
Biofilm-Forming Potential of Ocular Fluid Staphylococcus aureus and Staphylococcus epidermidis on Ex Vivo Human Corneas from Attachment to Dispersal Phase


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Cultures and Characterisation
2.2. Determination of Biofilm Formation by Various Methods
2.2.1. Congo Red Agar Method
2.2.2. Crystal Violetmethod
2.2.3. [2,3-Bis (2-methoxy-4-nitro-5-sulfophenyl)-5-(phenylamino) carbonyl-2H-tetra-zolium hydroxide)] Method (XTT)
2.3. Monitoring the Thickness of the Biofilm and Visualisation of Extracellular Polymeric Substance (EPS) by Confocal Laser Scanning Microscopy
2.4. Visualisation of Biofilm on Cover Slips by Scanning Electron Microscopy
2.5. Visualisation of Biofilm on Human Cadaveric Cornea by Scanning Electron Microscopy
2.6. Antibiotic Susceptibility in Planktonic and Biofilm Phase
3. Results
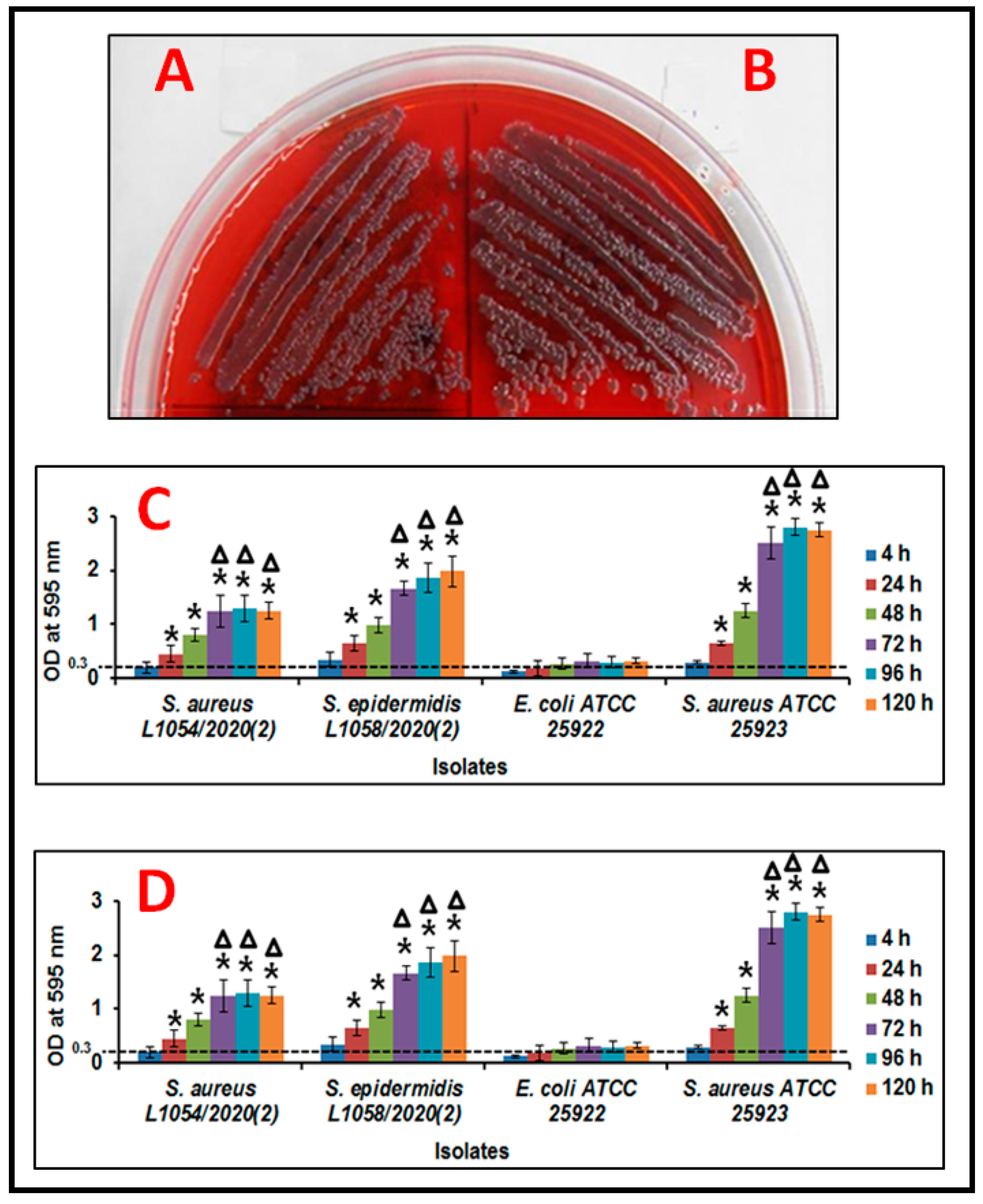
3.1. Biofilm Formation in Ocular Fluid Staphylococcus aureus and S. epidermidis
3.2. Monitoring Biofilm Formation in Ocular Fluid Staphylococcus aureus and S. epidermidis by Scanning Electron Microscopy Using Cover Slips as a Substratum
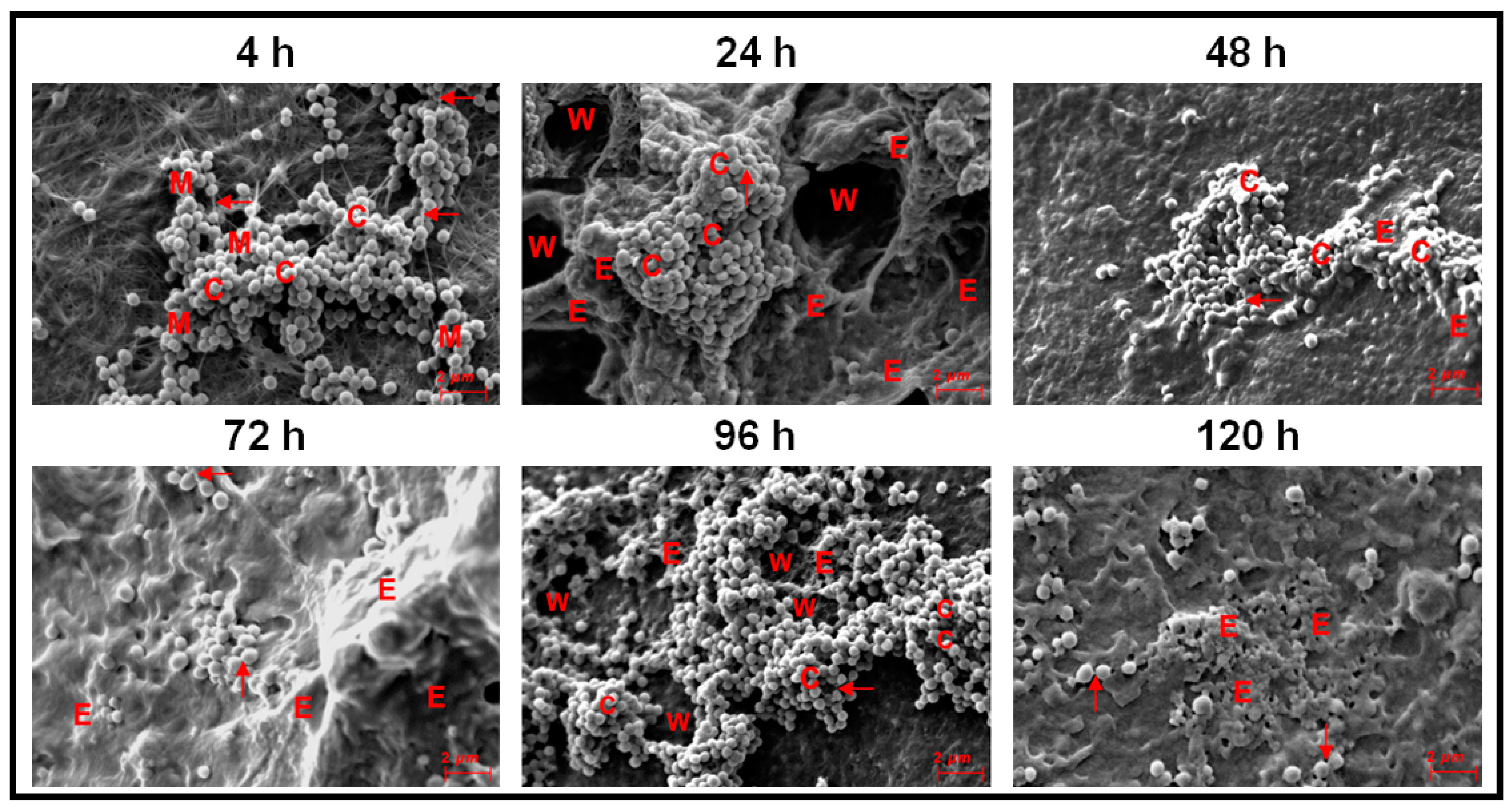
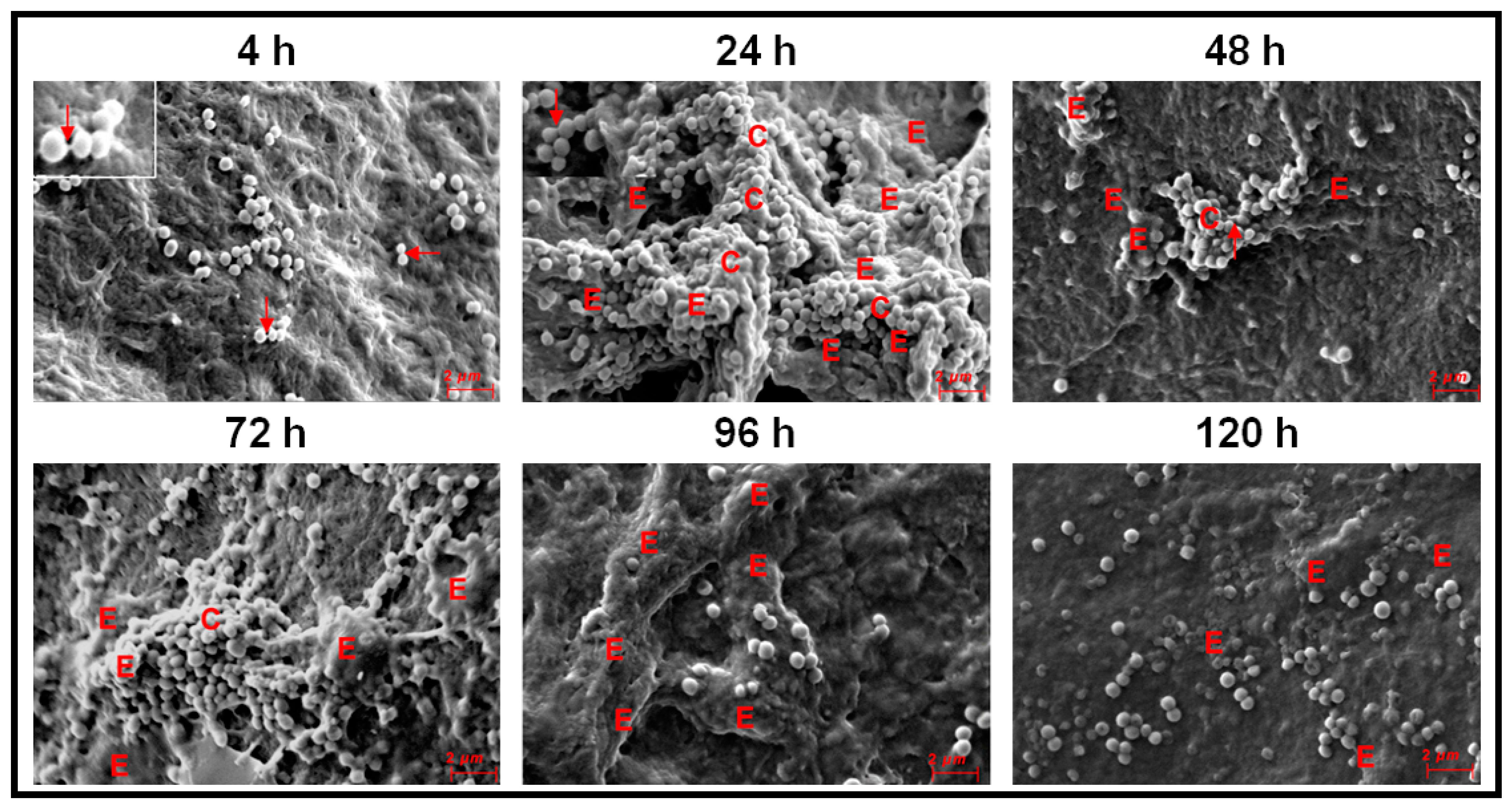
3.3. Monitoring Biofilm Formation in Ocular Fluid Staphylococcus aureus and S. epidermidis by SEM Using Cornea as a Substratum
3.4. Antibiotic Susceptibility in Ocular Staphylococcus aureus and S. epidermidis in the Planktonic and Biofilm Phase
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McClellan, K.A. Mucosal defense of the outer eye. Surv. Ophthalmol. 1997, 42, 233–246. [Google Scholar] [CrossRef]
- McDermott, A.M. Antimicrobial compounds in tears. Exp. Eye Res. 2013, 117, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Pleyer, U.; Baatz, H. Antibacterial Protection of the Ocular Surface. Ophthalmologica 1997, 211, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Shovlin, J.P.; Argüeso, P.; Carnt, N.; Chalmers, R.L.; Efron, N.; Fleiszig, S.M.J.; Nichols, J.J.; Polse, K.A.; Stapleton, F.; Wiley, L.; et al. Ocular surface health with contact lens wear. Cont. Lens Anterior Eye 2013, 36, S14–S21. [Google Scholar] [CrossRef]
- Chandler, J.W.; Cummings, M.; Gillette, T.E. Presence of Langerhans cells in the central corneas of normal human infants. Investig. Ophthalmol. Vis. Sci. 1985, 26, 113–116. [Google Scholar]
- Gillette, T.E.; Chandler, J.W.; Greiner, J.V. Langerhans Cells of the Ocular Surface. Ophthalmology 1982, 89, 700–711. [Google Scholar] [CrossRef]
- Akpek, E.K.; Gottsch, J.D. Immune defense at the ocular surface. Eye 2003, 17, 949–956. [Google Scholar] [CrossRef]
- Dong, Q.; Brulc, J.M.; Iovieno, A.; Bates, B.; Garoutte, A.; Miller, D.; Revanna, K.V.; Gao, X.; Antonopoulos, D.A.; Slepak, V.Z.; et al. Diversity of Bacteria at Healthy Human Conjunctiva. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5408–5413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, J.E.; Moore, J.E.; Jiru, X.; Moore, J.E.; Goodall, E.A.; Dooley, J.S.; Hayes, V.E.; Dartt, D.A.; Downes, C.S.; Moore, T.C. Ocular pathogen or commensal: A PCR-based study of surface bacterial flora in normal and dry eyes. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5616–5623. [Google Scholar] [CrossRef]
- Ozkan, J.; Nielsen, S.; Diez-Vives, C.; Coroneo, M.; Thomas, T.; Willcox, M. Temporal Stability and Composition of the Ocular Surface Microbiome. Sci. Rep. 2017, 7, 9880. [Google Scholar] [CrossRef] [Green Version]
- Schabereiter-Gurtner, C.; Maca, S.; Rolleke, S.; Nigl, K.; Lukas, J.; Hirschl, A.; Lubitz, W.; Barisani-Asenbauer, T. 16S rDNA-based identification of bacteria from conjunctival swabs by PCR and DGGE fingerprinting. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1164–1171. [Google Scholar] [PubMed]
- Shivaji, S.; Jayasudha, R.; Chakravarthy, S.K.; SaiAbhilash, C.R.; Sai Prashanthi, G.; Sharma, S.; Garg, P.; Murthy, S.I. Alterations in the conjunctival surface bacterial microbiome in bacterial keratitis patients. Exp. Eye Res. 2021, 203, 108418. [Google Scholar] [CrossRef]
- Shivaji, S.; Jayasudha, R.; Sai Prashanthi, G.; Kalyana Chakravarthy, S.; Sharma, S. The Human Ocular Surface Fungal Microbiome. Investig. Ophthalmol. Vis. Sci. 2019, 60, 451–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keilty, R.A. The Bacterial Flora of the Normal Conjunctiva with Comparative Nasal Culture Study. Am. J. Ophthalmol. 1930, 13, 876–879. [Google Scholar] [CrossRef]
- Bispo, P.J.M.; Melo, G.B.D.; d’Azevedo, P.A.; Höfling-Lima, A.L.; Yu, M.C.Z.; Pignatari, A.C.C. Coagulase-Negative Staphylococcus (CoNS), Staphylococcus aureus, S. epidermidis, and Streptococcus viridians cause acute postoperative endophthalmitis. Arq. Bras. Ophthalmol. 2008, 71, 617–622. [Google Scholar]
- Han, D.P.; Wisniewski, S.R.; Wilson, L.A.; Barza, M.; Vine, A.K.; Doft, B.H.; Kelsey, S.F. Spectrum and susceptibilities of microbiologic isolates in the Endophthalmitis Vitrectomy Study. Am. J. Ophthalmol. 1996, 122, 1–17. [Google Scholar] [CrossRef]
- Kresloff, M.S.; Castellarin, A.A.; Zarbin, M.A. Endophthalmitis. Surv. Ophthalmol. 1998, 43, 193–224. [Google Scholar] [CrossRef]
- Melo, G.B.; Bispo, P.J.M.; Yu, M.C.Z.; Pignatari, A.C.C.; Höfling-Lima, A.L. Microbial profile and antibiotic susceptibility of culture-positive bacterial endophthalmitis. Eye 2011, 25, 382–388. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.M.; Vedula, A.S.; Flynn, H.W., Jr.; Miller, D.; Scott, I.U.; Smiddy, W.E.; Murray, T.G.; Venkatraman, A.S. Endophthalmitis caused by Staphylococcus epidermidis: In vitro antibiotic susceptibilities and clinical outcomes. Ophthalmic Surg. Lasers Imaging 2007, 38, 446–451. [Google Scholar] [CrossRef]
- Behlau, I.; Gilmore, M.S. Microbial biofilms in ophthalmology and infectious disease. Arch. Ophthalmol. 2008, 126, 1572–1581. [Google Scholar] [CrossRef] [Green Version]
- Bispo, P.J.; Haas, W.; Gilmore, M.S. Biofilms in infections of the eye. Pathogens 2015, 4, 111–136. [Google Scholar] [CrossRef] [PubMed]
- Catalanotti, P.; Lanza, M.; Del Prete, A.; Lucido, M.; Catania, M.R.; Galle, F.; Boggia, D.; Perfetto, B.; Rossano, F. Slime-Producing Staphylococcus epidermidis and S. aureus in acute bacterial conjunctivitis in soft contact lens wearers. New Microbiol. 2005, 28, 345–354. [Google Scholar] [PubMed]
- Duggirala, A.; Kenchappa, P.; Sharma, S.; Peeters, J.K.; Ahmed, N.; Garg, P.; Das, T.; Hasnain, S.E. High-Resolution genome profiling differentiated Staphylococcus epidermidis isolated from patients with ocular infections and normal individuals. Investig. Ophthalmol. Vis. Sci. 2007, 48, 3239–3245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darouiche, R.O. Treatment of infections associated with surgical implants. N. Engl. J. Med. 2004, 350, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; dos Santos, D.M.; Lamartine de Moraes Melo Neto, C.; Luiz de Melo Moreno, A.; de Magalhães Bertoz, A.P.; Goiato, M.C. In Vitro evaluation of the effect of different disinfectants on the biofilm of Staphylococcus epidermidis and Staphylococcus aureus formed on acrylic ocular prostheses. PLoS ONE 2020, 15, e0240116. [Google Scholar] [CrossRef]
- Paharik, A.E.; Horswill, A.R. The Staphylococcal Biofilm: Adhesins, Regulation, and Host Response. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Fridkin, S.K.; Hageman, J.C.; Morrison, M.; Sanza, L.T.; Como-Sabetti, K.; Jernigan, J.A.; Harriman, K.; Harrison, L.H.; Lynfield, R.; Farley, M.M.; et al. Methicillin-resistant Staphylococcus aureus disease in three communities. N. Engl. J. Med. 2005, 352, 1436–1444. [Google Scholar] [CrossRef] [PubMed]
- Weigel, L.M.; Clewell, D.B.; Gill, S.R.; Clark, N.C.; McDougal, L.K.; Flannagan, S.E.; Kolonay, J.F.; Shetty, J.; Killgore, G.E.; Tenover, F.C. Genetic Analysis of a High-Level Vancomycin-Resistant Isolate of Staphylococcus aureus. Science 2003, 302, 1569–1571. [Google Scholar] [CrossRef]
- Costerton, J.W.; Cheng, K.J.; Geesey, G.G.; Ladd, T.I.; Nickel, J.C.; Dasgupta, M.; Marrie, T.J. Bacterial Biofilms in Nature and Disease. Annu. Rev. Microbiol. 1987, 41, 435–464. [Google Scholar] [CrossRef]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial Biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef]
- Nikolaev, Y.A.; Plakunov, V.K. Biofilm—“City of microbes” or an analogue of multicellular organisms? Microbiology 2007, 76, 125–138. [Google Scholar] [CrossRef]
- Donlan, R.M. Role of Biofilms in Antimicrobial Resistance. ASAIO J. 2000, 46, S47–S52. [Google Scholar] [CrossRef]
- Tolker-Nielsen, T.; Brinch, U.C.; Ragas, P.C.; Andersen, J.B.; Jacobsen, C.S.; Molin, S. Development and Dynamics of Pseudomonas sp. Biofilms. J. Bacteriol. 2000, 182, 6482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolker-Nielsen, T.; Molin, S. Spatial Organization of Microbial Biofilm Communities. Microb. Ecol. 2000, 40, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, Z. Structure and function of biofilms. In Evans LV, e. Biofilms: Recent Advances in Their Study and Control; Harwood Academic Publishers: Amsterdam, The Netherlands, 2000; pp. 1–17. [Google Scholar]
- Tombolini, R.; Unge, A.; Davey, M.E.; de Bruijn, F.J.; Jansson, J.K. Flow cytometric and microscopic analysis of GFP-tagged Pseudomonas fluorescens bacteria. FEMS Microbiol. Ecol. 1997, 22, 17–28. [Google Scholar] [CrossRef]
- Andersen, J.B.; Sternberg, C.; Poulsen, L.K.; Bjørn, S.P.; Givskov, M.; Molin, S. New Unstable Variants of Green Fluorescent Protein for Studies of Transient Gene Expression in Bacteria. Appl. Environ. Microbiol. 1998, 64, 2240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarwood, J.M.; Bartels, D.J.; Volper, E.M.; Greenberg, E.P. Quorum Sensing in Staphylococcus aureus Biofilms. J. Bacteriol. 2004, 186, 1838. [Google Scholar] [CrossRef] [Green Version]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Kolter, R. Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: A genetic analysis. Mol. Microbiol. 1998, 28, 449–461. [Google Scholar] [CrossRef]
- Guilhen, C.; Forestier, C.; Balestrino, D. Biofilm dispersal: Multiple elaborate strategies for dissemination of bacteria with unique properties. Mol. Microbiol. 2017, 105, 188–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, H.-S.; Otto, M. Molecular Basis of In Vivo Biofilm Formation by Bacterial Pathogens. Chem. Biol. 2012, 19, 1503–1513. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.; Zhu, L.; Chen, L.; Qiu, Y.; Wang, J. Detection Techniques for Extracellular Polymeric Substances in Biofilms: A Review. BioResources 2010, 11, 8092–8115. [Google Scholar] [CrossRef] [Green Version]
- Characklis, W.G.; Marshall, K.C. Biofilms: A Basis for an Interdisciplinary Approach; Characklis, W.G., Marshall, K.C., Eds.; Wiley: New York, NY, USA, 1990; pp. 3–15. [Google Scholar]
- Branda, S.S.; Vik, Å.; Friedman, L.; Kolter, R. Biofilms: The matrix revisited. Trends Microbiol. 2005, 13, 20–26. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- McBain, A.J. In Vitro Biofilm Models: An Overview. Adv. Appl. Microbiol. 2009, 69, 99–132. [Google Scholar] [PubMed]
- Burtseva, O.; Baulina, O.; Zaytseva, A.; Fedorenko, T.; Chekanov, K.; Lobakova, E. In vitro Biofilm Formation by Bioluminescent Bacteria Isolated from the Marine Fish Gut. Microb. Ecol. 2021, 81, 932–940. [Google Scholar] [CrossRef]
- Silva, V.O.; Soares, L.O.; Silva Júnior, A.; Mantovani, H.C.; Chang, Y.-F.; Moreira, M.A.S. Biofilm Formation on Biotic and Abiotic Surfaces in the Presence of Antimicrobials by Escherichia coli Isolates from Cases of Bovine Mastitis. Appl. Environ. Microbiol. 2014, 80, 6136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez-Manzano, J.; León-Romero, Á.; Olmo-Ruiz, C.; Bautista-Gallego, J.; Arroyo-López, F.N.; Garrido-Fernández, A.; Jiménez-Díaz, R. Biofilm formation on abiotic and biotic surfaces during Spanish style green table olive fermentation. Int. J. Food Microbiol. 2012, 157, 230–238. [Google Scholar] [CrossRef]
- Guiotti, A.M.; da Silva, E.V.F.; Catanoze, I.A.; de Carvalho, K.H.T.; Malavazi, E.M.; Goiato, M.C.; Dos Santos, D.M.; de Almeida, M.T.G. Microbiological analysis of conjunctival secretion in anophthalmic cavity, contralateral eye and ocular prosthesis of patients with maxillofacial abnormalities. Lett. Appl. Microbiol. 2018, 66, 104–109. [Google Scholar] [CrossRef]
- Paranhos, R.M.Z.F.; Batalhão, C.H.; Semprini, M.; Regalo, S.C.H.; Ito, I.Y.; Mattos, M.D.G.C.D. Evaluation of ocular prosthesis biofilm and anophthalmic cavity contamination after use of three cleansing solutions. J. Appl. Oral Sci. 2007, 15, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchhoff, L.; Arweiler-Harbeck, D.; Arnolds, J.; Hussain, T.; Hansen, S.; Bertram, R.; Buer, J.; Lang, S.; Steinmann, J.; Höing, B. Imaging studies of bacterial biofilms on cochlear implants—Bioactive glass (BAG) inhibits mature biofilm. PLoS ONE 2020, 15, e0229198. [Google Scholar] [CrossRef] [Green Version]
- Hou, W.; Sun, X.; Wang, Z.; Zhang, Y. Biofilm-Forming capacity of Staphylococcus epidermidis, Staphylococcus aureus, and Pseudomonas aeruginosa from ocular infections. Investig. Ophthalmol. Vis. Sci. 2012, 53, 5624–5631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerca, N.; Martins, S.; Cerca, F.; Jefferson, K.K.; Pier, G.B.; Oliveira, R.; Azeredo, J. Comparative assessment of antibiotic susceptibility of coagulase-negative staphylococci in biofilm versus planktonic culture as assessed by bacterial enumeration or rapid XTT colorimetry. J. Antimicrob. Chemother. 2005, 56, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Baidamshina, D.R.; Trizna, E.Y.; Holyavka, M.G.; Bogachev, M.I.; Artyukhov, V.G.; Akhatova, F.S.; Rozhina, E.V.; Fakhrullin, R.F.; Kayumov, A.R. Targeting microbial biofilms using Ficin, a nonspecific plant protease. Sci. Rep. 2017, 7, 46068. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, K.K.; Goldmann, D.A.; Pier, G.B. Use of Confocal Microscopy to Analyze the Rate of Vancomycin Penetration through Staphylococcus aureus Biofilms. Antimicrob. Chemother. 2005, 49, 2467–2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonesson, A.; Przybyszewska, K.; Eriksson, S.; Mörgelin, M.; Kjellström, S.; Davies, J.; Potempa, J.; Schmidtchen, A. Identification of bacterial biofilm and the Staphylococcus aureus derived protease, staphopain, on the skin surface of patients with atopic dermatitis. Sci. Rep. 2017, 7, 8689. [Google Scholar] [CrossRef] [PubMed]
- Cordoba, A.; Graue-Hernandez, E.O.; Bermudez-Magner, J.A.; Ramirez-Miranda, A.; Irusteta, L.; Bautista-de Lucio, V.M.; Ponce-Angulo, D.G.; Bautista-Hernandez, L.A.; Navas, A. Corneal Biofilm Plaques: A Novel Clinical Presentation. Cornea 2019, 38, 764–767. [Google Scholar] [CrossRef] [PubMed]
- Nonhoff, C.; Rottiers, S.; Struelens, M.J. Evaluation of the Vitek 2 system for identification and antimicrobial susceptibility testing of Staphylococcus spp. Clin. Microbiol. Infect. 2005, 11, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Dell, B.J. Capability of Selective Media to Detect Heat-Injured Shigella flexneri. J. Food Prot. 1990, 53, 141–144. [Google Scholar] [CrossRef]
- Ranjith, K.; Arunasri, K.; Reddy, G.S.; Adicherla, H.; Sharma, S.; Shivaji, S. Global gene expression in Escherichia coli, isolated from the diseased ocular surface of the human eye with a potential to form biofilm. Gut Pathog. 2017, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Ranjith, K.; Ramchiary, J.; Prakash, J.S.S.; Arunasri, K.; Sharma, S.; Shivaji, S. Gene Targets in Ocular Pathogenic Escherichia coli for Mitigation of Biofilm Formation to Overcome Antibiotic Resistance. Front. Microbiol. 2019, 10, 1308. [Google Scholar] [CrossRef] [Green Version]
- Freeman, D.J.; Falkiner, F.R.; Keane, C.T. New method for detecting slime production by coagulase negative staphylococci. J. Clin. Pathol. 1989, 42, 872. [Google Scholar] [CrossRef] [Green Version]
- Romero, D.; Aguilar, C.; Losick, R.; Kolter, R. Amyloid Fibers Provide Structural Integrity to Bacillus subtilis Biofilms. Proc. Natl. Acad. Sci. USA 2010, 200910560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichhardt, C.; Jacobson, A.N.; Maher, M.C.; Uang, J.; McCrate, O.A.; Eckart, M.; Cegelski, L. Congo Red Interactions with Curli-Producing E. coli and Native Curli Amyloid Fibers. PLoS ONE 2015, 10, e0140388. [Google Scholar] [CrossRef] [Green Version]
- Phuengmaung, P.; Somparn, P.; Panpetch, W.; Singkham-In, U.; Wannigama, D.L.; Chatsuwan, T.; Leelahavanichkul, A. Coexistence of Pseudomonas aeruginosa With Candida albicans Enhances Biofilm Thickness Through Alginate-Related Extracellular Matrix but Is Attenuated by N-acetyl-l-cysteine. Front. Cell Infect. Microbiol. 2020, 10, 594336. [Google Scholar] [CrossRef]
- Cruz, C.D.; Shah, S.; Tammela, P. Defining conditions for biofilm inhibition and eradication assays for Gram-positive clinical reference strains. BMC Microbiol. 2018, 18, 173. [Google Scholar] [CrossRef] [PubMed]
- Dhale, R.P.; Ghorpade, M.V.; Dharmadhikari, C.A. Comparison of various methods used to detect biofilm production of Candida species. J. Clin. Diagn. Res. 2014, 8, DC18. [Google Scholar] [CrossRef] [PubMed]
- Sabaeifard, P.; Abdi-Ali, A.; Soudi, M.R.; Dinarvand, R. Optimization of tetrazolium salt assay for Pseudomonas aeruginosa biofilm using microtiter plate method. J. Microbiol. Methods 2014, 105, 134–140. [Google Scholar] [CrossRef]
- Cowan, S.E.; Gilbert, E.; Liepmann, D.; Keasling, J.D. Commensal Interactions in a Dual-Species Biofilm Exposed to Mixed Organic Compounds. Appl. Environ. Microbiol. 2000, 66, 4481. [Google Scholar] [CrossRef] [Green Version]
- Ledeboer, N.A.; Jones, B.D. Exopolysaccharide Sugars Contribute to Biofilm Formation by Salmonella enterica serovar Typhimurium on HEp-2 Cells and Chicken Intestinal Epithelium. J. Bacteriol. 2005, 187, 3214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aquavella, J.V.; Van Horn, D.L.; Haggerty, C.J. Corneal Preservation Using M-K Medium. Am. J. Ophthalmol. 1975, 80, 791–799. [Google Scholar] [CrossRef]
- Pinnock, A.; Shivshetty, N.; Roy, S.; Rimmer, S.; Douglas, I.; MacNeil, S.; Garg, P. Ex Vivo rabbit and human corneas as models for bacterial and fungal keratitis. Graefes Arch. Clin. Exp. Ophthalmol. 2017, 255, 333–342. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Second Informational Supplement; CLSI/NCCLS Document M100-S22; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Okajima, Y.; Kobayakawa, S.; Tsuji, A.; Tochikubo, T. Biofilm formation by Staphylococcus epidermidis on intraocular lens material. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2971–2975. [Google Scholar] [CrossRef] [Green Version]
- Saraswathi, P.; Beuerman, R.W. Corneal Biofilms: From Planktonic to Microcolony Formation in an Experimental Keratitis Infection with Pseudomonas aeruginosa. Ocul. Surf. 2015, 13, 331–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Liang, Y.; Lin, S.; Chen, D.; Li, B.; Li, L.; Deng, Y. Crystal Violet and XTT Assays on Staphylococcus aureus Biofilm Quantification. Curr. Microbiol. 2016, 73, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Blumer, C.; Kleefeld, A.; Lehnen, D.; Heintz, M.; Dobrindt, U.; Nagy, G.; Michaelis, K.; Emödy, L.; Polen, T.; Rachel, R.; et al. Regulation of type 1 fimbriae synthesis and biofilm formation by the transcriptional regulator LrhA of Escherichia coli. Microbiology 2005, 151, 3287–3298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Déziel, E.; Comeau, Y.; Villemur, R. Initiation of Biofilm Formation by Pseudomonas aeruginosa 57RP Correlates with Emergence of Hyperpiliated and Highly Adherent Phenotypic Variants Deficient in Swimming, Swarming, and Twitching Motilities. J. Bacteriol. 2001, 183, 1195. [Google Scholar] [CrossRef] [Green Version]
- Büttner, H.; Mack, D.; Rohde, H. Structural basis of Staphylococcus epidermidis biofilm formation: Mechanisms and molecular interactions. Front. Cell. Infect. Microbiol. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Gross, M.; Cramton, S.E.; Götz, F.; Peschel, A. Key Role of Teichoic Acid Net Charge in Staphylococcus aureus Colonization of Artificial Surfaces. Infect. Immun. 2001, 69, 3423. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Conover, M.; Lu, H.; Parsek, M.R.; Bayles, K.; Wozniak, D.J. Assembly and Development of the Pseudomonas aeruginosa Biofilm Matrix. PLoS Pathog. 2009, 5, e1000354. [Google Scholar] [CrossRef] [Green Version]
- Pakkulnan, R.; Anutrakunchai, C.; Kanthawong, S.; Taweechaisupapong, S.; Chareonsudjai, P.; Chareonsudjai, S. Extracellular DNA facilitates bacterial adhesion during Burkholderia pseudomallei biofilm formation. PLoS ONE 2019, 14, e0213288. [Google Scholar] [CrossRef] [PubMed]
- Clarke, S.R.; Foster, S.J. Surface Adhesins of Staphylococcus aureus. Adv. Microb. Physiol. 2006, 51, 187–224. [Google Scholar] [PubMed]
- Pei, L.; Palma, M.; Nilsson, M.; Guss, B.; Flock, J.-I. Functional Studies of a Fibrinogen Binding Protein from Staphylococcus epidermidis. Infect. Immun. 1999, 67, 4525. [Google Scholar] [CrossRef] [Green Version]
- McElroy, M.C.; Cain, D.J.; Tyrrell, C.; Foster, T.J.; Haslett, C. Increased Virulence of a Fibronectin-Binding Protein Mutant of Staphylococcus aureus in a Rat Model of Pneumonia. Infect. Immun. 2002, 70, 3865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Du, X.; Villaruz, A.E.; Diep, B.A.; Wang, D.; Song, Y.; Tian, Y.; Hu, J.; Yu, F.; Lu, Y.; et al. MRSA epidemic linked to a quickly spreading colonization and virulence determinant. Nat. Med. 2012, 18, 816–819. [Google Scholar] [CrossRef] [PubMed]
- Castonguay, M.-H.; van der Schaaf, S.; Koester, W.; Krooneman, J.; van der Meer, W.; Harmsen, H.; Landini, P. Biofilm formation by Escherichia coli is stimulated by synergistic interactions and co-adhesion mechanisms with adherence-proficient bacteria. Res. Microbiol. 2006, 157, 471–478. [Google Scholar] [CrossRef]
- Baillif, S.; LeDuff, F.; Hartmann, D.J.; Kodjikian, L. Staphylococcus epidermidis Biofilm Formation and Structural Organization on Different Types of Intraocular Lenses under in vitro Flow Conditions. Ophthalmic Res. 2013, 50, 83–90. [Google Scholar] [CrossRef]
- Jassim, S.H.; Sivaraman, K.R.; Jimenez, J.C.; Jaboori, A.H.; Federle, M.J.; de la Cruz, J.; Cortina, M.S. Bacteria Colonizing the Ocular Surface in Eyes with Boston Type 1 Keratoprosthesis: Analysis of Biofilm-Forming Capability and Vancomycin Tolerance. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4689–4696. [Google Scholar] [CrossRef] [Green Version]
- Otto, M. Staphylococcal Biofilms. In Bacterial Biofilms; Romeo, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 207–228. [Google Scholar]
- Sivaraman, K.R.; Hou, J.H.; Chang, J.H.; Behlau, I.; Cortina, M.S.; De La Cruz, J. Scanning Electron Microscopic Analysis of Biofilm Formation in Explanted Human Boston Type I Keratoprostheses. Cornea 2016, 35, 25–29. [Google Scholar] [CrossRef]
- Elder, M.J.; Matheson, M.; Stapleton, F.; Dart, J.K.G. Biofilm Formation in Infectious Crystalline Keratopathy Due to Candida albicans. Cornea 1996, 15, 301–304. [Google Scholar] [CrossRef]
- Fulcher, T.P.; Dart, J.K.G.; McLaughlin-Borlace, L.; Howes, R.; Matheson, M.; Cree, I. Demonstration of biofilm in infectious crystalline keratopathy using ruthenium red and electron microscopy. Ophthalmology 2001, 108, 1088–1092. [Google Scholar] [CrossRef]
- Georgiou, T.; Qureshi, S.H.; Chakrabarty, A.; Noble, B.A. Biofilm formation and coccal organisms in infectious crystalline keratopathy. Eye 2002, 16, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Mihara, E.; Shimizu, M.; Touge, C.; Inoue, Y. Case of a large, movable bacterial concretion with biofilm formation on the ocular surface. Cornea 2004, 23, 513–515. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M.; Vaudaux, P.E.; Pittet, D.; Auckenthaler, R.; Lew, P.D.; Perdreau, F.S.; Peters, G.; Waldvogel, F.A. Fibronectin, Fibrinogen, and Laminin Act as Mediators of Adherence of Clinical Staphylococcal Isolates to Foreign Material. J. Infect. Dis. 1988, 158, 693–701. [Google Scholar] [CrossRef] [PubMed]
- McGavin, M.H.; Krajewska-Pietrasik, D.; Ryden, C.; Hook, M. Identification of a Staphylococcus aureus extracellular matrix-binding protein with broad specificity. Infect. Immun. 1993, 61, 2479–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacKintosh, E.E.; Patel, J.D.; Marchant, R.E.; Anderson, J.M. Effects of biomaterial surface chemistry on the adhesion and biofilm formation of Staphylococcus epidermidis in vitro. J. Biomed. Mater. Res. 2006, 78, 836–842. [Google Scholar] [CrossRef]
- Kaplan, J.B. Biofilm Dispersal: Mechanisms, Clinical Implications, and Potential Therapeutic Uses. J. Dent. Res. 2010, 89, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Karatan, E.; Watnick, P. Signals, Regulatory Networks, and Materials That Build and Break Bacterial Biofilms. Microbiol. Mol. Biol. Rev. 2009, 73, 310. [Google Scholar] [CrossRef] [Green Version]
- Barraud, N.; Moscoso, J.A.; Ghigo, J.-M.; Filloux, A. Methods for Studying Biofilm Dispersal in Pseudomonas aeruginosa. In Pseudomonas Methods and Protocols; Filloux, A., Ramos, J.-L., Eds.; Springer: New York, NY, USA, 2014; pp. 643–651. [Google Scholar]
- Zegans, M.E.; Becker, H.I.; Budzik, J.; O’Toole, G. The Role of Bacterial Biofilms in Ocular Infections. DNA Cell Biol. 2002, 21, 415–420. [Google Scholar] [CrossRef]
- Kobayakawa, S.; Jett, B.D.; Gilmore, M.S. Biofilm Formation by Enterococcus faecalis on Intraocular Lens Material. Curr. Eye Res. 2005, 30, 741–745. [Google Scholar] [CrossRef]
- Mah, T.-F.C.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Xu, K.D.; McFeters, G.A.; Stewart, P.S. Biofilm resistance to antimicrobial agents. Microbiology 2000, 146, 547–549. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Mendis, N.; Trigui, H.; Oliver, J.D.; Faucher, S.P. The importance of the viable but non-culturable state in human bacterial pathogens. Front. Microbiol. 2014, 5, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerdes, K.; Maisonneuve, E. Bacterial Persistence and Toxin-Antitoxin Loci. Annu. Rev. Microbiol. 2012, 66, 103–123. [Google Scholar] [CrossRef] [PubMed]
- Khan, W.; Bernier, S.P.; Kuchma, S.L.; Hammond, J.H.; Hasan, F.; O’Toole, G.A. Aminoglycoside resistance of Pseudomonas aeruginosa biofilms modulated by extracellular polysaccharide. Int. Microbiol. 2010, 13, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Billings, N.; Birjiniuk, A.; Samad, T.S.; Doyle, P.S.; Ribbeck, K. Material properties of biofilms—A review of methods for understanding permeability and mechanics. Rep. Prog. Phys. 2015, 78, 036601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilton, M.; Charron-Mazenod, L.; Moore, R.; Lewenza, S. Extracellular DNA Acidifies Biofilms and Induces Aminoglycoside Resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2016, 60, 544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjith, K.; SaiAbhilash, C.R.; Sai Prashanthi, G.; Padakandla, S.R.; Sharma, S.; Shivaji, S. Phylogenetic Grouping of Human Ocular Escherichia coli Based on Whole-Genome Sequence Analysis. Microorganisms 2020, 8, 422. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biofilm Formation (h) | S. aureus Biofilm Thickness (Z Axis, Average ± Standard Deviation in µm) | S. epidermidis Biofilm Thickness (Z Axis, Average ± Standard Deviation in µm) |
|---|---|---|
| 4 | 2.00 ± 0.25 | 2.35 ± 0.19 |
| 24 | 2.36 ± 0.23 * | 3.68 ± 0.26 * |
| 48 | 4.68 ± 0.60 * | 6.98 ± 0.45 * |
| 72 | 11.96 ± 0.90 * | 12.35 ± 0.16 * |
| 96 | 12.06 ± 0.11 * | 18.36 ± 0.21 * |
| 120 | 12.99 ± 0.46 * | 16.35 ± 0.16 * |
| Antibiotic | S. aureus MIC * (µg/mL) | S. epidermidis MIC * (µg/mL) | Fold Change ** in MIC (Planktonic vs. Biofilm Phase) | |||
|---|---|---|---|---|---|---|
| Planktonic Phase | Biofilm Phase | Planktonic Phase | Biofilm Phase | S. aureus | S. epidermidis | |
| Amikacin | 12 | 120 | 12 | 1024 | 10 | 85.3 |
| Gentamicin | 24 | 480 | 24 | 1024 | 20 | 42.7 |
| Tobramycin | 24 | 128 | 48 | 256 | 5.3 | 5.3 |
| Ampicillin | 24 | 256 | 48 | 1024 | 10.7 | 21.3 |
| Cefuroxime | 24 | 512 | 24 | 512 | 21.3 | 21.3 |
| Ceftriaxone | 12 | 512 | 12 | 512 | 42.7 | 42.7 |
| Cefepime | 48 | 1024 | 48 | 1024 | 21.3 | 21.3 |
| Cefazolin | 24 | 480 | 12 | 480 | 20 | 40 |
| Ceftazidime | 24 | 1024 | 24 | 1024 | 42.7 | 42.7 |
| Gatifloxacin | 20 | 1024 | 20 | 1024 | 51.2 | 51.2 |
| Moxifloxacin | 48 | 1024 | 48 | 1024 | 21.3 | 21.3 |
| Ciprofloxacin | 24 | 64 | 24 | 128 | 2.7 | 5.3 |
| Ofloxacin | 12 | 512 | 32 | 1024 | 42.7 | 32 |
| Vancomycin | 6 | 64 | 10 | 32 | 10.7 | 3.2 |
| Chloramphenicol | 12 | 32 | 20 | 128 | 2.7 | 6.4 |
| Azithromycin | 48 | >1054 | 128 | >1024 | 22 | 22 |
| Metronidazole | 24 | >1054 | 24 | >1024 | 22 | 22 |
| Triamcinolone | 12 | 128 | 24 | 128 | 10.6 | 5.3 |
| Deriphyllin | 6 | 64 | 6 | 256 | 10.6 | 42.6 |
| Clindamycin | 48 | >1054 | 48 | >1024 | 22 | 22 |
| Lincomycin | 24 | 512 | 32 | 1024 | 21.3 | 32 |
| Monocycline | 24 | 64 | 20 | 256 | 2.6 | 12.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konduri, R.; Saiabhilash, C.R.; Shivaji, S. Biofilm-Forming Potential of Ocular Fluid Staphylococcus aureus and Staphylococcus epidermidis on Ex Vivo Human Corneas from Attachment to Dispersal Phase. Microorganisms 2021, 9, 1124. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061124
Konduri R, Saiabhilash CR, Shivaji S. Biofilm-Forming Potential of Ocular Fluid Staphylococcus aureus and Staphylococcus epidermidis on Ex Vivo Human Corneas from Attachment to Dispersal Phase. Microorganisms. 2021; 9(6):1124. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061124
Chicago/Turabian StyleKonduri, Ranjith, Chinthala Reddy Saiabhilash, and Sisinthy Shivaji. 2021. "Biofilm-Forming Potential of Ocular Fluid Staphylococcus aureus and Staphylococcus epidermidis on Ex Vivo Human Corneas from Attachment to Dispersal Phase" Microorganisms 9, no. 6: 1124. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061124