
Neospora caninum and/or Toxoplasma gondii Seroprevalence: Vaccination against PCV2 and Muscle Enzyme Activity in Seropositive and Seronegative Pigs

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Inclusion Criteria
2.2.1. Farms
- Minimum capacity of 50 sows.
- Operation type of exclusive farrow-to-finish farms.
- Vaccination of gilts/sows against Aujeszky’s disease virus, parvovirus, atrophic rhinitis, erysipelas, Porcine Reproductive and Respiratory Virus (PRRSV), Escherichia coli, and Clostridium infections. Vaccination of weaners against Mycoplasma hyopneumoniae.
- Regular control of endo/ectoparasites by administration of ivermectin in sows.
- Single housing systems of sows with farrowing crate, including cast iron slat and solid plastic slat.
- Implementation of environmental conditions and stocking density in compliance with swine welfare requirements (Council Directive 2008/120/EC of 18 December 2008).
- Implementation of good biosecurity practices to reduce the likelihood of disease introduction and/or spread.
- Balanced diet (essential amino acids, minerals, and vitamins) according to National Research Council [37].
- Regular use of toxin binders in the feed of sows during gestation and lactation period and in the feed of weaning pigs.
2.2.2. Animals
2.3. Sampling
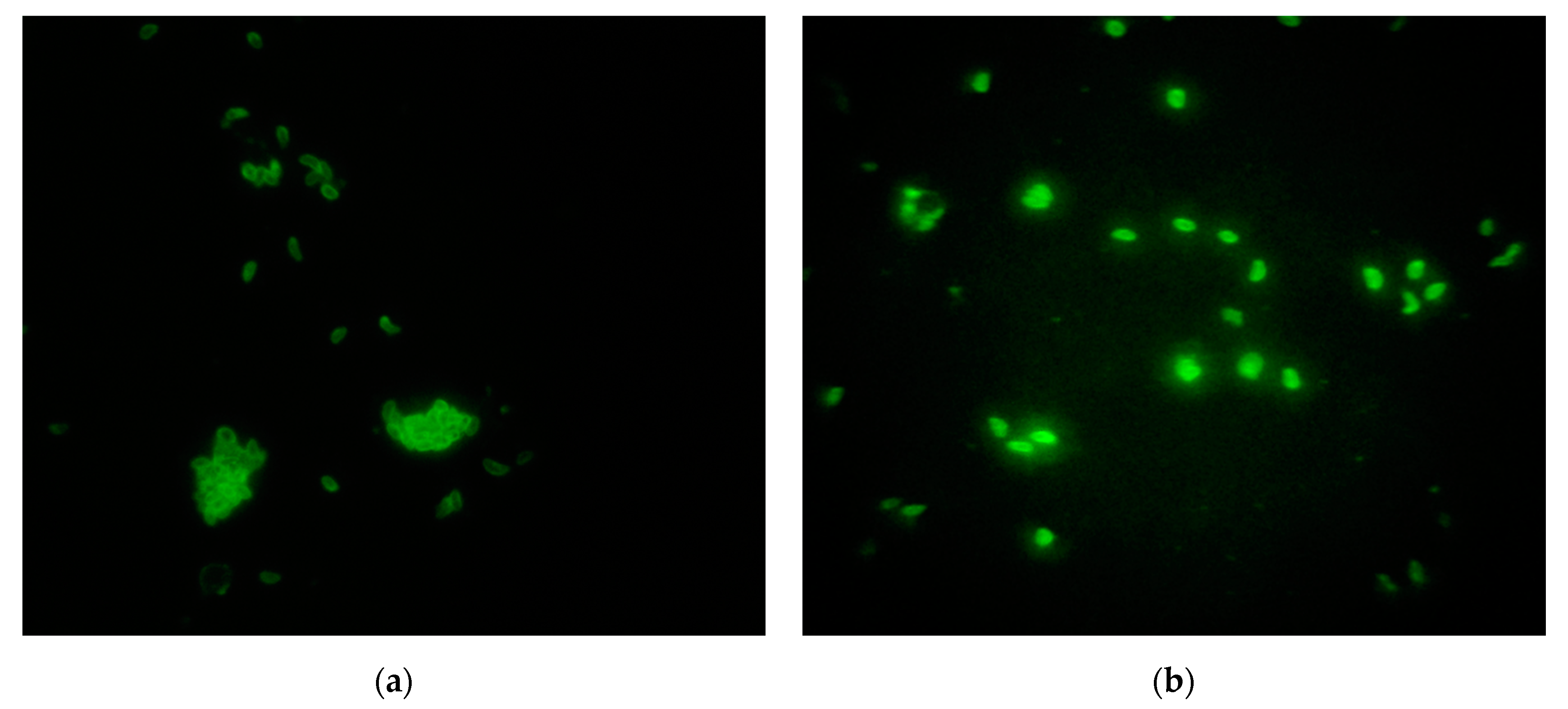
2.4. Indirect Immunofluorescence Antibody (IFA) Assay
2.5. Biochemical Analysis
2.6. Data Analysis
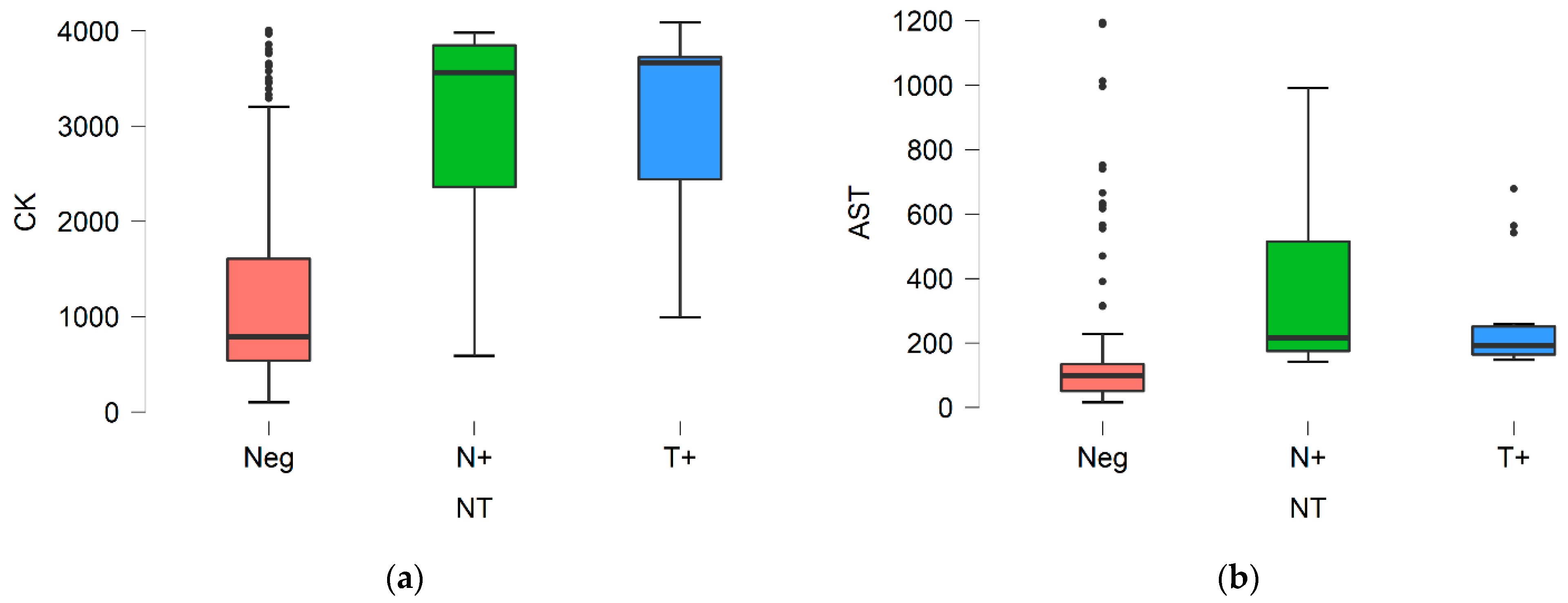
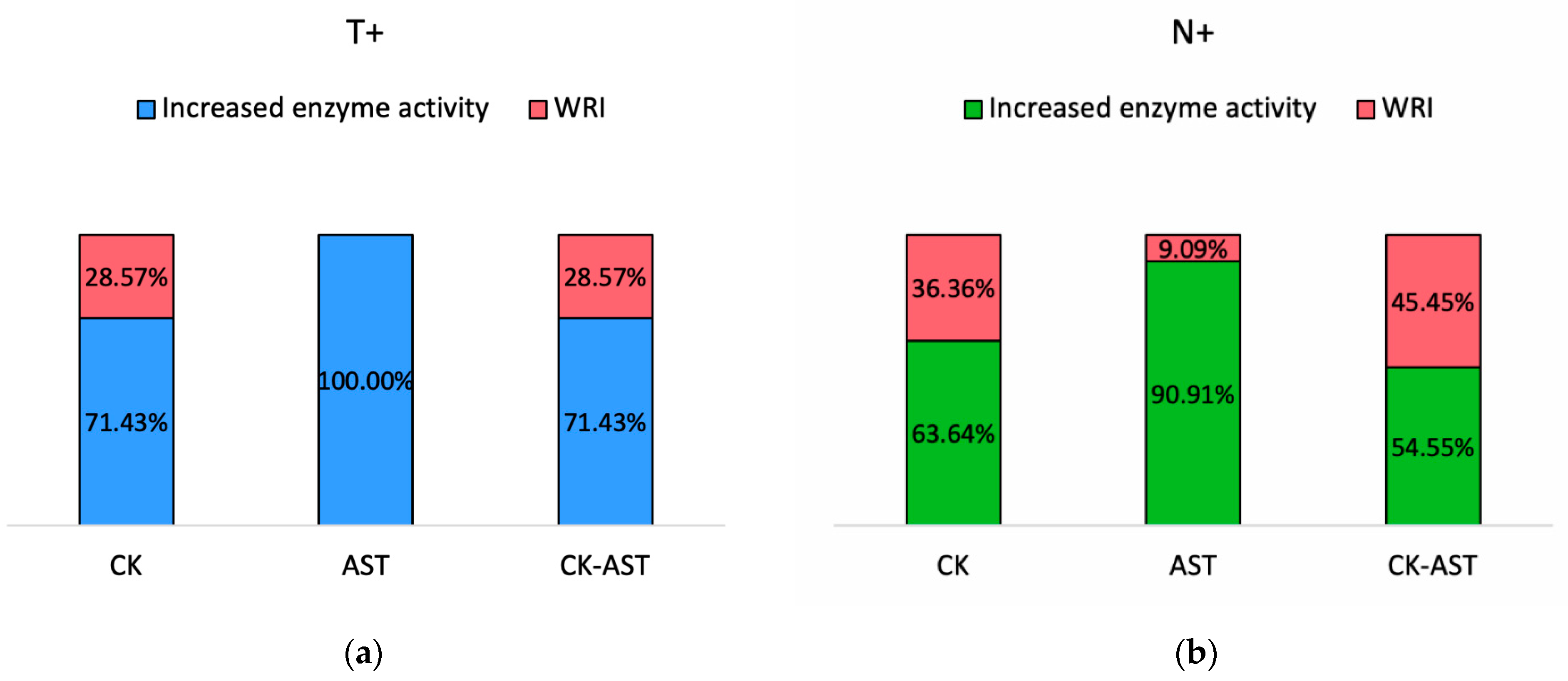
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Berman, J.J. Chapter 19—Apicomplexa. In Taxonomic Guide to Infectious Diseases; Berman, J.J., Ed.; Academic Press: Boston, MA, USA, 2012; pp. 103–109. [Google Scholar] [CrossRef]
- Khan, A.; Shaik, J.S.; Sikorski, P.; Dubey, J.P.; Grigg, M.E. Neosporosis: An Overview of Its Molecular Epidemiology and Pathogenesis. Engineering 2020, 6, 10–19. [Google Scholar] [CrossRef]
- Hill, D.; Dubey, J.P. Toxoplasma gondii: Transmission, diagnosis and prevention. Clin. Microbiol. Infect. 2002, 8, 634–640. [Google Scholar] [CrossRef] [Green Version]
- Flegr, J.; Prandota, J.; Sovickova, M.; Israili, Z.H. Toxoplasmosis—A global threat. Correlation of latent toxoplasmosis with specific disease burden in a set of 88 countries. PLoS ONE 2014, 9, e90203. [Google Scholar] [CrossRef] [Green Version]
- WHO. Toxoplasmosis Fact Sheet. Available online: https://www.euro.who.int/__data/assets/pdf_file/0011/294599/Factsheet-Toxoplasmosis-en.pdf?ua=1 (accessed on 19 February 2021).
- Duarte, P.O.; Oshiro, L.M.; Zimmermann, N.P.; Csordas, B.G.; Dourado, D.M.; Barros, J.C.; Andreotti, R. Serological and molecular detection of Neospora caninum and Toxoplasma gondii in human umbilical cord blood and placental tissue samples. Sci. Rep. 2020, 10, 9043. [Google Scholar] [CrossRef]
- Foroutan, M.; Fakhri, Y.; Riahi, S.M.; Ebrahimpour, S.; Namroodi, S.; Taghipour, A.; Spotin, A.; Gamble, H.R.; Rostami, A. The global seroprevalence of Toxoplasma gondii in pigs: A systematic review and meta-analysis. Vet. Parasitol. 2019, 269, 42–52. [Google Scholar] [CrossRef]
- Dubey, J.P.; Gamble, H.R.; Hill, D.; Sreekumar, C.; Romand, S.; Thulliez, P. High Prevalence Of Viable Toxoplasma Gondii Infection In Market Weight Pigs From A Farm In Massachusetts. J. Parasitol. 2002, 88, 1234–1238. [Google Scholar] [CrossRef]
- Damriyasa, I.M.; Bauer, C.; Edelhofer, R.; Failing, K.; Lind, P.; Petersen, E.; Schares, G.; Tenter, A.M.; Volmer, R.; Zahner, H. Cross-sectional survey in pig breeding farms in Hesse, Germany: Seroprevalence and risk factors of infections with Toxoplasma gondii, Sarcocystis spp. and Neospora caninum in sows. Vet. Parasitol. 2004, 126, 271–286. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, S.S.; Pena, H.F.; Alves, C.J.; Guimarães Filho, A.A.; Oliveira, R.M.; Maksimov, P.; Schares, G.; Gennari, S.M. Prevalence of anti-Toxoplasma gondii and anti-Neospora caninum antibodies in swine from Northeastern Brazil. Rev. Bras. Parasitol. Vet. 2010, 19, 80–84. [Google Scholar] [CrossRef] [Green Version]
- Feitosa, T.F.; Vilela, V.L.; de Melo, L.R.; de Almeida Neto, J.L.; Souto, D.V.; de Morais, D.F.; Athayde, A.C.; Azevedo, S.S.; Pena, H.F. Toxoplasma gondii and Neospora caninum in slaughtered pigs from Northeast, Brazil. Vet. Parasitol. 2014, 202, 305–309. [Google Scholar] [CrossRef]
- Minetto, M.K.; Witter, R.; Oliveira, A.C.S.; Minetto, J.A.; Barros, M.L.; Aguiar, D.M.; Pacheco, R.C. Antibodies anti-Toxoplasma gondii and anti-Neospora caninum in backyard pigs from the state of Mato Grosso, Brazil. Rev. Bras. Parasitol. Vet. 2019, 28, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Gui, B.Z.; Lv, Q.Y.; Ge, M.; Li, R.C.; Zhu, X.Q.; Liu, G.H. First report of Neospora caninum infection in pigs in China. Transbound. Emerg. Dis. 2020, 67, 29–32. [Google Scholar] [CrossRef]
- Bártová, E.; Sedlák, K. Seroprevalence of Toxoplasma gondii and Neospora caninum in slaughtered pigs in the Czech Republic. Parasitology 2011, 138, 1369–1371. [Google Scholar] [CrossRef]
- Bevins, S.; Blizzard, E.; Bazan, L.; Whitley, P. Neospora caninum exposure in overlapping populations of coyotes (Canis latrans) and feral swine (Sus scrofa). J. Wildl. Dis. 2013, 49, 1028–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefkaditis, M.; Spanoudis, K.; Tsakiroglou, M.; Panorias, A.; Sossidou, A. Seroprevalence of Neospora caninum infection in stray dogs in Chalkidiki, Northern Greece. J. Hell. Vet. Med Soc. 2021, 71, 2511–2514. [Google Scholar] [CrossRef]
- Kouam, M.; Kantzoura, V.; Cabezón, O.; Nogareda, C.; Almería, S.; Theodoropoulos, G. Comparative cross-sectional study of Neospora caninum and Toxoplasma gondii: Seroprevalence in sheep of Greece and North-Eastern Spain. Sustain. Dev. Cult. Tradit. J. 2019, 1–7. [Google Scholar] [CrossRef]
- Diakou, A.; Papadopoulos, E.; Panousis, N.; Charilaos, K.; Giadinis, N. Toxoplasma gondii and Neospora caninum seroprevalence in dairy sheep and goats mixed stock farming. Vet. Parasitol. 2013, 198. [Google Scholar] [CrossRef]
- Lefkaditis, M.; Mpairamoglou, R.; Sossidou, A.; Spanoudis, K.; Tsakiroglou, M. Neospora caninum, A potential cause of reproductive failure in dairy cows from Northern Greece. Vet. Parasitol. Reg. Stud. Rep. 2020, 19, 100365. [Google Scholar] [CrossRef]
- Lefkaditis, M.; Evagelopoulou, G.; Sossidou, A.; Spanoudis, K. Neosporosis and toxoplasmosis are two prevalent and important protozoses in dairy cows in small farms from Thessaly, Central Greece. J. Hell. Vet. Med Soc. 2020, 71, 2357–2362. [Google Scholar] [CrossRef]
- Touloudi, A.; Valiakos, G.; Athanasiou, L.V.; Birtsas, P.; Giannakopoulos, A.; Papaspyropoulos, K.; Kalaitzis, C.; Sokos, C.; Tsokana, C.N.; Spyrou, V.; et al. A serosurvey for selected pathogens in Greek European wild boar. Vet. Rec. Open 2015, 2, e000077. [Google Scholar] [CrossRef] [Green Version]
- Tsokana, C.N.; Sokos, C.; Giannakopoulos, A.; Birtsas, P.; Athanasiou, L.V.; Valiakos, G.; Sofia, M.; Chatzopoulos, D.C.; Kantere, M.; Spyrou, V.; et al. Serological and molecular investigation of selected parasitic pathogens in European brown hare (Lepus europaeus) in Greece: Inferring the ecological niche of Toxoplasma gondii and Leishmania infantum in hares. Parasitol. Res. 2019, 118, 2715–2721. [Google Scholar] [CrossRef]
- Diza, E.; Frantzidou, F.; Souliou, E.; Arvanitidou, M.; Gioula, G.; Antoniadis, A. Seroprevalence of Toxoplasma gondii in northern Greece during the last 20 years. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2005, 11, 719–723. [Google Scholar] [CrossRef] [Green Version]
- Tzanidakis, N.; Maksimov, P.; Conraths, F.; Kiossis, E.; Brozos, C.; Sotiraki, S.; Schares, G. Toxoplasma gondii in sheep and goats: Seroprevalence and potential risk factors under dairy husbandry practices. Vet. Parasitol. 2012, 190. [Google Scholar] [CrossRef]
- Papatsiros, V.G.; Athanasiou, L.V.; Stougiou, D.; Papadopoulos, E.; Maragkakis, G.G.; Katsoulos, P.D.; Lefkaditis, M.; Kantas, D.; Tzika, E.D.; Tassis, P.D.; et al. Cross-Sectional Serosurvey and Risk Factors Associated with the Presence of Toxoplasma gondii Antibodies in Pigs in Greece. Vector Borne Zoonotic Dis. 2016, 16, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Kouam, M.K.; Diakou, A.; Kanzoura, V.; Papadopoulos, E.; Gajadhar, A.A.; Theodoropoulos, G. A seroepidemiological study of exposure to Toxoplasma, Leishmania, Echinococcus and Trichinella in equids in Greece and analysis of risk factors. Vet. Parasitol. 2010, 170, 170–175. [Google Scholar] [CrossRef]
- Webster, J.P.; Dubey, J.P. Toxoplasmosis of Animals and Humans. Parasites Vectors 2010, 3, 112. [Google Scholar] [CrossRef] [Green Version]
- Lobato, J.; Silva, D.A.O.; Mineo, T.W.P.; Amaral, J.D.H.F.; Segundo, G.R.S.; Costa-Cruz, J.M.; Ferreira, M.S.; Borges, A.S.; Mineo, J.R. Detection of immunoglobulin G antibodies to Neospora caninum in humans: High seropositivity rates in patients who are infected by human immunodeficiency virus or have neurological disorders. Clin. Vaccine Immunol. 2006, 13, 84–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paspalaki, P.K.; Mihailidou, E.P.; Bitsori, M.; Tsagkaraki, D.; Mantzouranis, E. Polyomyositis and myocarditis associated with acquired toxoplasmosis in an immunocompetent girl. BMC Musculoskelet. Disord. 2001, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Hassene, A.; Vital, A.; Anghel, A.; Guez, S.; Series, C. Acute acquired toxoplasmosis presenting as polymyositis and chorioretinitis in immunocompetent patient. Jt. Bone Spine 2008, 75, 603–605. [Google Scholar] [CrossRef] [PubMed]
- Cuomo, G.; D’Abrosca, V.; Rizzo, V.; Nardiello, S.; La Montagna, G.; Gaeta, G.B.; Valentini, G. Severe polymyositis due to Toxoplasma gondii in an adult immunocompetent patient: A case report and review of the literature. Infection 2013, 41, 859–862. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.; Levesque, D.; Shelton, G.D. Canine Inflammatory Myopathies: A Clinicopathologic Review of 200 Cases. J. Vet. Intern. Med. 2004, 18, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Butts, D.R.; Langley-Hobbs, S.J. Lameness, generalised myopathy and myalgia in an adult cat with toxoplasmosis. J. Feline Med. Surg. Open Rep. 2020, 6, 2055116920909668. [Google Scholar] [CrossRef]
- Dubey, J.P.; Schlafer, D.H.; Urban, J.F., Jr.; Lindsay, D.S. Lesions in fetal pigs with transplacentally-induced toxoplasmosis. Vet. Pathol. 1990, 27, 411–418. [Google Scholar] [CrossRef]
- Dubey, J.P. A review of toxoplasmosis in pigs. Vet. Parasitol. 1986, 19, 181–223. [Google Scholar] [CrossRef]
- Snak, A.; Henrique, S.M.; Sebolt, A.P.R.; Cristani, J.; Sato, M.E.; Miletti, L.C.; de Moura, A.B. Experimental infection of tachyzoites of the NC1 strain of Neosporacaninum in female swine. Parasitol. Res. 2021, 120, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- National-Research-Council. Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar] [CrossRef] [Green Version]
- Basso, W.; Grimm, F.; Ruetten, M.; Djokic, V.; Blaga, R.; Sidler, X.; Deplazes, P. Experimental Toxoplasma gondii infections in pigs: Humoral immune response, estimation of specific IgG avidity and the challenges of reproducing vertical transmission in sows. Vet. Parasitol. 2017, 236, 76–85. [Google Scholar] [CrossRef]
- OIE. Chapter 2.9.10.—Toxoplasmosis. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/maj2011/2.09.10_TOXO.pdf (accessed on 15 May 2021).
- Jamieson, D.J.; Theiler, R.N.; Rasmussen, S.A. Emerging infections and pregnancy. Emerg. Infect. Dis. 2006, 12, 1638–1643. [Google Scholar] [CrossRef] [PubMed]
- Bjorkman, C.; Alenius, S.; Manuelsson, U.; Uggla, A. Neospora caninum and Bovine Virus Diarrhoea Virus Infections in Swedish Dairy Cows in Relation to Abortion. Vet. J. 2000, 159, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.E.M.; Fernandes, F.; Monteiro, F.L.; Braunig, P.; Cargnelutti, J.F.; Flores, E.F.; Weiblen, R.; Vogel, F.S. Co-infection by Neopora caninum and bovine viral diarrhea virus in cattle from Rio Grande do Sul, Brazil, destined to exportation. Pesq. Vet. Bras. 2020. [Google Scholar] [CrossRef]
- Segalés, J.; Domingo, M.; Chianini, F.; Majó, N.; Domínguez, J.; Darwich, L.; Mateu, E. Immunosuppression in postweaning multisystemic wasting syndrome affected pigs. Vet. Microbiol. 2004, 98, 151–158. [Google Scholar] [CrossRef]
- Liu, S.; Li, Q.; Qiao, J.; Wang, J.; Cui, D.; Gu, K.; Zhou, S.; Li, H. Endothelial IL-8 induced by porcine circovirus type 2 affects dendritic cell maturation and antigen-presenting function. Virol. J. 2019, 16, 154. [Google Scholar] [CrossRef]
- Dubey, J.P.; Lappin, M.R. Toxoplasmosis and neosporosis. In Infectious Diseases of Dogs and Cats; Greene, C.E., Ed.; Saunders-Elsevier: St. Louis, MO, USA, 2005; pp. 754–775. [Google Scholar]
- Jin, R.M.; Blair, S.J.; Warunek, J.; Heffner, R.R.; Blader, I.J.; Wohlfert, E.A. Regulatory T Cells Promote Myositis and Muscle Damage in Toxoplasma gondii Infection. J. Immunol. 2017, 198, 352–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grindem, C.B.; Neel, J.A.; Escobar, C. Evaluation of Skeletal Muscle Function and Injury. In The Clinical Chemistry of Laboratory Animals, 3rd ed.; Kurtz, D.M., Travlos, G.S., Eds.; Taylor & Francis: Boca Raton, FL USA, 2018; pp. 475–514. [Google Scholar]
- Stockham, S.L.; Scott, M.A. Enzymes. In Fundamentals of Veterinary Clinical Pathology, 2nd ed.; Wiley-Blackwell: Ames, IA, USA, 2008; pp. 640–669. [Google Scholar]
- Boyd, J.W. The mechanisms relating to increases in plasma enzymes and isoenzymes in diseases of animals. Vet. Clin. Pathol. 1983, 12, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Brancaccio, P.; Maffulli, N.; Buonauro, R.; Limongelli, F.M. Serum enzyme monitoring in sports medicine. Clin. Sports Med. 2008, 27, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Lucas, V.; Barrera, R.; Duque, F.J.; Ruiz, P.; Zaragoza, C. Effect of exercise on serum markers of muscle inflammation in Spanish Greyhounds. Am. J. Vet. Res. 2015, 76, 637–643. [Google Scholar] [CrossRef]
- Doize, F.; Laporte, R.; DeRoth, L. Effects of exercise on skeletal muscle and serum enzyme activities in pigs. Vet. Res. Commun. 1989, 13, 341–347. [Google Scholar] [CrossRef]
- Szumilak, D.; Sułowicz, W.; Walatek, B. Rhabdomyolysis: Clinical features, causes, complications and treatment. Przegl. Lek. 1998, 55, 274–279. [Google Scholar]
- Mokuno, K.; Riku, S.; Sugimura, K.; Takahashi, A.; Kato, K.; Osugi, S. Serum creatine kinase isoenzymes in Duchenne muscular dystrophy determined by sensitive enzyme immunoassay methods. Muscle Nerve 1987, 10, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Athanasiou, L.V.; Polizopoulou, Z.S.; Papaioannou, N.; Roubies, N.; Bodina, E.S. Effects of injection volume and excipient on muscle enzyme activity and post injection muscle damage. In Proceedings of the congress of the International Society of Animal Clinical Biochemistry, Instabul, Turkey, 1 May 2006; p. 93. [Google Scholar]
- Gudmundson, J. The clinicopathological findings of mulberry heart disease in a piglet. Can. Vet. J. 1976, 17, 45–47. [Google Scholar]
- Loynachan, A.T. Cardiovascular and Hematopoietic Systems. In Diseases of Swine, 10th ed.; John Wiley & Sons, Inc.: West Sussex, UK, 2012; pp. 189–198. [Google Scholar]
- Done, S.; Williamson, S.M.; Strugnell, B.W. Porcine Stress Syndrome. In Diseases of Swine, 10th ed.; John Wiley & Sons, Inc.: West Sussex, UK, 2012; pp. 314–315. [Google Scholar]
- Thorén-Tolling, K.; Jönsson, L. Creatine kinase isoenzymes in serum of pigs having myocardial and skeletal muscle necrosis. Can. J. Comp. Med. 1983, 47, 207–216. [Google Scholar]
- Webb, A.J.; Jordan, C.H.C. Halothane sensitivity as a field test for stress-susceptibility in the pig. Anim. Sci. 1978, 26, 157–168. [Google Scholar] [CrossRef]
- Konradt, G.; Cruz, R.A.S.; Bassuino, D.M.; Bianchi, M.V.; de Andrade, C.P.; da Silva, F.S.; Driemeier, D.; Pavarini, S.P. Granulomatous Necrotizing Myositis in Swine Affected by Porcine Circovirus Disease. Vet. Pathol. 2018, 55, 268–272. [Google Scholar] [CrossRef]
- Opriessnig, T.; Janke, B.H.; Halbur, P.G. Cardiovascular lesions in pigs naturally or experimentally infected with porcine circovirus type 2. J. Comp. Pathol. 2006, 134, 105–110. [Google Scholar] [CrossRef]
- Wu, Z.; Sofronic-Milosavljevic, L.; Nagano, I.; Takahashi, Y. Trichinella spiralis: Nurse cell formation with emphasis on analogy to muscle cell repair. Parasites Vectors 2008, 1, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Giessen, J.; Deksne, G.; Gómez-Morales, M.A.; Troell, K.; Gomes, J.; Sotiraki, S.; Rozycki, M.; Kucsera, I.; Djurković-Djaković, O.; Robertson, L.J. Surveillance of foodborne parasitic diseases in Europe in a One Health approach. Parasite Epidemiol. Control 2021, 13, e00205. [Google Scholar] [CrossRef] [PubMed]
- Boutsini, S.; Papatsiros, V.G.; Stougiou, D.; Marucci, G.; Liandris, E.; Athanasiou, L.V.; Papadoudis, A.; Karagiozopoulos, E.; Bisias, A.; Pozio, E. Emerging Trichinella britovi infections in free ranging pigs of Greece. Vet. Parasitol. 2014, 199, 278–282. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Farm Size 1 | Farms | Blood Samples |
|---|---|---|
| 50–100 | 6 | 30 |
| 101–250 | 10 | 100 |
| 251–500 | 10 | 150 |
| >500 | 5 | 100 |
| Farm Size 1 | PCV2 Unvaccinated Farms/Total Farms | Blood Samples from Unvaccinated Farms/Total Blood Samples |
|---|---|---|
| 50–100 | 5/6 | 25/30 |
| 101–250 | 2/10 | 20/100 |
| 251–500 | 0/10 | 0/150 |
| >500 | 0/5 | 0/100 |
| PCV2 Vaccination Status | Seropositive Pigs T. gondii (16) N. caninum (13) | |||
|---|---|---|---|---|
| N | % | N | % | |
| Vaccinated (322) | 9 | 2.79 1 | 1 | 0.31 1 |
| Unvaccinated (42) | 7 | 16.67 2 | 12 | 28.57 2 |
| Parity | Seropositive Pigs T. gondii (16) N. caninum (13) | |||
|---|---|---|---|---|
| N | % | N | % | |
| 1st parity (143) | 6 | 4.2 | 7 | 4.9 |
| 2nd parity (221) | 10 | 4.5 | 6 | 2.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Athanasiou, L.V.; Papatsiros, V.G.; Spanou, V.M.; Katsogiannou, E.G.; Dedousi, A. Neospora caninum and/or Toxoplasma gondii Seroprevalence: Vaccination against PCV2 and Muscle Enzyme Activity in Seropositive and Seronegative Pigs. Microorganisms 2021, 9, 1097. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051097
Athanasiou LV, Papatsiros VG, Spanou VM, Katsogiannou EG, Dedousi A. Neospora caninum and/or Toxoplasma gondii Seroprevalence: Vaccination against PCV2 and Muscle Enzyme Activity in Seropositive and Seronegative Pigs. Microorganisms. 2021; 9(5):1097. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051097
Chicago/Turabian StyleAthanasiou, Labrini V., Vasileios G. Papatsiros, Victoria M. Spanou, Eleni G. Katsogiannou, and Anna Dedousi. 2021. "Neospora caninum and/or Toxoplasma gondii Seroprevalence: Vaccination against PCV2 and Muscle Enzyme Activity in Seropositive and Seronegative Pigs" Microorganisms 9, no. 5: 1097. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051097