
Isolation Procedure for CP E. coli from Caeca Samples under Review towards an Increased Sensitivity

 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
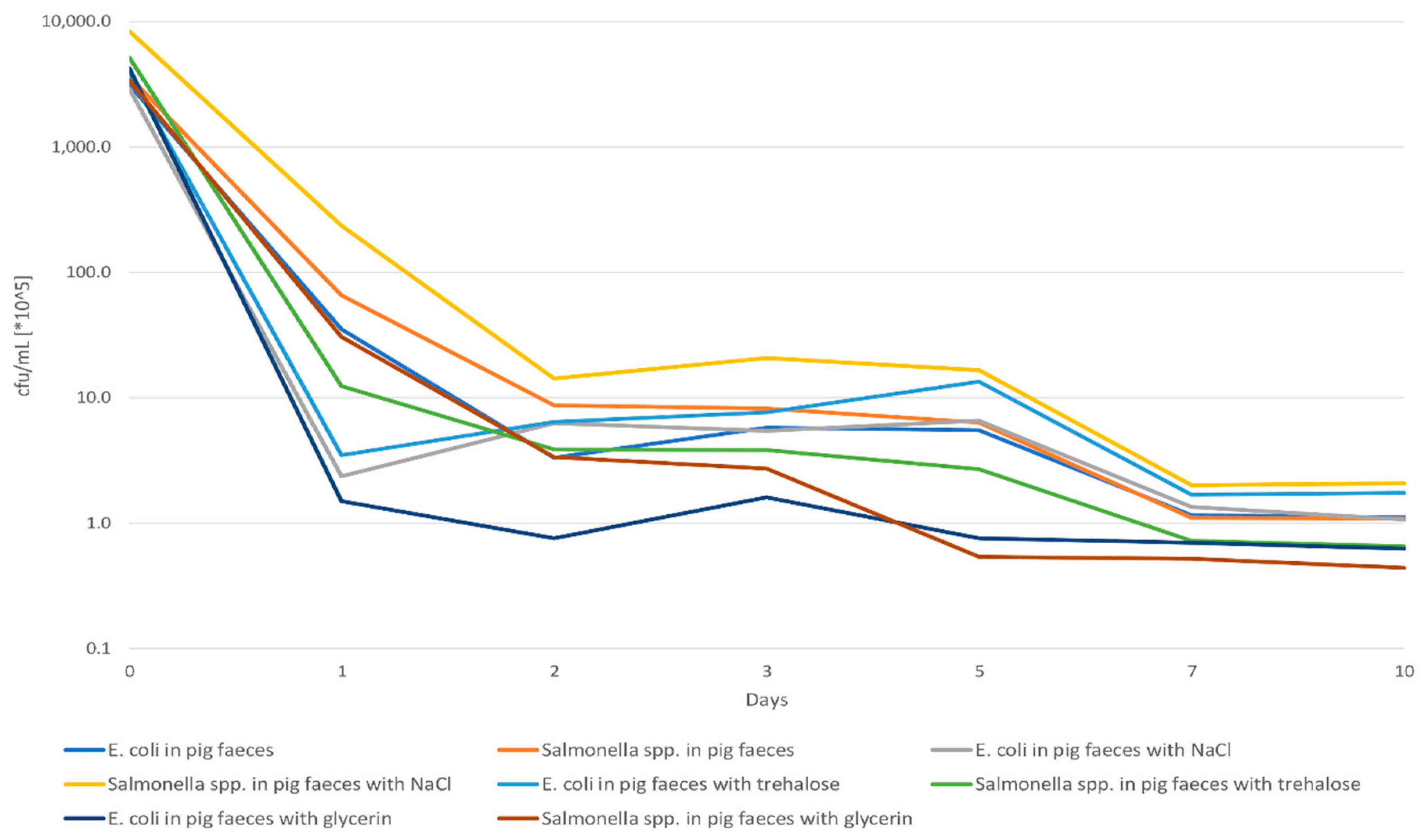
2.1. Pre-Studies
2.2. Method Comparison
2.2.1. Bacterial Strains
2.2.2. Sample Preparation for the Method Comparison
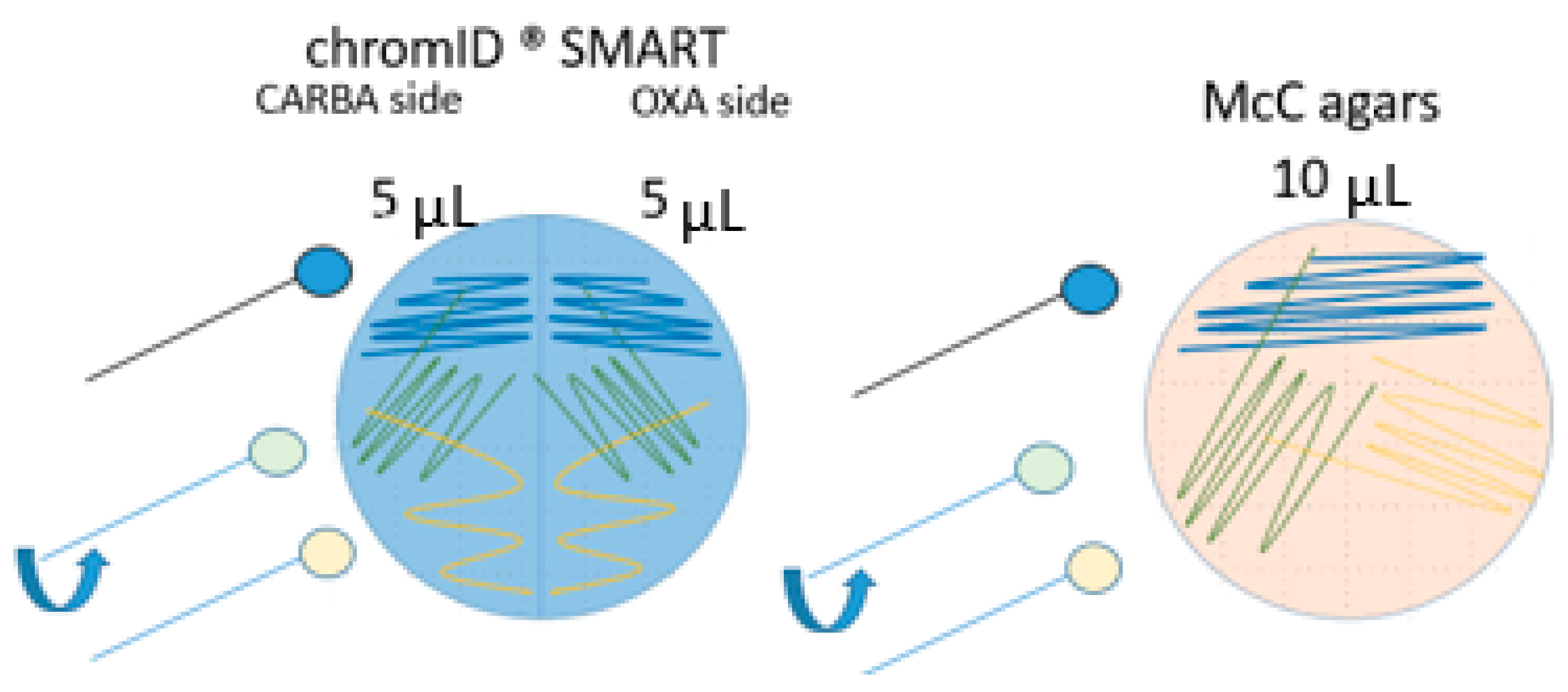
2.2.3. Isolation and Detection Methods
2.2.4. Typing
Antimicrobial Susceptibility-Testing (AST)
MALDI-ToF MS
Molecular Typing of CP E. coli
2.2.5. Statistics
3. Results
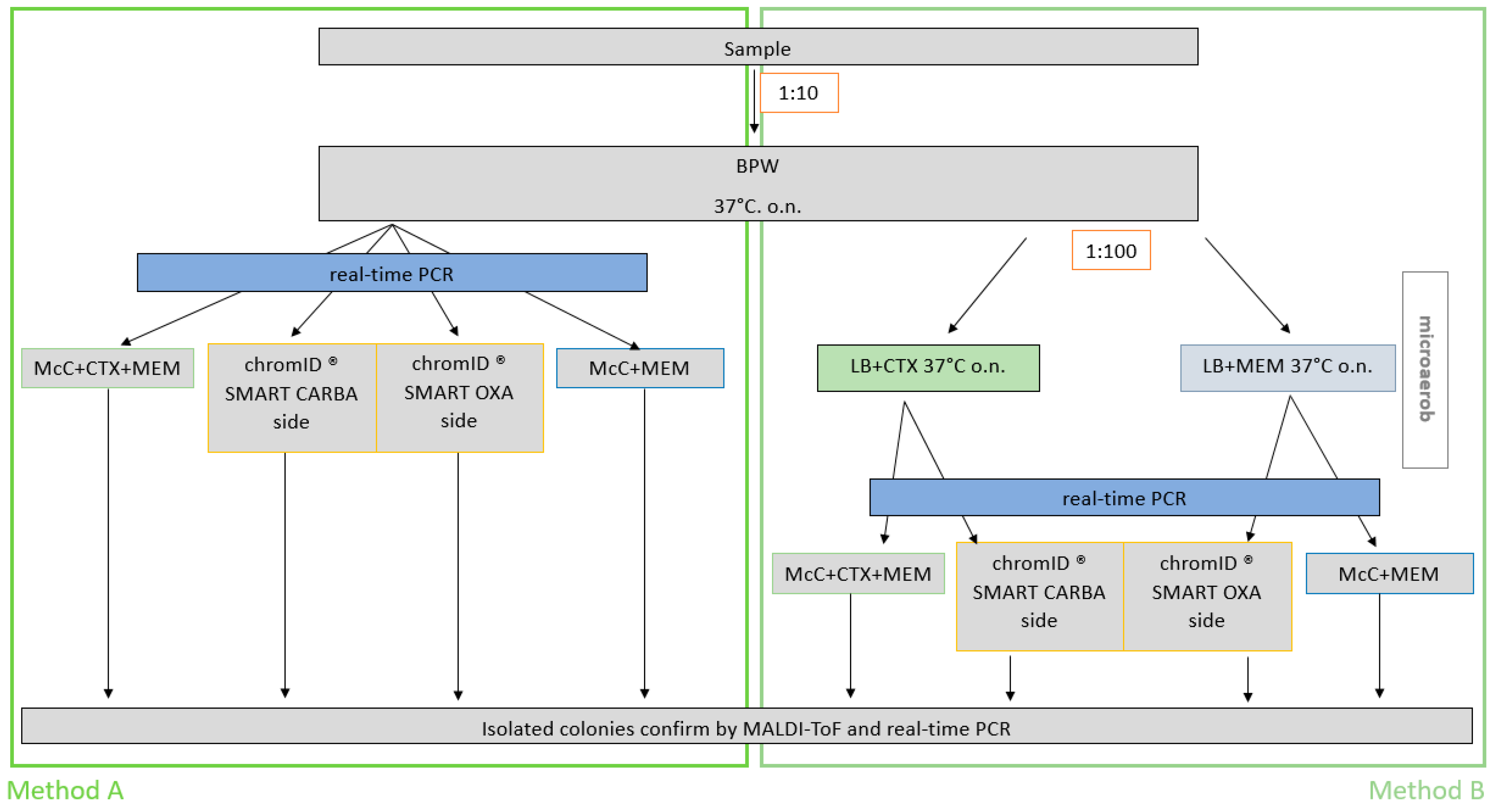
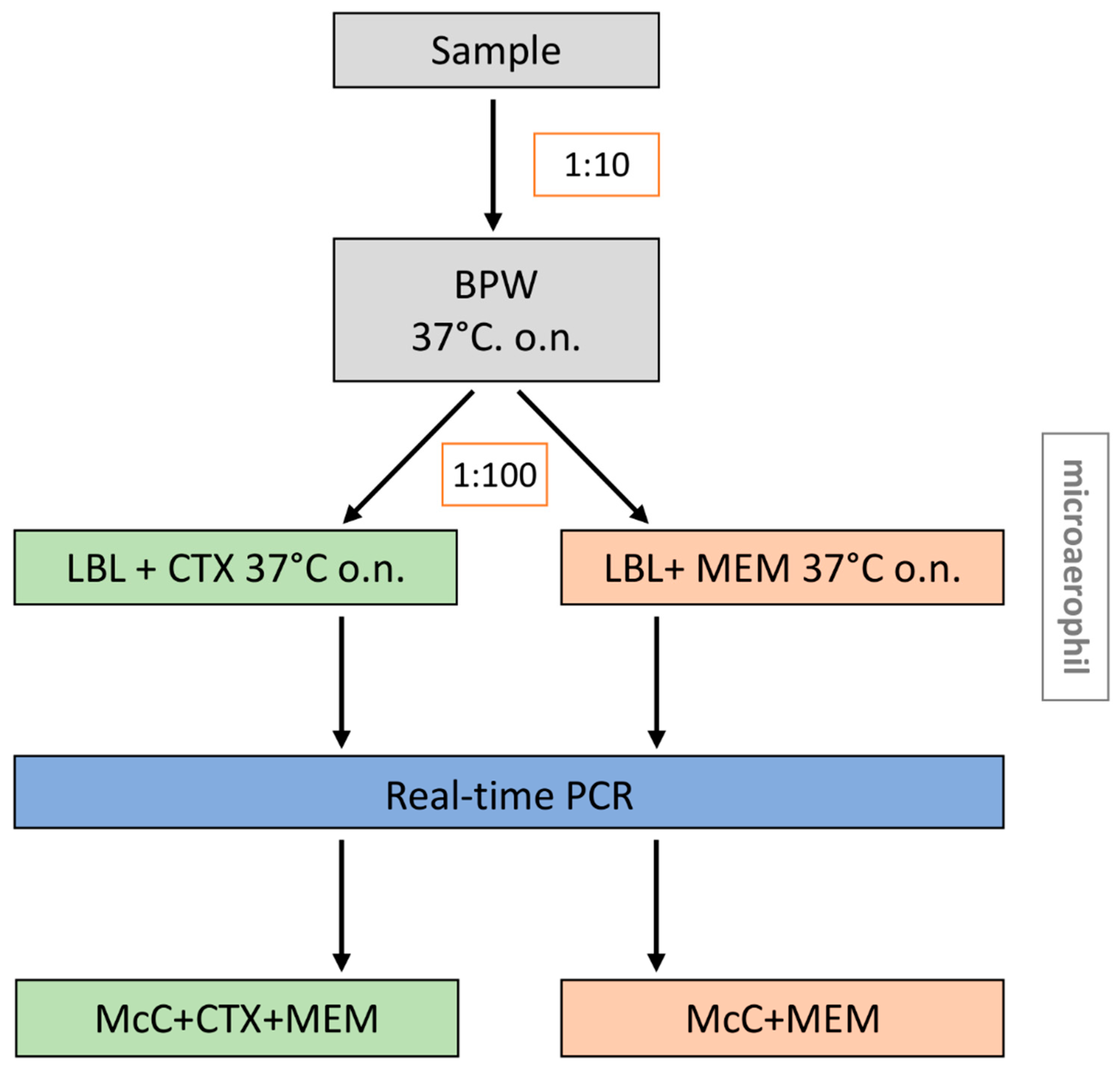
3.1. Modification of the EURL-Protocol for Isolation of CP E. coli from Caeca Samples
3.2. The Comparison of the Modified Method with the EURL-Method for Isolation of the CP E. coli from Caeca Samples
3.3. Molecular Detection of the Carbapenemase Genes
4. Discussion
4.1. Modification Steps of the Official Isolation Protocol
4.2. The Comparison of Both Microbiological Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Critically Important Antimicrobials for Human Medicine, 6th ed.; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Madec, J.Y.; Haenni, M.; Nordmann, P.; Poirel, L. Extended-spectrum β-lactamase/AmpC- and carbapenemase-producing Enterobacteriaceae in animals: A threat for humans? Clin. Microbiol. Infect. 2017, 23, 826–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aworh, M.K.; Kwaga, J.; Okolocha, E.; Mba, N.; Thakur, S. Prevalence and risk factors for multi-drug resistant Escherichia coli among poultry workers in the Federal Capital Territory, Abuja, Nigeria. PLoS ONE 2019, 14, e0225379. [Google Scholar] [CrossRef]
- EFSA/ECDC. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from hu-mans, animals and food in 2016. EFSA J. 2018, 16, e05182. [Google Scholar]
- Köck, R.; Daniels-Haardt, I.; Becker, K.; Mellmann, A.; Friedrich, A.; Mevius, D.; Schwarz, S.; Jurke, A. Carbapenem-resistant Enterobacteriaceae in wildlife, food-producing, and companion animals: A systematic review. Clin. Microbiol. Infect. 2018, 24, 1241–1250. [Google Scholar] [CrossRef] [Green Version]
- Taneja, N.; Sharma, M. Antimicrobial resistance in the environment: The Indian scenario. Indian J. Med. Res. 2019, 149, 119–128. [Google Scholar] [CrossRef]
- Miriagou, V.; Cornaglia, G.; Edelstein, M.; Galani, I.; Giske, C.G.; Gniadkowski, M.; Malamou-Lada, E.; Martinez-Martinez, L.; Navarro, F.; Nordmann, P.; et al. Acquired car-bapenemases in Gram-negative bacterial pathogens: Detection and surveillance issues. Clin. Microbiol. Infect. 2010, 16, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botelho, J.; Roberts, A.P.; León-Sampedro, R.; Grosso, F.; Peixe, L. Carbapenemases on the move: It’s good to be on ICEs. Mob. DNA 2018, 9, 37. [Google Scholar] [CrossRef] [Green Version]
- Palzkill, T. Metallo-β-lactamase structure and function. Ann. N. Y. Acad. Sci. 2013, 1277, 91–104. [Google Scholar] [CrossRef]
- Salahuddin, P.; Kumar, A.; Kha, A.U. Structure, function of serine and Metallo-β-lactamases and their Inhibitors. Curr. Protein. Pept. Sci. 2018, 19, 130–144. [Google Scholar] [CrossRef]
- Bush, K. Past and present perspectives on β-lactamases. Antimicrob. Agents Chemother. 2018, 62, e01076-18. [Google Scholar] [CrossRef] [Green Version]
- Woodford, N.; Wareham, D.W.; Guerra, B.; Teale, C. Carbapenemase-producing Enterobacteriaceae and non-Enterobacteriaceae from animals and the environment: An emerging public health risk of our own making? J. Antimicrob. Chemother. 2014, 69, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Grundmann, H.; Glasner, C.; Albiger, B.; Aanensen, D.M.; Tomlinson, C.T.; Andrasević, A.T.; Cantón, R.; Carmeli, Y.; Friedrich, A.W.; Giske, C.G.; et al. Occurrence of carbapenemase-producing Klebsiella pneumoniae and Escherichia coli in the European survey of carbapenemase-producing Enterobacteriaceae (EuSCAPE): A prospective, multinational study. Lancet Infect. Dis. 2017, 17, 153–163. [Google Scholar] [CrossRef] [Green Version]
- EFSA and ECDC. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017. EFSA J. 2019, 17, e05598. [Google Scholar]
- Fischer, J.; Rodriguez, I.; Schmoger, S.; Friese, A.; Roesler, U.; Helmuth, R.; Guerra, B. Escherichia coli producing VIM-1 car-bapenemase isolated on a pig farm. J. Antimicrob. Chemother. 2012, 67, 1793–1795. [Google Scholar] [CrossRef]
- Schrijver, R.; Stijntjes, M.; Rodríguez-Baño, J.; Tacconelli, E.; Rajendran, N.B.; Voss, A. Review of antimicrobial resistance surveillance programmes in livestock and meat in EU with focus on humans. Clin. Microbiol. Infect. 2018, 24, 577–590. [Google Scholar] [CrossRef] [Green Version]
- Simner, P.J.; Gilmour, M.W.; DeGagne, P.; Nichol, K.; Karlowsky, J.A. Evaluation of Five Chromogenic Agar Media and the Rosco Rapid Carb Screen Kit for Detection and Confirmation of Carbapenemase Production in Gram-Negative Bacilli. J. Clin. Microbiol. 2014, 53, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, K.M.; Winstanley, T.G.; Lanyon, C.; Cummings, S.P.; Raza, M.W.; Perry, J.D. Comparison of Four Chromogenic Culture Media for Carbapenemase-Producing Enterobacteriaceae. J. Clin. Microbiol. 2012, 50, 3102–3104. [Google Scholar] [CrossRef] [Green Version]
- Vrioni, G.; Daniil, I.; Voulgari, E.; Ranellou, K.; Koumaki, V.; Ghirardi, S.; Kimouli, M.; Zambardi, G.; Tsakris, A. Comparative evaluation of a prototype chromogenic medium (ChromID CARBA) for detecting carbapenemase-producing Enterobacte-riaceae in surveillance rectal swabs. J. Clin. Microbiol. 2012, 50, 1841–1846. [Google Scholar] [CrossRef] [Green Version]
- Papadimitriou-Olivgeris, M.; Bartzavali, C.; Christofidou, M.; Bereksi, N.; Hey, J.; Zambardi, G.; Spiliopoulou, I. Performance of chromID® CARBA medium for carbapenemases-producing enterobacteriaceae detection during rectal screening. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 35–40. [Google Scholar] [CrossRef]
- Pauly, N.; Hammerl, J.A.; Grobbel, M.; Tenhagen, B.-A.; Käsbohrer, A.; Bisenius, S.; Fuchs, J.; Horlacher, S.; Lingstädt, H.; Mauermann, U.; et al. ChromID® CARBA Agar Fails to Detect Carbapenem-Resistant Enterobacteriaceae with Slightly Reduced Susceptibility to Carbapenems. Front. Microbiol. 2020, 11, 1678. [Google Scholar] [CrossRef]
- Irrgang, A.; Tenhagen, B.-A.; Pauly, N.; Schmoger, S.; Kaesbohrer, A.; Hammerl, J.A. Characterization of VIM-1-Producing E. coli Isolated from a German Fattening Pig Farm by an Improved Isolation Procedure. Front. Microbiol. 2019, 10, 2256. [Google Scholar] [CrossRef]
- Pauly, N.; Hammerl, J.A.; Schwarz, S.; Grobbel, M.; Meemken, D.; Malorny, B.; Tenhagen, B.-A.; Käsbohrer, A.; Irrgang, A. Co-occurrence of the blaVIM-1 and blaSHV-12 genes on an IncHI2 plasmid of an Escherichia coli isolate recovered from German livestock. J. Antimicrob. Chemother. 2021, 76, 531–533. [Google Scholar] [CrossRef] [PubMed]
- Algara, M.M.; Kuczyńska-Wiśnik, D.; Dębski, J.; Stojowska-Swędrzyńska, K.; Sominka, H.; Bukrejewska, M.; Laskowska, E. Trehalose protects Escherichia coli against carbon stress manifested by protein acetylation and aggregation. Mol. Microbiol. 2019, 112, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Kuczyńska-Wiśnik, D.; Stojowska, K.; Matuszewska, E.; Leszczyńska, D.; Algara, M.M.; Augustynowicz, M.; Laskowska, E. Lack of intracellular trehalose affects formation of Escherichia coli persister cells. Microbiol. Spectr. 2015, 161 Pt 4, 786–796. [Google Scholar] [CrossRef] [Green Version]
- Achá, S.; Kühn, I.; Mbazima, G.; Colque-Navarro, P.; Möllby, R. Changes of viability and composition of the Escherichia coli flora in faecal samples during long time storage. J. Microbiol. Methods 2005, 63, 229–238. [Google Scholar] [CrossRef]
- Hollander, D.H.; Nell, E.E. Improved Preservation of Treponema pallidum and Other Bacteria by Freezing with Glycerol. Appl. Microbiol. 1954, 2, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Jordan, D. Sodium chloride enhances recovery and growth of acid-stressed E. coli O157:H7. Lett. Appl. Microbiol. 2001, 32, 312–315. [Google Scholar] [CrossRef]
- Mañas, P.; Pagan, R.; Leguérinel, I.; Condon, S.; Mafart, P.; Sala, F. Effect of sodium chloride concentration on the heat resistance and recovery of Salmonella typhimurium. Int. J. Food Microbiol. 2001, 63, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Mossel, D.A.; Harrewijn, G.A.; Zadelhoff, C.F.N.-V. Standardization of the selective inhibitory effect of surface active compounds used in media for the detection of Enterobacteriaceae in foods and water. Health Lab. Sci. 1974, 11, 260–267. [Google Scholar]
- Andersson, D.I.; Hughes, D. Microbiological effects of sublethal levels of antibiotics. Nat. Rev. Genet. 2014, 12, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Girlich, D.; Poirel, L. Detection of Carbapenemase Producers in Enterobacteriaceae by Use of a Novel Screening Medium. J. Clin. Microbiol. 2012, 50, 2761–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gullberg, E.; Albrecht, L.M.; Karlsson, C.; Sandegren, L.; Andersson, D.I. Selection of a Multidrug Resistance Plasmid by Sublethal Levels of Antibiotics and Heavy Metals. mBio 2014, 5, e01918-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swayne, R.; Ludlam, H.; Shet, V.; Woodford, N.; Curran, M. Real-time TaqMan PCR for rapid detection of genes encoding five types of non-metallo- (class A and D) carbapenemases in Enterobacteriaceae. Int. J. Antimicrob. Agents 2011, 38, 35–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Zee, A.; Roorda, L.; Bosman, G.; Fluit, A.C.; Hermans, M.; Smits, P.H.; Van Der Zanden, A.G.; Witt, R.T.; Van Coppenraet, L.E.B.; Stuart, J.C.; et al. Multi-centre evaluation of real-time multiplex PCR for detection of carbapenemase genes OXA-48, VIM, IMP, NDM and KPC. BMC Infect. Dis. 2014, 14, 27. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.R-project.org/ (accessed on 11 March 2021).
- Octavia, S.; Lan, R. The Family Enterobacteriaceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014; pp. 225–286. ISBN 9783642389221. [Google Scholar]
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like carbapenemases: The phantom menace. J. Antimicrob. Chemother. 2012, 67, 1597–1606. [Google Scholar] [CrossRef] [Green Version]
- Codjoe, F.S.; Donkor, E.S. Carbapenem Resistance: A Review. Med. Sci. 2017, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Viau, R.; Frank, K.M.; Jacobs, M.R.; Wilson, B.; Kaye, K.; Donskey, C.J.; Perez, F.; Endimiani, A.; Bonomo, R.A. Intestinal Carriage of Carbapenemase-Producing Organisms: Current Status of Surveillance Methods. Clin. Microbiol. Rev. 2015, 29, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Brand, P.; Gobeli, S.; Perreten, V. Pathotyping and antibiotic resistance of porcine enterovirulent Escherichia coli strains from Switzerland (2014–2015). Schweiz Arch Tierheilkd 2017, 159, 373–380. [Google Scholar] [CrossRef]
- Mo, S.S.; Urdahl, A.M.; Madslien, K.; Sunde, M.; Nesse, L.L.; Slettemeås, J.S.; Norström, M. What does the fox say? Monitoring antimicrobial resistance in the environment using wild red foxes as an indicator. PLoS ONE 2018, 13, e0198019. [Google Scholar] [CrossRef]
- Hasman, H.; Agersø, Y.; Cavaco, L.M. Validation of selective MacConkey agar plates supplemented with 1 mg/L cefo-taxime for monitoring of ESBL- and AmpC-producing E. coli in meat and caeca samples. LABORATORY PROTOCOL Authored by the EURL-AR. 2017. Available online: https://www.eurl-ar.eu/CustomerData/Files/Folders/21-protocols/390_protocol-for-validation-of-macconkey-and-ctx-agar-plates-final03112017.pdf (accessed on November 2013).
- Xu, H.-S.; Roberts, N.; Singleton, F.L.; Attwell, R.W.; Grimes, D.J.; Colwell, R.R. Survival and viability of nonculturableEscherichia coli andVibrio cholerae in the estuarine and marine environment. Microb. Ecol. 1982, 8, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Mendis, N.; Trigui, H.; Oliver, J.D.; Faucher, S.P. The importance of the viable but non-culturable state in human bacterial pathogens. Front. Microbiol. 2014, 5, 258. [Google Scholar] [CrossRef] [Green Version]
- Grützke, J.; Malorny, B.; Hammerl, J.A.; Busch, A.; Tausch, S.H.; Tomaso, H.; Deneke, C. Fishing in the Soup—Pathogen Detection in Food Safety Using Metabarcoding and Metagenomic Sequencing. Front. Microbiol. 2019, 10, 1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method A + ChromID® SMART CARBA agars | Method A + in-house agars | Method B + in-house agars | Method B + ChromID® SMART CARBA agars | |
|---|---|---|---|---|
| Sensitivity | 75 (57.8–87.9) | 100 (90.2–100) | 86.1 (70.5–95.3) | 66.7 (49–81.4) |
| Specificity | 100 (81.5–100) | 100 (81.5–100) | 100 (81.5–100) | 100 (81.5–100) |
| False discovery rate | 0 (0–12.8) | 0 (0–9.7) | 0 (0–11.2) | 0 (0–14.2) |
| False omission rate | 33.3 (16.5–54) | 0 (0–18.5) | 21.7 (7.5–43.7) | 40 (22.7–59.4) |
| Accuracy | 83.3 (70.7–92.1) | 100 (93.4–100) | 90.7 (79.7–96.9) | 77.8 (64.4–88) |
| ChromID® SMART CARBA Side | ChromID® SMART OXA Side | McC + MEM | McC + CTX + MEM | |
|---|---|---|---|---|
| Method A | 48/108 (44.45%) | 25/108 (23.15%) | 40/108 (37.04%) | 36/108 (33.36%) |
| Method B | 43/108 (39.82%) | 17/108 (15.74%) | 32/108 (29.63%) | 39/108 (36.14%) |
| BPW | LB + CTX | LB + MEM | Both LB Enrichments | |
|---|---|---|---|---|
| Sensitivity | 61.1 (43.5–76.9) | 83.3 (67.2–93.6) | 58.3 (40.8–74.4) | 94.4 (81.3–99.3) |
| Specifity | 94.4 (72.7–99.6) | 94.4 (72.7–99.6) | 100 (81.5–100) | 94.4 (72.7–99.9) |
| False discovery rate | 4.3 (0.1–21.9) | 3.2 (0.1–16.7) | 0 (0–16.1) | 2.9 (0.1–14.9) |
| False omission rate | 45.2 (27.3–64) | 26.1 (10.2–48.4) | 45.4 (28.1–63.6) | 10.5 (1.3–33.1) |
| Accuracy | 72.2 (58.4–83.5) | 87.0 (75.1–94.7) | 72.2 (58.4–83.5) | 94.4 (84.6–98.8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pauly, N.; Klaar, Y.; Skladnikiewicz-Ziemer, T.; Juraschek, K.; Grobbel, M.; Hammerl, J.A.; Hemmers, L.; Käsbohrer, A.; Schwarz, S.; Meemken, D.; et al. Isolation Procedure for CP E. coli from Caeca Samples under Review towards an Increased Sensitivity. Microorganisms 2021, 9, 1105. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051105
Pauly N, Klaar Y, Skladnikiewicz-Ziemer T, Juraschek K, Grobbel M, Hammerl JA, Hemmers L, Käsbohrer A, Schwarz S, Meemken D, et al. Isolation Procedure for CP E. coli from Caeca Samples under Review towards an Increased Sensitivity. Microorganisms. 2021; 9(5):1105. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051105
Chicago/Turabian StylePauly, Natalie, Yvonne Klaar, Tanja Skladnikiewicz-Ziemer, Katharina Juraschek, Mirjam Grobbel, Jens André Hammerl, Lukas Hemmers, Annemarie Käsbohrer, Stefan Schwarz, Diana Meemken, and et al. 2021. "Isolation Procedure for CP E. coli from Caeca Samples under Review towards an Increased Sensitivity" Microorganisms 9, no. 5: 1105. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051105