
Unicellular versus Filamentous: The Glacial Alga Ancylonema alaskana comb. et stat. nov. and Its Ecophysiological Relatedness to Ancylonema nordenskioeldii (Zygnematophyceae, Streptophyta)


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Harvest and Field Measurements
2.2. Cell Cleaning, Storage and Microscopy
2.3. Cultivation Assays
2.4. Molecular Characterisation
2.5. Phylogenetic Analysis
2.6. Photosynthesis
2.7. Analysis of Phenols
2.8. Isolation and Fractionation of Lipids
2.9. Shotgun Lipidomics
2.10. Fatty Acid Methyl Esters Analysis (FAMEs)
3. Results
3.1. Field Blooms
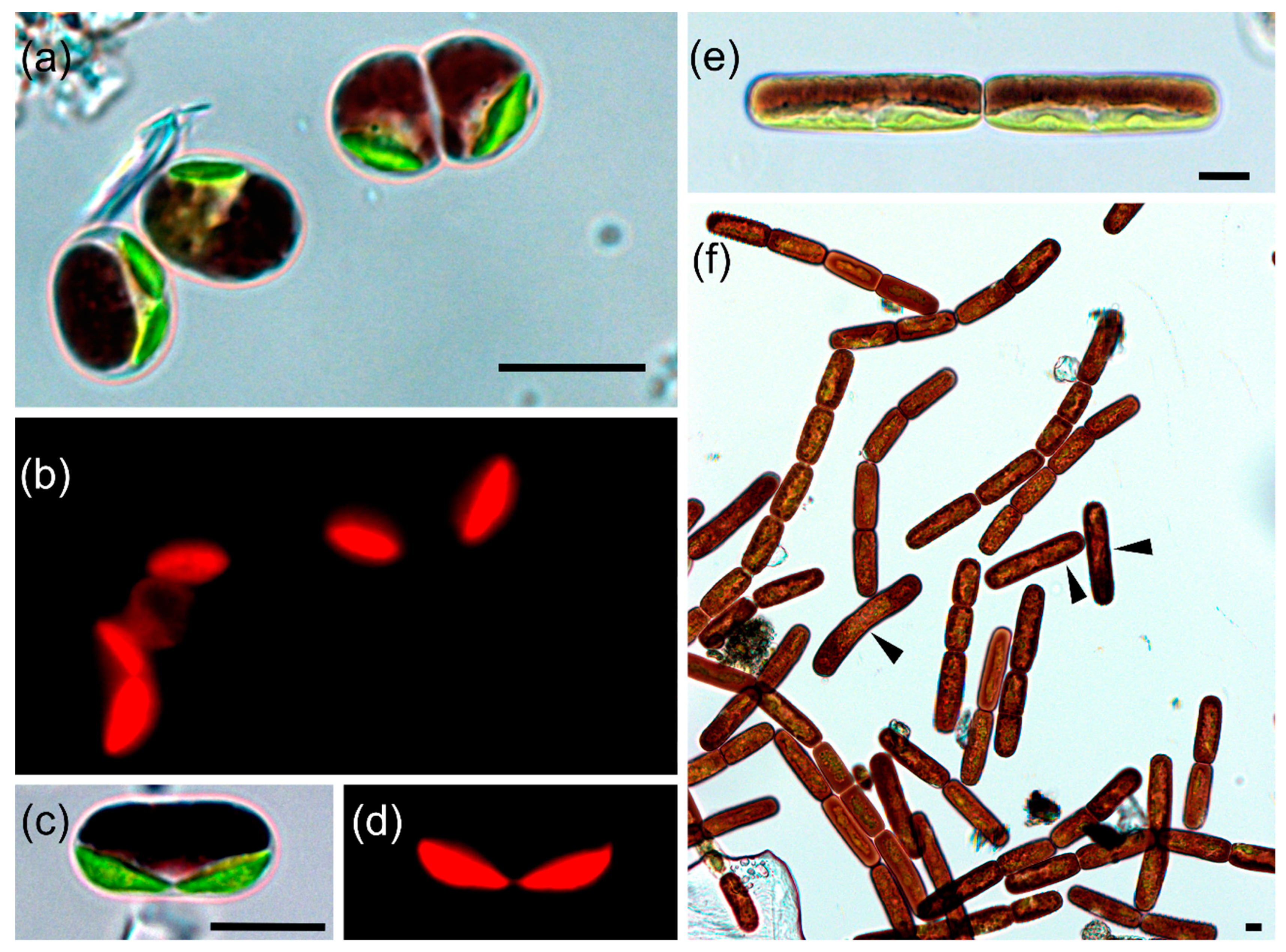
3.2. Taxonomic Treatment
3.3. Habitat Conditions, Population Densities and Cell Sizes of Glacial Algae
3.4. Cultivation Attempts
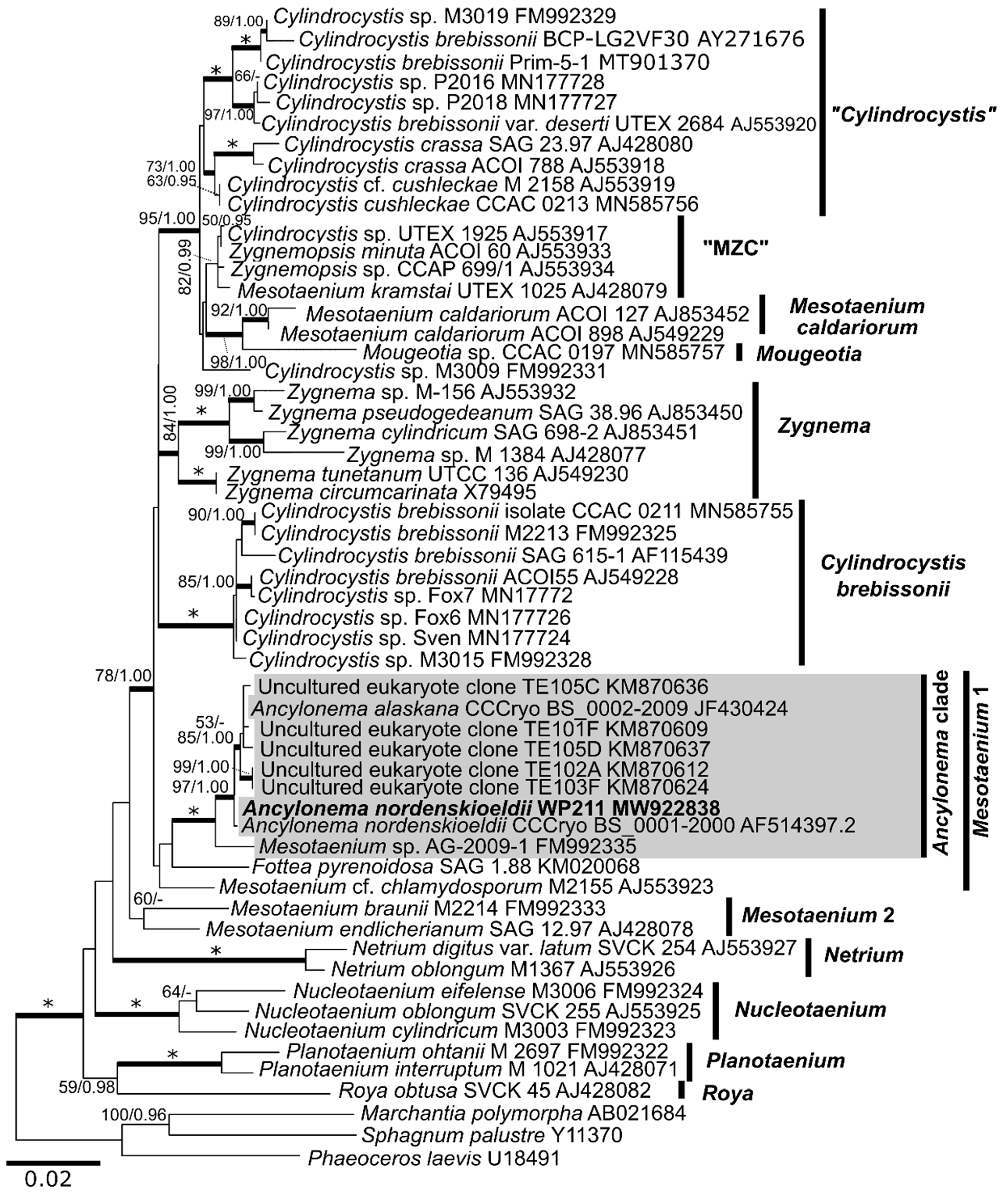
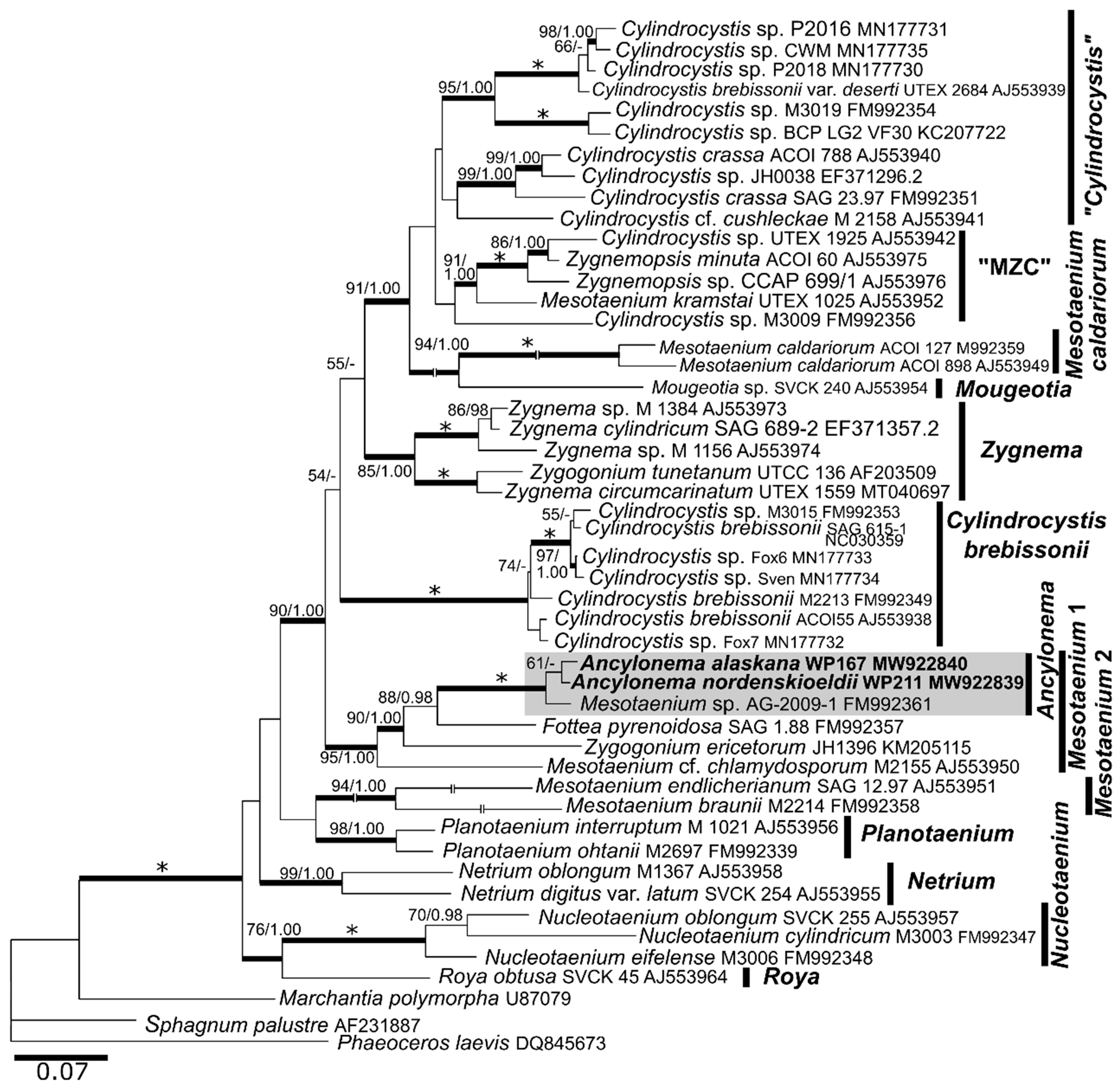
3.5. Molecular Taxonomy
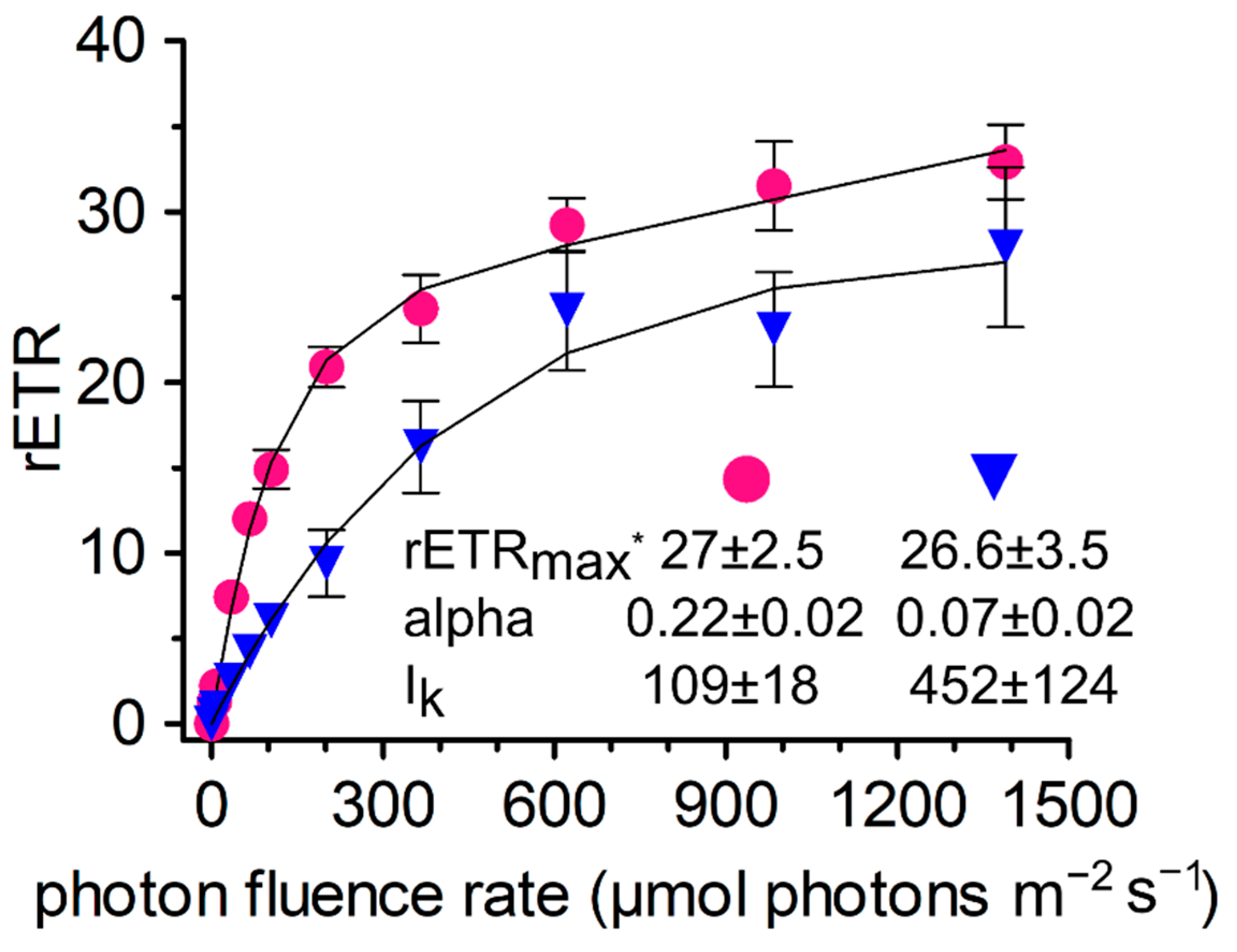
3.6. Photosynthesis
3.7. Phenolic Pigments
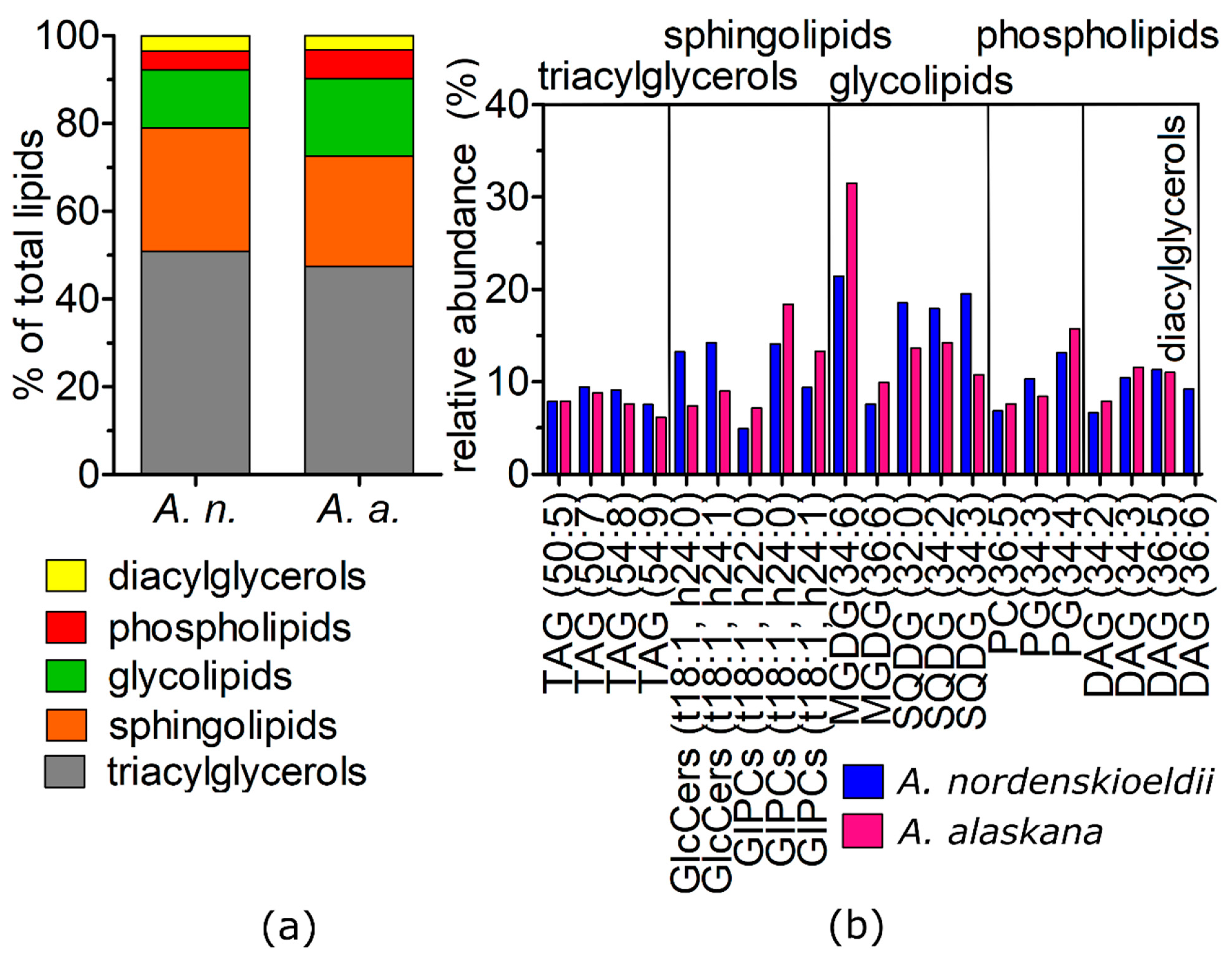
3.8. Shotgun Lipidomic
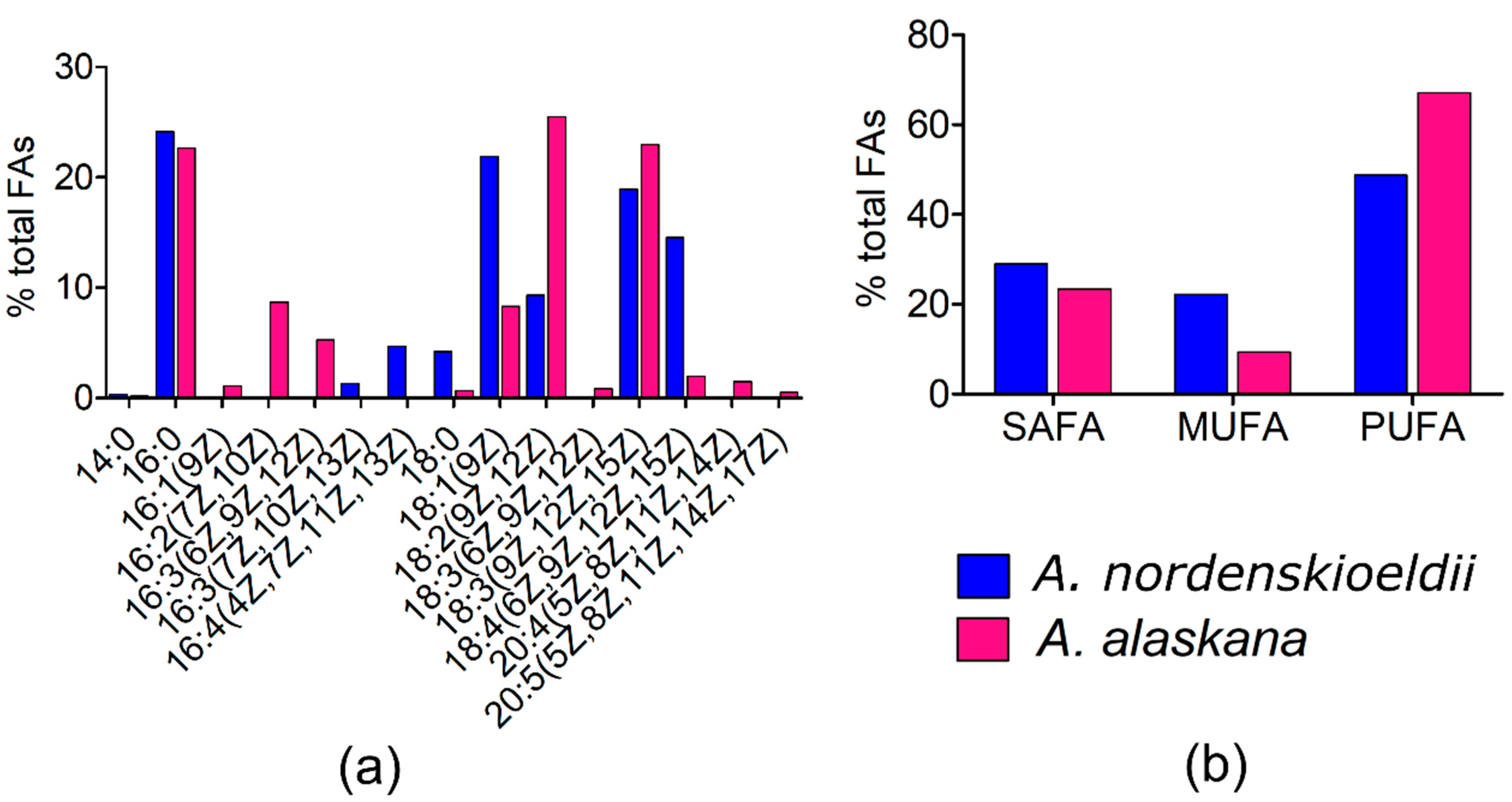
3.9. Fatty Acid (FA) Composition
4. Discussion
4.1. Geographical Distribution of Glacial Algae
4.2. Habitat Conditions, Population Densities and Cell Sizes of Glacial Algae
4.3. Phylogeny and Morphological Traits
4.4. Photosynthesis and Phenols
4.5. Lipidomics and Fatty Acids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anesio, A.M.; Lutz, S.; Chrismas, N.A.M.; Benning, L.G. The microbiome of glaciers and ice sheets. NPJ Biofilms Microbiomes. 2017, 3, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boetius, A.; Anesio, A.M.; Deming, J.W.; Mikucki, J.A.; Rapp, J.Z. Microbial ecology of the cryosphere: Sea ice and glacial habitats. Nat. Rev. Microbiol. 2015, 13, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Hotaling, S.; Hood, E.; Hamilton, T.L. Microbial ecology of mountain glacier ecosystems: Biodiversity, ecological connections and implications of a warming climate. Environ. Microbiol. 2017, 19, 2935–2948. [Google Scholar] [CrossRef] [Green Version]
- Hoham, R.W.; Remias, D. Snow and Glacial Algae: A Review. J. Phycol. 2020, 56, 264–282. [Google Scholar] [CrossRef] [Green Version]
- Ling, H.U.; Seppelt, R.D. Snow algae of the Windmill Islands, continental Antarctica. Mesotaenium berggrenii (Zygnematales, Chlorophyta) the alga of grey snow. Antarct. Sci. 1990, 2, 143–148. [Google Scholar] [CrossRef]
- Remias, D.; Holzinger, A.; Lütz, C. Physiology, ultrastructure and habitat of the ice alga Mesotaenium berggrenii (Zygnemaphyceae, Chlorophyta) from glaciers in the European Alps. Phycologia 2009, 48, 302–312. [Google Scholar] [CrossRef]
- Remias, D.; Holzinger, A.; Aigner, S.; Lütz, C. Ecophysiology and ultrastructure of Ancylonema nordenskiöldii (Zygnematales, Streptophyta), causing brown ice on glaciers in Svalbard (high arctic). Polar Biol. 2012, 35, 899–908. [Google Scholar] [CrossRef]
- Kol, E. Kryobiologie. Biologie und Limnologie des Schnees und Eises. I. Kryovegetation. In Die Binnengewässer; Elster, H.J., Ohle, W., Eds.; Band XXIV; Schweizerbart’sche Verlagsbuchhandlung: Stuttgart, Germany, 1968. [Google Scholar]
- Williamson, C.J.; Cameron, K.A.; Cook, J.M.; Žárský, J.D.; Stibal, M.; Edwards, A. Glacier algae: A dark past and a darker future. Front. Microbiol. 2019, 10, 524. [Google Scholar] [CrossRef] [Green Version]
- Barcytė, D.; Pilátová, J.; Mojzeš, P.; Nedbalová, L. The Arctic Cylindrocystis (Zygnematophyceae, Streptophyta) green algae are genetically and morphologically diverse and exhibit effective accumulation of polyphosphate. J. Phycol. 2020, 56, 217–232. [Google Scholar] [CrossRef]
- Remias, D.; Schwaiger, S.; Aigner, S.; Leya, T.; Stuppner, H.; Lütz, C. Characterization of an UV- and VIS-absorbing, purpurogallin-derived secondary pigment new to algae and highly abundant in Mesotaenium berggrenii (Zygnematophyceae, Chlorophyta), an extremophyte living on glaciers. FEMS Microbiol. Ecol. 2012, 79, 638–648. [Google Scholar] [CrossRef] [PubMed]
- Williamson, C.J.; Cook, J.; Tedstone, A.; Yallop, M.; McCutcheon, J.; Poniecka, E.; Campbell, D.; Irvine-Fynn, T.; McQuaid, J.; Tranter, M.; et al. Algal photophysiology drives darkening and melt of the Greenland Ice Sheet. Proc. Natl. Acad. Sci. USA 2020, 117, 5694–5705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yallop, M.L.; Anesio, A.M.; Perkins, R.G.; Cook, J.; Telling, J.; Fagan, D.; MacFarlane, J.; Stibal, M.; Barker, G.; Bellas, C.; et al. Photophysiology and albedo-changing potential of the ice algal community on the surface of the Greenland ice sheet. ISME J. 2012, 6, 2302–2313. [Google Scholar] [CrossRef] [Green Version]
- Lutz, S.; Anesio, A.M.; Jorge Villar, S.E.; Benning, L.G. Variations of algal communities cause darkening of a Greenland glacier. FEMS Microbiol. Ecol. 2014, 89, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Stibal, M.; Box, J.E.; Cameron, K.A.; Langen, P.L.; Yallop, M.L.; Mottram, R.H.; Khan, A.L.; Molotch, N.P.; Chrismas, N.A.M.; Quaglia, F.C.; et al. Algae drive enhanced darkening of bare ice on the Greenland ice sheet. Geophys. Res. Lett. 2017. [Google Scholar] [CrossRef]
- McCutcheon, J.; Lutz, S.; Williamson, C.; Cook, J.M.; Tedstone, A.J.; Vanderstraeten, A.; Wilson, S.A.; Stockdale, A.; Bonneville, S.; Anesio, A.M.; et al. Mineral phosphorus drives glacier algal blooms on the Greenland Ice Sheet. Nat. Commun. 2021, 12, 570. [Google Scholar] [CrossRef]
- Williamson, C.; Anesio, A.M.; Cook, J.; Tedstone, A.; Poniecka, E.; Holland, A.; Fagan, D.; Tranter, M.; Yallop, M. Ice algal bloom development on the surface of the Greenland Ice Sheet. FEMS Microbiol. Ecol. 2018, 94, fiy025. [Google Scholar] [CrossRef] [PubMed]
- Di Mauro, B.; Garzonio, R.; Baccolo, G.; Franzetti, A.; Pittino, F.; Leoni, B.; Remias, D.; Colombo, R.; Rossini, M. Glacier algae foster ice-albedo feedback in the European Alps. Sci. Rep. 2020, 10, 4739. [Google Scholar] [CrossRef] [Green Version]
- Procházková, L.; Leya, T.; Křížková, H.; Nedbalová, L. Sanguina nivaloides and Sanguina aurantia gen. et spp. nov. (Chlorophyta): The taxonomy, phylogeny, biogeography and ecology of two newly recognised algae causing red and orange snow. FEMS Microbiol. Ecol. 2019, 95, fiz064. [Google Scholar] [CrossRef] [Green Version]
- Procházková, L.; Remias, D.; Řezanka, T.; Nedbalová, L. Chloromonas nivalis subsp. tatrae, subsp. nov. (Chlamydomonadales, Chlorophyta): Re–examination of a snow alga from the High Tatra Mountains (Slovakia). Fottea 2018, 18, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Gontcharov, A.A.; Melkonian, M. Molecular phylogeny and revision of the genus Netrium (Zygnematophyceae, Streptophyta): Nucleotaenium gen. nov. J. Phycol. 2010, 46, 346–362. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Nedbalová, L.; Mihál, M.; Kvíderová, J.; Procházková, L.; Řezanka, T.; Elster, J. Identity, ecology and ecophysiology of planktic green algae dominating in ice-covered lakes on James Ross Island (northeastern Antarctic Peninsula). Extremophiles. 2017, 21, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kates, M.; Volcani, B.E. Biosynthetic pathways for phosphatidylsulfocholine, the sulfonium analogue of phosphatidylcholine. In Biological and Environmental Chemistry of DMSP and Related Sulfonium Compounds, in Diatoms; Kiene, R.P., Visscher, P.T., Keller, M.D., Kirst, G.O., Eds.; Springer: Boston, MA, USA, 1996; pp. 109–119. [Google Scholar]
- Saunders, R.D.; Horrocks, L.A. Simultaneous extraction and preparation for high-performance liquid chromatography of prostaglandins and phospholipids. Anal. Biochem. 1984, 143, 71–75. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Řezanka, T.; Rozentsvet, O.A. Lipid composition of three macrophytes from the Caspian Sea. Phytochemistry 1993, 33, 1015–1019. [Google Scholar] [CrossRef]
- Kol, E. The snow and ice algae of Alaska. Smithson. Misc. Collect. 1942, 101, 1–36. [Google Scholar]
- Takeuchi, N. The altitudinal distribution of snow algae on an Alaska glacier (Gulkana Glacier in the Alaska Range). Hydrol. Process. 2001, 15, 3447–3459. [Google Scholar] [CrossRef]
- Darcy, S.K.; Schmidt, J.L. Phylogeny of ulotrichalean algae from extreme high-altitude and high-latitude ecosystems. Polar Biol. 2015, 38, 689–697. [Google Scholar]
- Takeuchi, N.; Tanaka, S.; Irvine-Fynn, T.D.L.; Rassner, S.M.E.; Edwards, A. Variations in phototroph communities on the ablating bare-ice surface of glaciers on Brøggerhalvøya, Svalbard. Front. Earth Sci. 2019, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Lutz, S.; McCutcheon, J.; McQuaid, J.B.; Benning, L.G. The diversity of ice algal communities on the Greenland Ice Sheet as revealed by oligotyping. Microbial. Genomics. 2018, 4, e000159. [Google Scholar] [CrossRef]
- Takeuchi, N.; Uetake, J.; Fujita, K.; Aizen, V.B.; Nikitin, S.D. A snow algal community on Akkem Glacier in the Russian Altai Mountains. Ann. Glaciol. 2006, 43, 378–384. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, Y.; Kohshima, S.; Ohtani, S. A community of snow algae on a Himalayan glacier: Change of algal biomass and community structure with altitude. Arctic Alpine Res. 1997, 29, 126–137. [Google Scholar] [CrossRef]
- Berggren, S. Alger från Grönlands inlandis. In Öfversigt af Kongl. Vetenskaps-Akademiens Förhandlingar; P. A. Norstedt & Söner: Stockholm, Sweden, 1871; pp. 293–297. [Google Scholar]
- Wittrock, B. Om snöns och isens flora, särskildt i de arktiska trakterna. In Studier och Forskningar Föranledda af Mina Resor i Hoga Norden; Nordenskiöld, A.E., Ed.; F. & G. Beijers Forlag: Stockholm, Sweden, 1883; pp. 63–124. [Google Scholar]
- Lagerheim, G. Die Schnee Flora des Pichincha. Ein Beitrag zur Kenntnis der Nivalen Algen und Pilzen. Ber. der Dtsch. Bot. Gessellschaft. 1892, 10, 517–534. [Google Scholar]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. (Eds.) International Code of Nomenclature For algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017; Regnum Veg. Glashütten 159; Koeltz Botanical Books: Glashütten, Germany, 2018. [Google Scholar] [CrossRef]
- Webb, W.L.; Newton, M.; Starr, D. Carbon dioxide exchange of Alnus rubra. A mathematical model. Oecologia. 1974, 17, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, N.; Kohshima, S. A snow algal community on Tyndall Glacier in the Southern Patagonia Icefield, Chile. Arctic Alpine Res. 2004, 36, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Komárek, O.; Komárek, J. Contribution to the taxonomy and ecology of green cryosestic algae in the summer season 1995-96 at King George Island, S. Shetland Islands. Nova Hedwig. Beiheft. 2001, 123, 121–140. [Google Scholar]
- Kol, E. Über roten and grünen Schnee der Alpen. Verh Intern. Ver. Limnol. 1961, XIV, 912–917. [Google Scholar] [CrossRef]
- Krieger, W. Die Desmidiaceen Europas mit Berücksichtigung der aussereuropäischen Arten. Band 13. Abteilung 1, Teil 1, Lieferung 1 of Dr. L. Rabenhorst’s Kryptogamen-Flora von Deutschland, Österreich und der Schweiz; Akademische Verlagsgeselschaft M.B.H.: Leipzig, Germany, 1933. [Google Scholar]
- Yoshimura, Y.; Kohshima, S.; Takeuchi, N.; Seko, K.; Fujita, K. Himalayan ice-core dating with snow algae. J. Glaciol. 2000, 46, 335–340. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.; Xian, W.; Fu, Y.; Marin, B.; Keller, J.; Wu, T.; Sun, W.; Li, X.; Xu, Y.; Zhang, Y.; et al. Genomes of subaerial Zygnematophyceae provide insights into land plant evolution. Cell 2019, 179, 1057–1067.e14. [Google Scholar] [CrossRef]
- Stancheva, R.; Hall, J.D.; Herburger, K.; Lewis, L.A.; Mccourt, R.M.; Sheath, R.G. Phylogenetic position of Zygogonium ericetorum (Zygnematophyceae, Charophyta) from a high alpine habitat and ultrastructural characterization of unusual aplanospores. J. Phycol. 2014, 803, 790–803. [Google Scholar] [CrossRef] [Green Version]
- Broady, P.A. Six new species of terrestrial algae from Signy Island, South Orkney Islands, Antarctica. Brit. Phycol. J. 1976, 11, 387–405. [Google Scholar] [CrossRef]
- Herburger, K.; Remias, D.; Holzinger, A. The green alga Zygogonium ericetorum (Zygnematophyceae, Charophyta) shows high iron and aluminium tolerance: Protection mechanisms and photosynthetic performance. FEMS Microbiol. Ecol. 2016, 92, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nägeli, C. Gattungen einzelliger Algen, physiologisch und systematisch bearbeitet. Neue Denkschr. der Allg Schweiz. Gesellschaft für die Gesammten Naturwissenschaften. 1849, 10, 1–139. [Google Scholar]
- O’Neal, S.W.; Hoover, A.M. Comparison of UVB effects on growth and induction of UVB screening compounds in isolates of metaphytic algae from temperate zone streams and ponds. J. Phycol. 2018, 54, 818–828. [Google Scholar] [CrossRef]
- Nedbalová, L.; Sklenář, P. New records of snow algae from the Andes of Ecuador. Arnaldoa. 2008, 15, 17–20. [Google Scholar]
- Garduño-Solórzano, G.; Martínez-García, M.; Scotta Hentschke, G.; Lopes, G.; Castelo Branco, R.; Vasconcelos, V.M.O.; Campos, J.E.; López-Cano, R.; Quintanar-Zúñiga, R.E. The phylogenetic placement of Temnogametum (Zygnemataceae) and description of Temnogametum iztacalense sp. nov., from a tropical high mountain lake in Mexico. Eur. J. Phycol. 2020, 56, 1–15. [Google Scholar] [CrossRef]
- Kalisch, B.; Dörmann, P.; Hölzl, G. DGDG and Glycolipids in Plants and Algae. In Lipids in Plant and Algae Development. Subcellular Biochemistry; Nakamura, Y., Li-Beisson, Y., Eds.; Springer: Cham, Switzerland, 2016; pp. 51–83. [Google Scholar]
- Mock, T.; Kroon, B.M.A. Photosynthetic energy conversion under extreme conditions-II: The significance of lipids under light limited growth in Antarctic sea ice diatoms. Phytochemistry 2002, 61, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Lukeš, M.; Procházková, L.; Shmidt, V.; Nedbalová, L.; Kaftan, D. Temperature dependence of photosynthesis and thylakoid lipid composition in the red snow alga Chlamydomonas cf. nivalis (Chlorophyceae). FEMS Microbiol. Ecol. 2014, 89, 303–315. [Google Scholar] [CrossRef]
- Wang, X.; Li, W.; Li, M.; Welti, R. Profiling lipid changes in plant response to low temperatures. Physiol. Plantarum. 2006, 126, 90–96. [Google Scholar] [CrossRef]
- Holzinger, A.; Roleda, M.Y.; Lütz, C. The vegetative arctic freshwater green alga Zygnema is insensitive to experimental UV exposure. Micron 2009, 40, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Řezanka, T.; Nedbalová, L.; Procházková, L.; Sigler, K. Lipidomic profiling of snow algae by ESI-MS and silver-LC/APCI-MS. Phytochemistry 2014, 100, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Gronnier, J.; Germain, V.; Gouguet, P.; Cacas, J.L.; Mongrand, S. GIPC: Glycosyl inositol phospho ceramides, the major sphingolipids on earth. Plant Signal. Behav. 2016, 11, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markham, J.E.; Li, J.; Cahoon, E.B.; Jaworski, J.G. Separation and identification of major plant sphingolipid classes from leaves. J. Biol. Chem. 2006, 281, 22684–22694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, I.; Hodač, L.; Friedl, T.; Feussner, I. Fatty acid profiles and their distribution patterns in microalgae: A comprehensive analysis of more than 2000 strains from the SAG culture collection. BMC Plant Biol. 2011, 11, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichrtová, M.; Arc, E.; Stöggl, W.; Kranner, I.; Hájek, T.; Hackl, H.; Holzinger, A. Formation of lipid bodies and changes in fatty acid composition upon pre-akinete formation in Arctic and Antarctic Zygnema (Zygnematophyceae, Streptophyta) strains. FEMS Microbiol. Ecol. 2016, 92, fiw096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan-Kiss, R.M.; Priscu, J.C.; Pocock, T.; Gudynaite-Savitch, L.; Huner, N.P.A. Adaptation and acclimation of photosynthetic microorganisms to permanently cold environments. Microbiol. Mol. Biol. R. 2006, 70, 222–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Harvest | Glacier | Elevation | GPS |
|---|---|---|---|---|
| P39 | 6 September 2006 | Tiefenbach Ferner, AT | 3000 | 46°55′ N 10°56′ E |
| WP167 | 30 August 2017 | Gurgler Ferner, AT | 2728 | 46°48.280′ N 10°58.804′ E |
| WP210 | 22 August 2018 | Morteratsch Glacier, CH | 2211 | 46°24.999′ N 9°56.021′ E |
| WP211 | 22 August 2018 | Morteratsch Glacier, CH | 2275 | 46°24.847′ N 9°56.032′ E |
| WP213 | 24 August 2018 | Morteratsch Glacier, CH | 2161 | 46°25.195′ N 9°55.913′ E |
| WP249 | 18 August 2020 | Morteratsch Glacier, CH | 2203 | 46°25.075′ N 9°55.985′ E |
| WP251 | 25 August 2020 | Gurgler Ferner, AT | 2727 | 46°48.327′ N 10°58.739′ E |
| Species | Sample | Cell Width × Length (µm) | Cells mL−1 |
|---|---|---|---|
| A. alaskana | P39 1 | 8.7 ± 0.8 × 12.1 ± 2.8; 7.3–11.6 × 8.1–20.6 (83) | – |
| A. alaskana | WP167 | 7.6 ± 0.3 × 12.1 ± 1.9; 7–8.4 × 8.5–17.1 (30) | 4.77 × 105 ± 0.07 × 105 |
| A. alaskana | WP212 | 8.5 ± 0.7 × 12.8 ± 2.7; 7.5–10.2 × 9.4–19.7 (22) | – |
| A. alaskana | WP251 | 8.4 ± 0.8 × 11.4 ± 2.4; 7.0–10.5 × 8.4–16.3 (27) | – |
| A. nordenskioeldii | WP210 | 12.9 ± 1.1 × 29.4 ± 6.8; 10.7–15.2 × 19.6–50 (19) | – |
| A. nordenskioeldii | WP211 | 13.6 ± 0.8 × 37.5 ± 7.6; 12.4–14.7 × 22.1–51 (28) | 4.8 × 105 ± 0.25 × 105 |
| A. nordenskioeldii | WP249 | 13.7 ± 0.7 × 28.3 ± 5.5; 12.6–15 × 20.6–43.4 (33) | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Procházková, L.; Řezanka, T.; Nedbalová, L.; Remias, D. Unicellular versus Filamentous: The Glacial Alga Ancylonema alaskana comb. et stat. nov. and Its Ecophysiological Relatedness to Ancylonema nordenskioeldii (Zygnematophyceae, Streptophyta). Microorganisms 2021, 9, 1103. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051103
Procházková L, Řezanka T, Nedbalová L, Remias D. Unicellular versus Filamentous: The Glacial Alga Ancylonema alaskana comb. et stat. nov. and Its Ecophysiological Relatedness to Ancylonema nordenskioeldii (Zygnematophyceae, Streptophyta). Microorganisms. 2021; 9(5):1103. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051103
Chicago/Turabian StyleProcházková, Lenka, Tomáš Řezanka, Linda Nedbalová, and Daniel Remias. 2021. "Unicellular versus Filamentous: The Glacial Alga Ancylonema alaskana comb. et stat. nov. and Its Ecophysiological Relatedness to Ancylonema nordenskioeldii (Zygnematophyceae, Streptophyta)" Microorganisms 9, no. 5: 1103. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051103