
Functional Characterization of the mazEF Toxin-Antitoxin System in the Pathogenic Bacterium Agrobacterium tumefaciens

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, Primers, and Chemicals
2.2. Cloning of mazE, mazF, mazEF, and mazF Mutants
2.3. Cell Toxicity on Plates and in Liquid Medium
2.4. Cellular Target Assay of DNA, RNA, and Protein Synthesis In Vivo
2.5. Protein Expression and Purification
2.6. mRNA Stability Analysis Using Northern Blot
2.7. Endoribonuclease Activity Assay of MazE-at, MazF-at, and MazEF-at In Vitro
2.8. Secondary Structure Analysis by Circular Dichroism
2.9. Protein Interaction Analysis by GST Pull-Down Assay with Beads
3. Results
3.1. MazF-at Is Toxic in E. coli and Is Neutralized by MazE-at
3.2. The MazF-at Belongs to the MazF Family and E24 of MazF-at Is an Active Residue for Its Toxicity
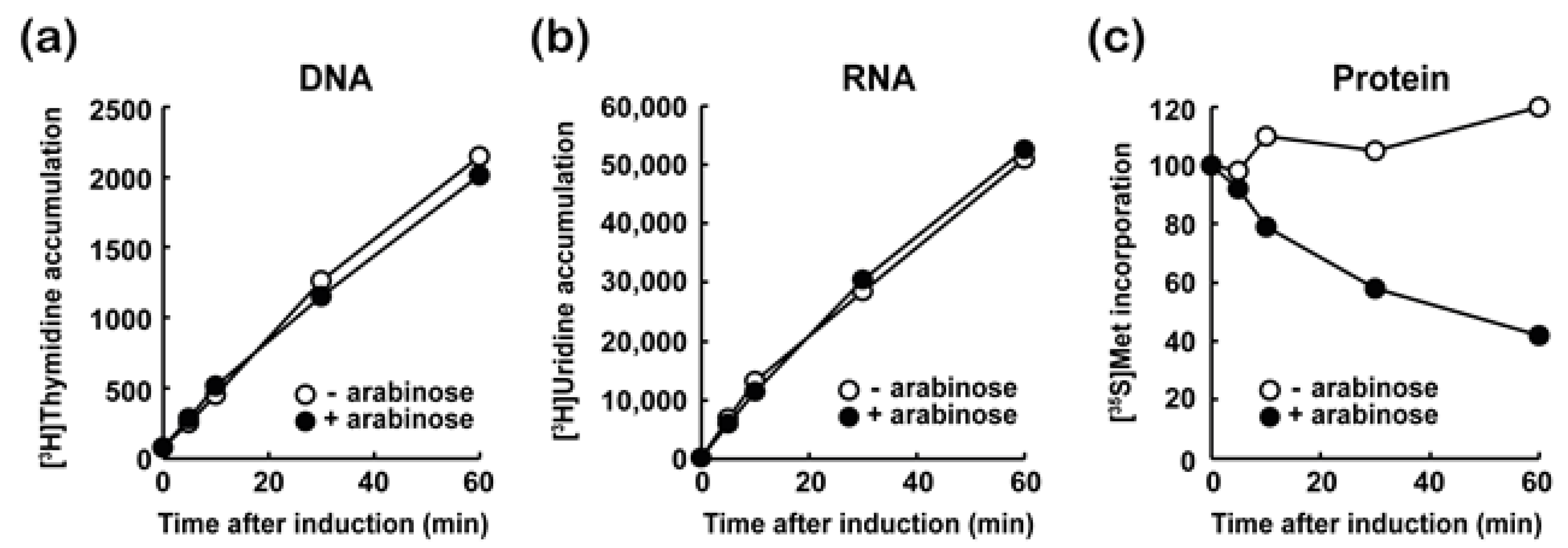
3.3. MazF-at Inhibits Protein Synthesis
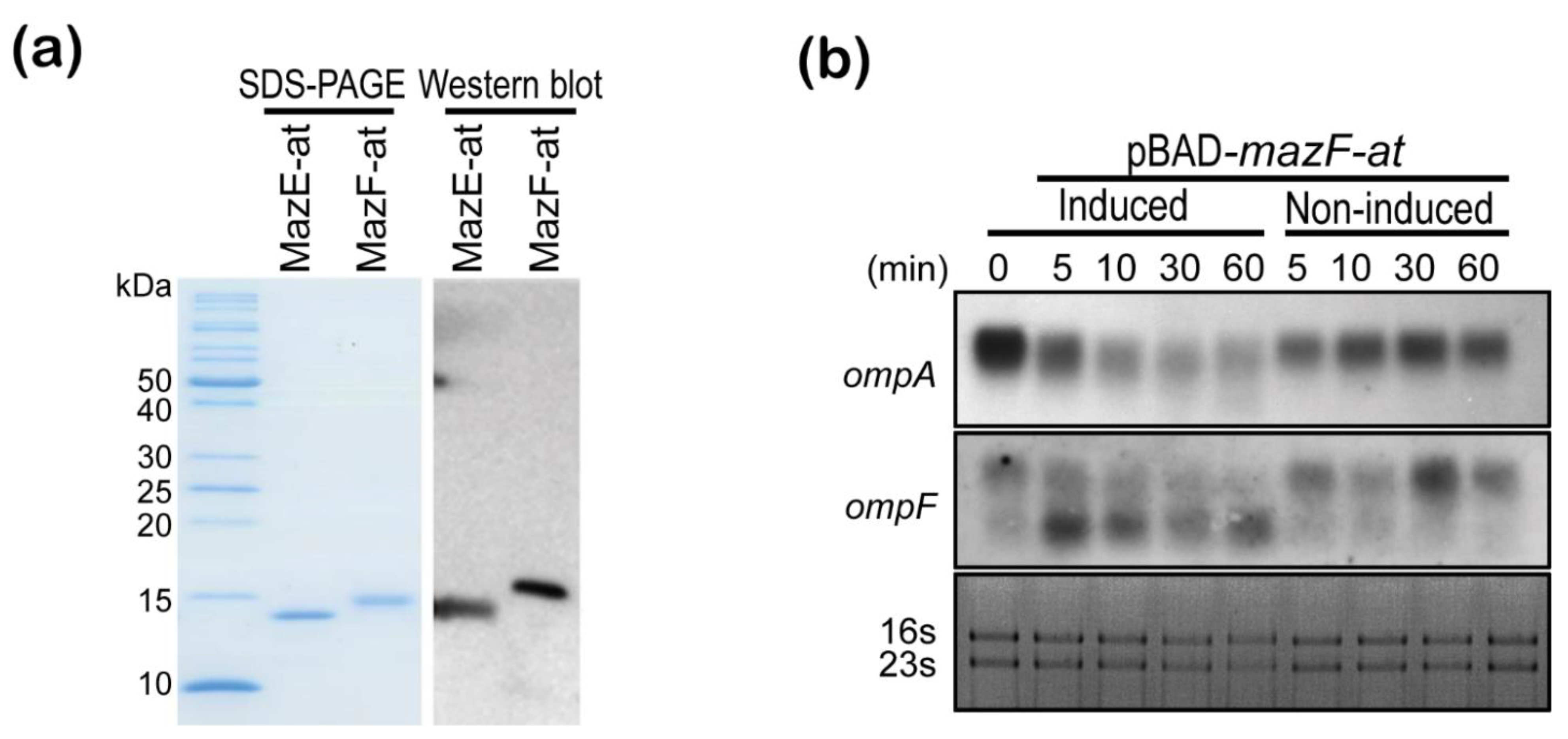
3.4. MazF-at Is an Endoribonuclease
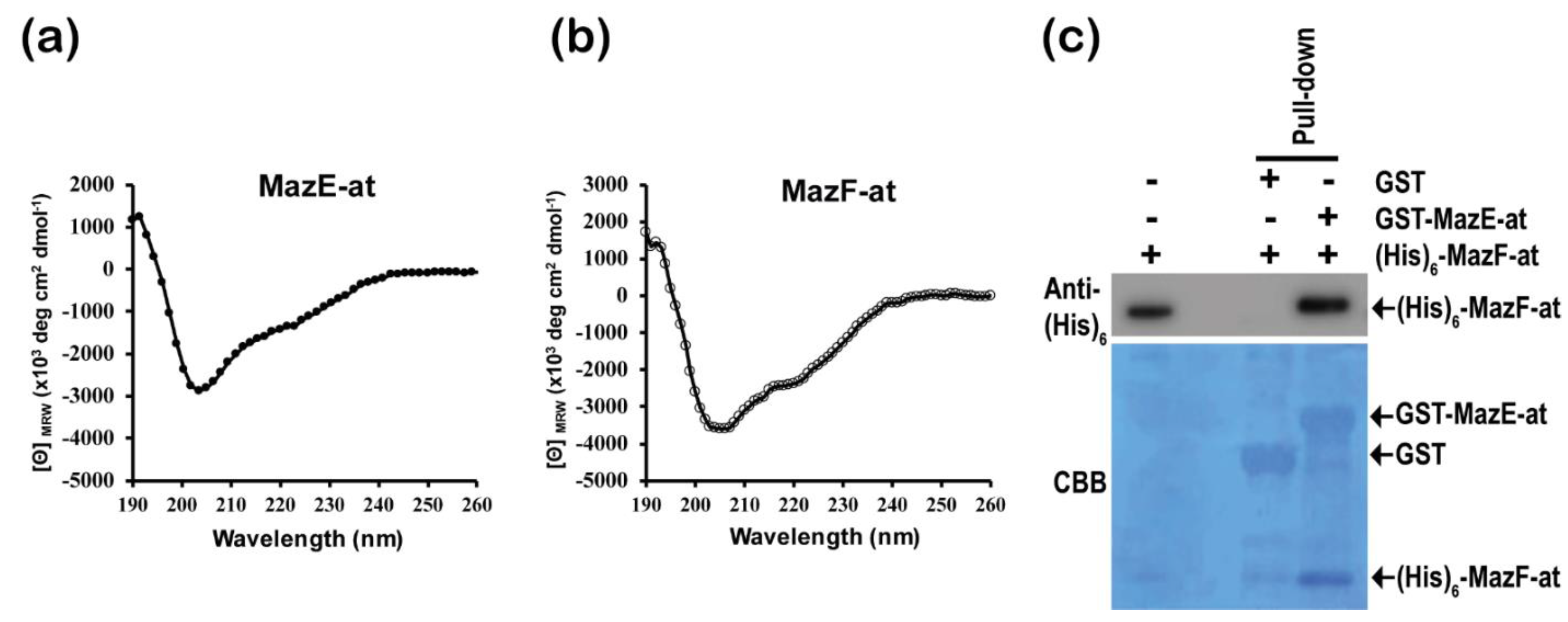
3.5. Characterization of Purified MazE-at and MazF-at Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fraikin, N.; Goormaghtigh, F.; Van Melderen, L. Type II Toxin-Antitoxin Systems: Evolution and Revolutions. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogura, T.; Hiraga, S. Mini-F plasmid genes that couple host cell division to plasmid proliferation. Proc. Natl. Acad. Sci. USA 1983, 80, 4784–4788. [Google Scholar] [CrossRef] [Green Version]
- Gerdes, K.; Rasmussen, P.B.; Molin, S. Unique type of plasmid maintenance function: Postsegregational killing of plasmid-free cells. Proc. Natl. Acad. Sci. USA 1986, 83, 3116–3120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo, A.; de Torrontegui, G.; Diaz, R. Identification of components of a new stability system of plasmid R1, ParD, that is close to the origin of replication of this plasmid. Mol. Gen. Genet. 1987, 210, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Dewan, P.C.; Barua, B.; Varadarajan, R. Additional role for the ccd operon of F-plasmid as a transmissible persistence factor. Proc. Natl. Acad. Sci. USA 2012, 109, 12497–12502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, Targets, and Triggers: An Overview of Toxin-Antitoxin Biology. Mol. Cell 2018, 70, 768–784. [Google Scholar] [CrossRef] [Green Version]
- Aizenman, E.; Engelberg-Kulka, H.; Glaser, G. An Escherichia coli chromosomal “addiction module” regulated by guanosine [corrected] 3′,5′-bispyrophosphate: A model for programmed bacterial cell death. Proc. Natl. Acad. Sci. USA 1996, 93, 6059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamruzzaman, M.; Iredell, J. A ParDE-family toxin antitoxin system in major resistance plasmids of Enterobacteriaceae confers antibiotic and heat tolerance. Sci. Rep. 2019, 9, 9872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.M.; Koo, J.S.; Kim, C.M.; Kim, D.H.; Lee, B.J. mRNA Interferase Bacillus cereus BC0266 Shows MazF-Like Characteristics Through Structural and Functional Study. Toxins 2020, 12, 380. [Google Scholar] [CrossRef] [PubMed]
- Cook, G.M.; Robson, J.R.; Frampton, R.A.; McKenzie, J.; Przybilski, R.; Fineran, P.C.; Arcus, V.L. Ribonucleases in bacterial toxin-antitoxin systems. Biochim. Biophys. Acta 2013, 1829, 523–531. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Hara, H.; Kato, I.; Inouye, M. Insights into the mRNA cleavage mechanism by MazF, an mRNA interferase. J. Biol. Chem. 2005, 280, 3143–3150. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, K.; Zavialov, A.V.; Pavlov, M.Y.; Elf, J.; Gerdes, K.; Ehrenberg, M. The bacterial toxin RelE displays codon-specific cleavage of mRNAs in the ribosomal A site. Cell 2003, 112, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Jurenaite, M.; Markuckas, A.; Suziedeliene, E. Identification and characterization of type II toxin-antitoxin systems in the opportunistic pathogen Acinetobacter baumannii. J. Bacteriol. 2013, 195, 3165–3172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korch, S.B.; Contreras, H.; Clark-Curtiss, J.E. Three Mycobacterium tuberculosis Rel toxin-antitoxin modules inhibit mycobacterial growth and are expressed in infected human macrophages. J. Bacteriol. 2009, 191, 1618–1630. [Google Scholar] [CrossRef] [Green Version]
- Denkovskienė, E.; Paškevičius, Š.; Stankevičiūtė, J.; Gleba, Y.; Ražanskienė, A. Control of T-DNA Transfer from Agrobacterium tumefaciens to Plants Based on an Inducible Bacterial Toxin-Antitoxin System. Mol. Plant-Microbe Interact. 2020, 33, 1142–1149. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Kiyokawa, K.; Tanaka, K.; Moriguchi, K.; Suzuki, K. Novel toxin-antitoxin system composed of serine protease and AAA-ATPase homologues determines the high level of stability and incompatibility of the tumor-inducing plasmid pTiC58. J. Bacteriol. 2009, 191, 4656–4666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ames, J.R.; McGillick, J.; Murphy, T.; Reddem, E.; Bourne, C.R. Identifying a Molecular Mechanism That Imparts Species-Specific Toxicity to YoeB Toxins. Front. Microbiol. 2020, 11, 959. [Google Scholar] [CrossRef] [PubMed]
- Pavelich, I.J.; Maehigashi, T.; Hoffer, E.D.; Ruangprasert, A.; Miles, S.J.; Dunham, C.M. Monomeric YoeB toxin retains RNase activity but adopts an obligate dimeric form for thermal stability. Nucleic Acids Res. 2019, 47, 10400–10413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGillick, J.; Ames, J.R.; Murphy, T.; Bourne, C.R. A YoeB toxin cleaves both RNA and DNA. Sci. Rep. 2021, 11, 3592. [Google Scholar] [CrossRef]
- Baker, K.E.; Mackie, G.A. Ectopic RNase E sites promote bypass of 5’-end-dependent mRNA decay in Escherichia coli. Mol. Microbiol. 2003, 47, 75–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohm, G.; Muhr, R.; Jaenicke, R. Quantitative analysis of protein far UV circular dichroism spectra by neural networks. Protein Eng. 1992, 5, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Harrison, E.M.; Bi, D.; Tai, C.; He, X.; Ou, H.Y.; Rajakumar, K.; Deng, Z. TADB: A web-based resource for Type 2 toxin-antitoxin loci in bacteria and archaea. Nucleic Acids Res. 2011, 39, D606–D611. [Google Scholar] [CrossRef] [Green Version]
- Li, G.Y.; Zhang, Y.; Chan, M.C.; Mal, T.K.; Hoeflich, K.P.; Inouye, M.; Ikura, M. Characterization of dual substrate binding sites in the homodimeric structure of Escherichia coli mRNA interferase MazF. J. Mol. Biol. 2006, 357, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Zorzini, V.; Mernik, A.; Lah, J.; Sterckx, Y.G.; De Jonge, N.; Garcia-Pino, A.; De Greve, H.; Versees, W.; Loris, R. Substrate Recognition and Activity Regulation of the Escherichia coli mRNA Endonuclease MazF. J. Biol. Chem. 2016, 291, 10950–10960. [Google Scholar] [CrossRef] [Green Version]
- Sterckx, Y.G.; De Gieter, S.; Zorzini, V.; Hadzi, S.; Haesaerts, S.; Loris, R.; Garcia-Pino, A. An efficient method for the purification of proteins from four distinct toxin-antitoxin modules. Protein Expr. Purif. 2015, 108, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.; Chen, Z.; Hou, J.; Wang, Y.; Guo, M.; Cao, J.; Wang, L.; Xu, H.; Tian, B.; Zhao, Y. MazEF Toxin-Antitoxin System-Mediated DNA Damage Stress Response in Deinococcus radiodurans. Front. Genet. 2021, 12, 632423. [Google Scholar] [CrossRef]
- Bezrukov, F.; Prados, J.; Renzoni, A.; Panasenko, O.O. MazF toxin causes alterations in Staphylococcus aureus transcriptome, translatome and proteome that underlie bacterial dormancy. Nucleic Acids Res. 2021, 49, 2085–2101. [Google Scholar] [CrossRef]
- Drozdetskiy, A.; Cole, C.; Procter, J.; Barton, G.J. JPred4: A protein secondary structure prediction server. Nucleic Acids Res. 2015, 43, W389–W394. [Google Scholar] [CrossRef]
- Chowdhury, N.; Kwan, B.W.; McGibbon, L.C.; Babitzke, P.; Wood, T.K. Toxin MqsR cleaves single-stranded mRNA with various 5’ ends. Microbiologyopen 2016, 5, 370–377. [Google Scholar] [CrossRef]
- Culviner, P.H.; Laub, M.T. Global Analysis of the E. coli Toxin MazF Reveals Widespread Cleavage of mRNA and the Inhibition of rRNA Maturation and Ribosome Biogenesis. Mol. Cell 2018, 70, 868–880.e10. [Google Scholar] [CrossRef] [Green Version]
- Rothenbacher, F.P.; Suzuki, M.; Hurley, J.M.; Montville, T.J.; Kirn, T.J.; Ouyang, M.; Woychik, N.A. Clostridium difficile MazF toxin exhibits selective, not global, mRNA cleavage. J. Bacteriol. 2012, 194, 3464–3474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mets, T.; Lippus, M.; Schryer, D.; Liiv, A.; Kasari, V.; Paier, A.; Maivali, U.; Remme, J.; Tenson, T.; Kaldalu, N. Toxins MazF and MqsR cleave Escherichia coli rRNA precursors at multiple sites. RNA Biol. 2017, 14, 124–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.Y.; Lee, B.J. Structure, Biology, and Therapeutic Application of Toxin-Antitoxin Systems in Pathogenic Bacteria. Toxins 2016, 8, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain and Plasmid | Description | Source |
|---|---|---|
| Strains | ||
| A. tumefaciens EHA105 | Genomic DNA for cloning | Lab stock |
| E. coli DH5α | General cloning host | Lab stock |
| E. coli BL21(DE3) | General protein expression host | Lab stock |
| E. coli BW25113 | Wild type | Lab stock |
| Plasmids | ||
| pBAD24 | Gene expression vector, PBAD, amp | Lab stock |
| pBAD24-mazE-at | mazE in pBAD24 | This study |
| pBAD24- mazF-at | mazF in pBAD24 | This study |
| pBAD24- mazEF-at | mazEF in pBAD24 | This study |
| pBAD24- mazF-at (E24N) | Mutant MazF of E24N in pBAD24 | This study |
| pBAD24- mazF-at (E24Q) | Mutant MazF of E24Q in pBAD24 | This study |
| pBAD24- mazF-at (E24K) | Mutant MazF of E24K in pBAD24 | This study |
| pBAD24- mazF-at (E24D) | Mutant MazF of E24D in pBAD24 | This study |
| pBAD24- mazF-at (E24A) | Mutant MazF of E24A in pBAD24 | This study |
| pET28a | Protein expression vector, PIPTG, kan | Lab stock |
| pET28- mazE-at | mazE in pET28a | This study |
| pET28- mazF-at | mazF in pET28a | This study |
| pET28- mazEF-at | mazEF in pET28a | This study |
| pGEX6p-1 | Protein expression vector, PIPTG, amp | Lab stock |
| pGEX6p-1-mazE-at | mazE in pGEX6p-1 | This study |
| Primer | Sequence | Source |
|---|---|---|
| A-mazE-F | 5′-TATAGGATCCATGACCGTGACCACGA-3′ | This study |
| A-mazE-R | 5′-TATAAAGCTTTCACAACGCTTCTTTG-3′ | This study |
| T-mazF-F | 5′-TATACATATGGTCCGCAACCAGAT-3′ | This study |
| T-mazF-R | 5′-TATACTCGAGTCAAGCTGGATCGATCAT-3′ | This study |
| TA-mazEF-F | 5′-TATAGGATCCATGACCGTGACCACGA-3′ | This study |
| TA-mazEF-R | 5′-TATACTCGAGTCAAGCTGGATCGATCAT-3′ | This study |
| Pull-mazE-F | 5′-TATAGGATCCATGACCGTGACCACGA-3′ | This study |
| Pull-mazE-R | 5′-TATAGCGGCCGCTCACAACGCTTCTTTGC-3′ | This study |
| Mu-MazF-E24N-F | 5′-GTCGTAGGCAGCAACATCAAGGACGAA-3′ | This study |
| Mu-MazF-E24N-R | 5′-TTCGTCCTTGATGTTGCTGCCTACGAC-3′ | This study |
| Mu-MazF-E24Q-F | 5′-GTCGTAGGCAGCCAAATCAAGGACGAA-3′ | This study |
| Mu-MazF-E24Q-R | 5′-TTCGTCCTTGATTTGGCTGCCTACGAC-3′ | This study |
| Mu-MazF-E24K-F | 5′-GTCGTAGGCAGCAAAATCAAGGACGAA-3′ | This study |
| Mu-MazF-E24K-R | 5′-TTCGTCCTTGATTTCGCTGCCTACGAC-3′ | This study |
| Mu-MazF-E24D-F | 5′-GTCGTAGGCAGCGACATCAAGGACGAA-3′ | This study |
| Mu-MazF-E24D-R | 5′-TTCGTCCTTGATGTCGCTGCCTACGAC-3′ | This study |
| Mu-MazF-E24A-F | 5′-GTCGTAGGCAGCGCAATCAAGGACGAA-3′ | This study |
| Mu-MazF-E24A-R | 5′-TTCGTCCTTGATTGCGCTGCCTACGAC-3′ | This study |
| ompA1-F | 5′-GGTGCATACAAAGCTCAGGG-3′ | [6] |
| ompA1-R | 5′-GTGACTGCGTACTCAACACC-3′ | [6] |
| ompF1-F | 5′-CTTTGGTCTGGTTGATGGCC-3′ | [6] |
| ompF1-R | 5′-ACTTCAGACCAGTAGCCCAC-3′ | [6] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, W.; Yamaguchi, Y.; Park, J.-Y.; Park, S.-H.; Lee, H.-W.; Lim, B.-K.; Otto, M.; Inouye, M.; Yoon, M.-H.; Park, J.-H. Functional Characterization of the mazEF Toxin-Antitoxin System in the Pathogenic Bacterium Agrobacterium tumefaciens. Microorganisms 2021, 9, 1107. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051107
Choi W, Yamaguchi Y, Park J-Y, Park S-H, Lee H-W, Lim B-K, Otto M, Inouye M, Yoon M-H, Park J-H. Functional Characterization of the mazEF Toxin-Antitoxin System in the Pathogenic Bacterium Agrobacterium tumefaciens. Microorganisms. 2021; 9(5):1107. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051107
Chicago/Turabian StyleChoi, Wonho, Yoshihiro Yamaguchi, Ji-Young Park, Sang-Hyun Park, Hyeok-Won Lee, Byung-Kwan Lim, Michael Otto, Masayori Inouye, Min-Ho Yoon, and Jung-Ho Park. 2021. "Functional Characterization of the mazEF Toxin-Antitoxin System in the Pathogenic Bacterium Agrobacterium tumefaciens" Microorganisms 9, no. 5: 1107. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051107