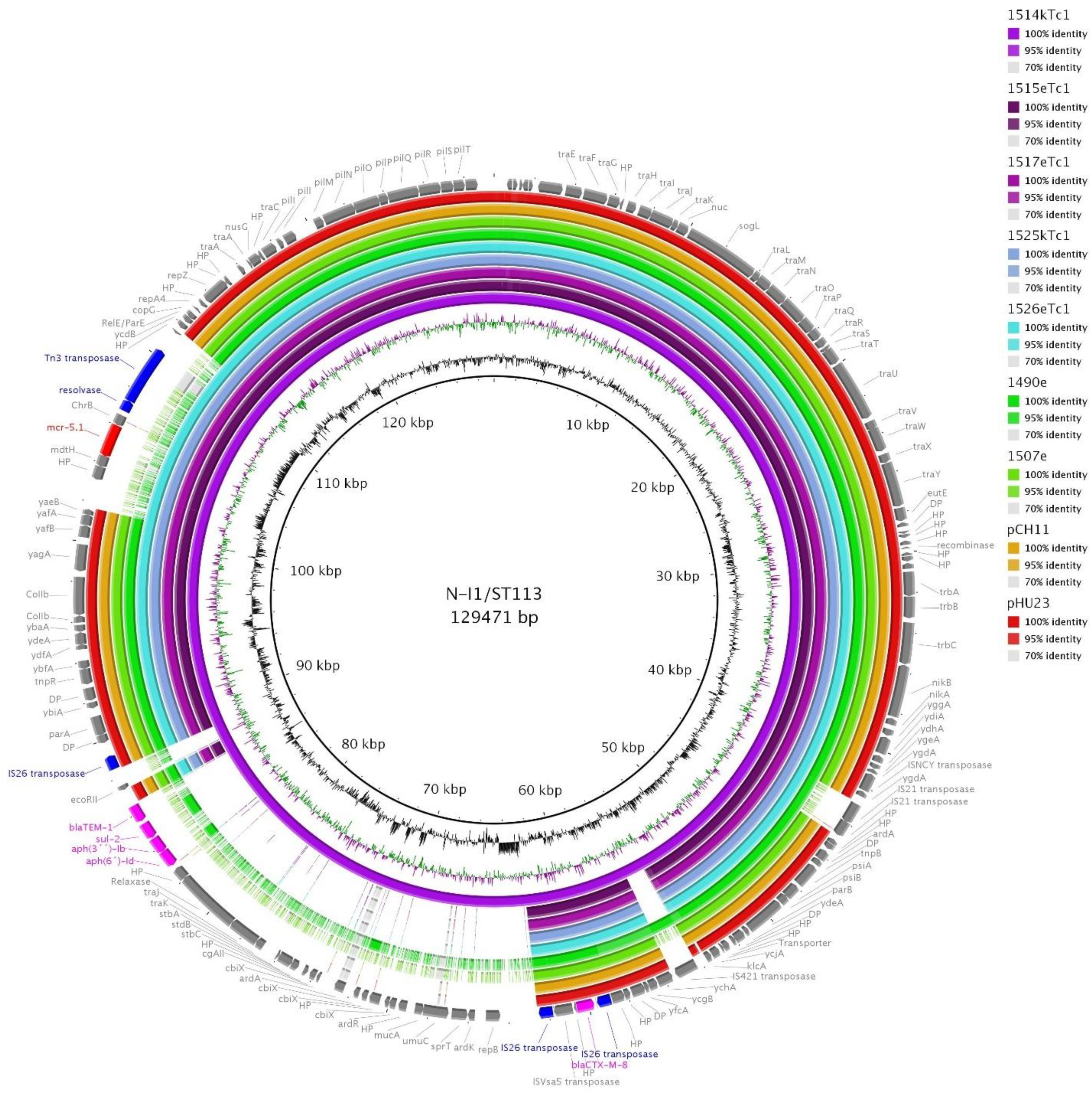
3.7. Detected Plasmid Replicons and mcr-5-Carrying Plasmids
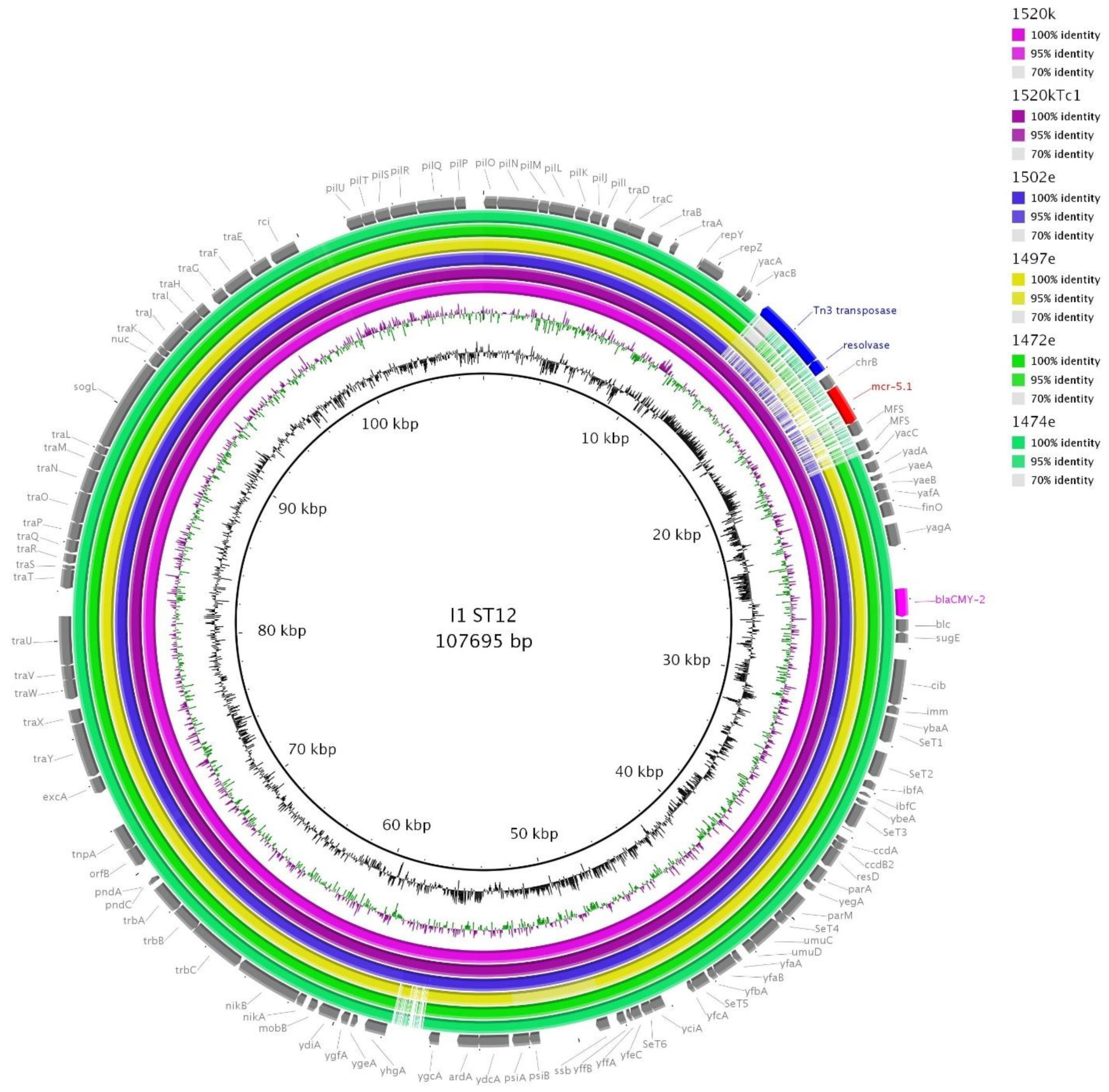
3.8. I1/ST113 and I1/ST12 Plasmids
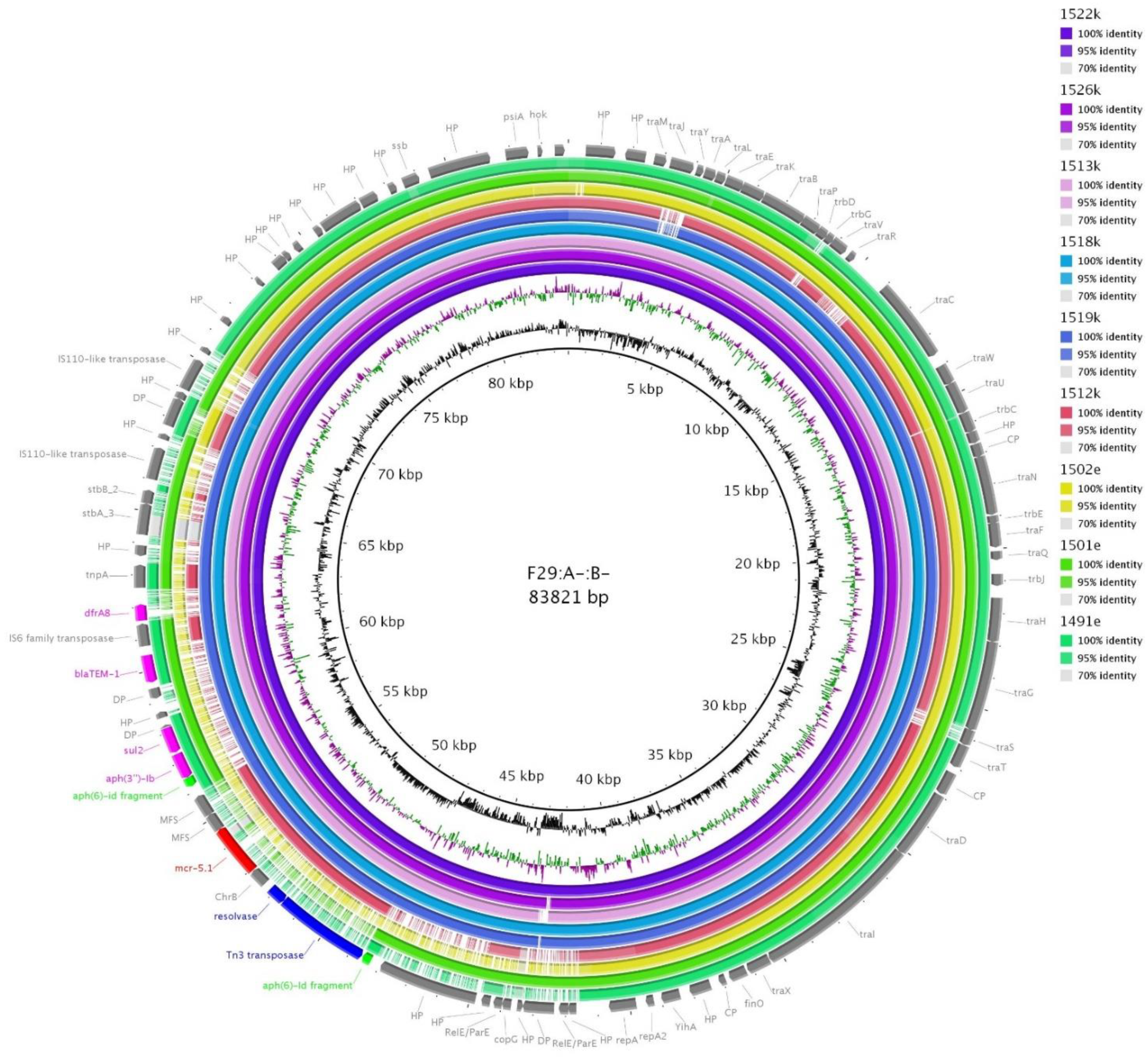
3.9. F64:A-:B27, F29:A-:B- and F16:A-:B1 Plasmids
3.10. HI1 ST15 Plasmids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ventola, C.L. The Antibiotic Resistance Crisis. Pharm. Ther. 2015, 40, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Aarestrup, F.M. Veterinary drug usage and antimicrobial resistance in bacteria of animal origin. Basic Clin. Pharmacol. Toxicol. 2005, 96, 271–281. [Google Scholar] [CrossRef]
- Carattoli, A. Animal reservoirs for extended spectrum β-lactamase producers. Clin. Microbiol. Infect. 2008, 14, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Leverstein-van Hall, M.A.; Dierikx, C.M.; Cohen Stuart, J.; Voets, G.M.; van den Munckhof, M.P.; van Essen-Zandbergen, A.; Platteel, T.; Fluit, A.C.; van de Sande-Bruinsma, N.; Scharinga, J.; et al. Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin. Microbiol. Infect. 2011, 17, 873–880. [Google Scholar] [CrossRef] [Green Version]
- Norizuki, C.; Wachino, J.I.; Suzuki, M.; Kawamura, K.; Nagano, N.; Kimura, K.; Arakawa, Y. Specific blaCTX-M-8/IncI1 Plasmid Transfer among Genetically Diverse Escherichia coli Isolates between Humans and Chickens. Antimicrob. Agent. Chemother. 2017, 61, e00663-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apostolakos, I.; Feudi, C.; Eichhorn, I.; Palmieri, N.; Fasolato, L.; Schwarz, S.; Piccirillo, A. High‑resolution characterisation of ESBL/pAmpC—producing Escherichia coli isolated from the broiler production pyramid. Sci. Rep. 2020, 10, 11123. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.C.; Penha Filho, R.A.C.; Neves Andrade, L.; Berchieri, A., Jr.; Costa Darini, A.N. Evaluation and characterization of plasmids carrying CTX-M genes in a non-clonal population of multidrug-resistant Enterobacteriaceae isolated from poultry in Brazil. Diagn. Microbiol. Infect. Dis. 2016, 85, 444–448. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Feng, Y.; Liu, L.; Wei, L.; Kang, M.; Zong, Z. Identification of novel mobile colistin resistance gene mcr-10. Emerg. Microbe. Infect. 2020, 9, 508–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiroga, C.; Nastro, M.; di Conza, J. Current scenario of plasmid-mediated colistin resistance in Latin America. Rev. Argent. Microbiol. 2019, 51, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.R.; Moura, Q.; Sartori, L.; Silva, K.C.; Cunha, M.P.; Esposito, F.; Lopes, R.; Otutumi, L.K.; Gonçalves, D.D.; Dropa, M.; et al. Silent dissemination of colistin-resistant Escherichia coli in South America could contribute to the global spread of the mcr-1 gene. Euro. Surveill. 2016, 21, 30214. [Google Scholar] [CrossRef] [PubMed]
- Dropa, M.; Balsalobre, L.C.; Lincopan, N.; Matté, G.R.; Matté, M.H. Complex class 1 integrons harboring CTX-M-2-encoding genes in clinical Enterobacteriaceae from a hospital in Brazil. J. Infect. Dev. Ctries. 2015, 9, 890–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesporova, K.; Jamborova, I.; Valcek, A.; Medvecky, M.; Literak, I.; Dolejska, M. Various conjugative plasmids carrying the mcr-5 gene in Escherichia coli isolates from healthy chickens in Paraguay. J. Antimicrob. Chemother. 2019, 74, 3394–3397. [Google Scholar] [CrossRef] [PubMed]
- Nesporova, K.; Wyrsch, E.R.; Valcek, A.; Bitar, I.; Chaw, K.; Harris, P.; Hrabak, J.; Literak, I.; Djordjevic, S.P.; Dolejska, M. Escherichia coli sequence type 457 is an emerging extended-spectrum-β-lactam-resistant lineage with reservoirs in wildlife and food-producing animals. Antimicrob. Agent. Chemother. 2021, 65, e01118-20. [Google Scholar] [CrossRef] [PubMed]
- Dobiasova, H.; Dolejska, M.; Jamborova, I.; Brhelova, E.; Blazkova, L.; Papousek, I.; Kozlova, M.; Klimes, J.; Cizek, A.; Literak, I. Extended spectrum beta-lactamase and fluoroquinolone resistance genes and plasmids among Escherichia coli isolates from zoo animals, Czech Republic. FEMS Microbiol. Ecol. 2013, 85, 604–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Perez, F.J.; Hanson, N.D. Detection of Plasmid-Mediated AmpC Beta-Lactamase Genes in Clinical Isolates by Using Multiplex PCR. JCM 2002, 157, 5336. [Google Scholar] [CrossRef]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated mcr-4 and mcr-5 for surveillance purposes. Euro. Surveill. 2018, 23, 17-00672. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2018, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 5, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Lykke Marvig, R.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Zankari, E.; Garciá-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In silico detection and typing of plasmids using plasmidfinder and plasmid multilocus sequence typing. Antimicrob. Agent. Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beghain, J.; Bridier-Nahmias, A.; le Nagard, H.; Denamur, E.; Clermont, O. ClermonTyping: An easy-to-use and accurate in silico method for Escherichia genus strain phylotyping. Microb. Genom. 2018, 4, e000192. [Google Scholar] [CrossRef] [PubMed]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the problem of comparing whole bacterial genomes across different sequencing platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A. Parkhill, Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darling, A.E.; Mau, B.; Perna, N.T. Progressive Mauve: Multiple Genome Alignment with Gene Gain, Loss and Rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Method. 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Alikhan, N.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [Green Version]
- Braykov, N.P.; Eisenberg, J.N.S.; Grossman, M.; Zhang, L.; Vasco, K.; Cevallos, W.; Muñoz, D.; Acevedo, A.; Moser, K.A.; Marrs, C.F.; et al. Antibiotic resistance in animal and environmental samples associated with small-scale poultry farming in northwestern Ecuador. mSphere 2016, 1, e00021-15. [Google Scholar] [CrossRef] [Green Version]
- Chauvin, C.; Devendec, L.; Jouy, E.; Cornec, L.; Francart, S.; Marois-Créhan, C.; Kempf, I. National Prevalence of Resistance to Third-Generation Cephalosporins in Escherichia coli Isolates from Layer Flocks in France. Antimicrob. Agent. Chemother. 2013, 57, 6351–6353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dame-Korevaar, A.; Fischer, E.A.J.; van der Goot, J.; Stegeman, A.; Mevius, D. Transmission routes of ESBL/pAmpC producing bacteria in the broiler production pyramid, a literature review. Prev. Vet. Med. 2019, 162, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Wakeham, D.; Brouwers, H.J.; Cobbold, R.N.; Abraham, S.; Mollinger, J.L.; Johnson, J.R.; Chapman, T.A.; Gordon, D.M.; Barrs, V.R.; et al. Human-associated fluoroquinolone-resistant Escherichia coli clonal lineages, including ST354, isolated from canine feces and extraintestinal infections in Australia. Microb. Infect. 2015, 17, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Zhuge, X.; Zhou, Z.; Jiang, M.; Wang, Z.; Sun, Y.; Tang, F.; Xue, F.; Ren, J.; Dai, J. Chicken-source Escherichia coli within phylogroup F shares virulence genotypes and is closely related to extraintestinal pathogenic E. coli causing human infections. Transbound. Emerg. Dis. 2020, 1–16. [Google Scholar] [CrossRef]
- Hung, W.T.; Cheng, M.F.; Tseng, F.C.; Chen, Y.S.; Lee, S.S.J.; Chang, T.H.; Lin, H.H.; Hung, C.H.; Wang, J.L. Bloodstream infection with extended-spectrum beta-lactamase-producing Escherichia coli: The role of virulence genes. J. Microbiol. Immunol. Infect. 2019, 52, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Maluta, R.P.; Logue, C.M.; Casas, M.R.T.; Meng, T.; Lopes Guastalli, E.A.; Galvão Rojas, T.C.; Montelli, A.C.; Sadatsune, T.; de Carvalho Ramos, M.; Nolan, K.L.; et al. Overlapped sequence types (STs) and serogroups of avian pathogenic (APEC) and human extra-intestinal pathogenic (ExPEC) Escherichia coli isolated in Brazil. PLoS ONE 2014, 9, e105016. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, M.P.N.; Fernandes, M.R.; Sellera, F.P.; Lopes, R.; Monte, D.F.; Hippólito, A.G.; Milanelo, L.; Raso, T.F.; Lincopan, N. International clones of extended-spectrum β -lactamase (CTX- M)-producing Escherichia coli in peri-urban wild animals, Brazil. Transbound. Emerg. Dis. 2020, 67, 1804–1815. [Google Scholar] [CrossRef] [Green Version]
- Hayer, S.S.; Lim, S.; Hong, S.; Elnekave, E.; Johnson, T.J.; Rovira, A.; Vannucci, F. Genetic Determinants of Resistance to Extended-Spectrum Cephalosporin and Fluoroquinolone in Escherichia coli Isolated from Diseased Pigs in the United States. mSphere 2020, 5, e00990-20. [Google Scholar] [CrossRef]
- Turton, J.F.; Doumith, M.; Hopkins, K.L.; Perry, C.; Meunier, D.; Woodford, N. Clonal expansion of Escherichia coli ST38 carrying a chromosomally integrated OXA-48 carbapenemase gene. J. Med. Microbiol. 2016, 65, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Guenther, S.; Semmler, T.; Stubbe, A.; Stubbe, M.; Wieler, L.H.; Schaufler, K. Chromosomally encoded ESBL genes in Escherichia coli of ST38 from Mongolian wild birds. J. Antimicrob. Chemother. 2017, 72, 310–1313. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Huang, M.; Zhang, X.; Yang, X.; Liu, Y.; Zhang, L. Microbial Pathogenesis Frequency of virulence factors in high biofilm formation blaKPC-2 producing Klebsiella pneumoniae strains from hospitals. Microb. Pathog. 2018, 116, 168–172. [Google Scholar] [CrossRef]
- Markovska, R.; Stoeva, T.; Boyanova, L.; Stankova, P.; Pencheva, D.; Keuleyan, E.; Murjeva, M.; Sredkova, M.; Ivanova, D.; Lazarova, G.; et al. Dissemination of successful international clone ST15 and clonal complex 17 among Bulgarian CTX-M-15 producing K. pneumoniae isolates. Diagn. Microbiol. Infect. Dis. 2017, 89, 310–313. [Google Scholar] [CrossRef]
- Ewers, C.; Stamm, I.; Pfeifer, Y.; Wieler, L.H.; Kopp, P.A.; Schønning, K.; Prenger-Berninghoff, E.; Scheufen, S.; Stolle, I.; Günther, S.; et al. Clonal spread of highly successful ST15-CTX-M-15 Klebsiella pneumoniae in companion animals and horses. J. Antimicrob. Chemother. 2014, 69, 2676–2680. [Google Scholar] [CrossRef] [Green Version]
- Lam, M.M.C.; Wyres, K.L.; Wick, R.R.; Judd, L.M.; Fostervold, A.; Holt, K.E.; Löhr, I.H. Convergence of virulence and MDR in a single plasmid vector in MDR Klebsiella pneumoniae ST15. J. Antimicrob. Chemother. 2019, 74, 1218–1222. [Google Scholar] [CrossRef] [Green Version]
- Palmeira, J.D.; Haenni, M.; Metayer, V.; Madec, J.Y.; Ferreira, M.H.N. Epidemic spread of IncI1/pST113 plasmid carrying the Extended-Spectrum Beta-Lactamase (ESBL) blaCTX-M-8 gene in Escherichia coli of Brazilian cattle. Vet. Microbiol. 2020, 243, 108629. [Google Scholar] [CrossRef]
- Melo, L.C.; Oresco, C.; Leigue, L.; Netto, H.M.; Melville, P.A.; Benites, N.R.; Saras, E.; Haenni, M.; Lincopan, N.; Madec, J.Y. Prevalence and molecular features of ESBL/pAmpC-producing Enterobacteriaceae in healthy and diseased companion animals in Brazil. Vet. Microbiol. 2018, 22, 59–66. [Google Scholar] [CrossRef]
- Eller, C.; Leistner, R.; Guerra, B.; Fischer, J.; Wendt, C.; Rabsch, W.; Werner, G.; Pfeifer, Y. Emergence of extended-spectrum β-lactamase (ESBL) CTX-M-8 in Germany. J. Antimicrob. Chemother. 2014, 69, 562–564. [Google Scholar] [CrossRef] [Green Version]
- Palmeira, J.D.; Ferreira, H.; Madec, J.Y.; Haenni, M. Pandemic Escherichia coli ST648 isolate harbouring fosA3 and blaCTX-M-8 on an IncI1/ST113 plasmid: A new successful combination for the spread of fosfomycin resistance? J. Glob. Antimicrob. Resist. 2018, 15, 254–255. [Google Scholar] [CrossRef] [PubMed]
- Roer, L.; Overballe-Petersen, S.; Hansen, F.; Johannesen, T.B.; Stegger, M.; Bortolaia, V.; Leekitcharoenphon, P.; Korsgaard, H.B.; Seyfarth, A.M.; Mossong, J.; et al. ST131 fimH22 Escherichia coli isolate with a blaCMY-2/IncI1/ST12 plasmid obtained from a patient with bloodstream infection: Highly similar to E. coli isolates of broiler origin. J. Antimicrob. Chemother. 2019, 74, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; Wagenaar, J.A.; Guerra, B.; Mevius, D.J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Approach | Plasmid | Size (kbp) | Antibiotic Resistance Genes |
|---|---|---|---|---|
| 1514k | PacBio-pDNA | N-I1/ST113 | 129.471 | mcr-5, blaCTX-M-8, blaTEM-1, aph(3´´)-Ib, aph(6)-Id, sul2 |
| 1524k | PacBio-pDNA | F16:A-:B1 | 142.664 | mcr-5, aadA2, sul1, tet(A), dfrA12 |
| 1520k | PacBio-Tc | I1/ST12 | 107.695 | mcr-5, blaCMY-2 |
| 1512e | PacBio-Tc | HI1/ST15 | 197.568 | mcr-5, lnu(G), tet(B), sul2 |
| 1525e | PacBio-Tc | HI1/ST15 | 209.411 | mcr-5, blaTEM-1, aph(3´´)-Ib, aph(6)-Id, lnu(G), tet(B), sul2 |
| 1522k | Illumina + PCR | F29:A-:B- | 83.821 | mcr-5, aph(3´´)-Ib, blaTEM-1, sul2, dfrA8 |
| 1517k | PacBio-WGS | F64:A-:B27 | 167.074 | - |
| 1517k | PacBio-WGS | K2 | 86.723 | blaCMY-2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nesporova, K.; Valcek, A.; Papagiannitsis, C.; Kutilova, I.; Jamborova, I.; Davidova-Gerzova, L.; Bitar, I.; Hrabak, J.; Literak, I.; Dolejska, M. Multi-Drug Resistant Plasmids with ESBL/AmpC and mcr-5.1 in Paraguayan Poultry Farms: The Linkage of Antibiotic Resistance and Hatcheries. Microorganisms 2021, 9, 866. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040866
Nesporova K, Valcek A, Papagiannitsis C, Kutilova I, Jamborova I, Davidova-Gerzova L, Bitar I, Hrabak J, Literak I, Dolejska M. Multi-Drug Resistant Plasmids with ESBL/AmpC and mcr-5.1 in Paraguayan Poultry Farms: The Linkage of Antibiotic Resistance and Hatcheries. Microorganisms. 2021; 9(4):866. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040866
Chicago/Turabian StyleNesporova, Kristina, Adam Valcek, Costas Papagiannitsis, Iva Kutilova, Ivana Jamborova, Lenka Davidova-Gerzova, Ibrahim Bitar, Jaroslav Hrabak, Ivan Literak, and Monika Dolejska. 2021. "Multi-Drug Resistant Plasmids with ESBL/AmpC and mcr-5.1 in Paraguayan Poultry Farms: The Linkage of Antibiotic Resistance and Hatcheries" Microorganisms 9, no. 4: 866. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040866