
The Abundance of Human Milk Oligosaccharide (HMO)-Metabolizing Genes in Fecal Samples from Six-Month-Old Human Infants


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Setting
2.2. Participants
2.2.1. Study Population and Sample Collection
2.2.2. Diet Analysis
2.2.3. Sample Storage and DNA Extraction
2.3. Quantitative Real-Time PCR
2.4. Statistical Analyses
3. Results
3.1. Participant Characteristics
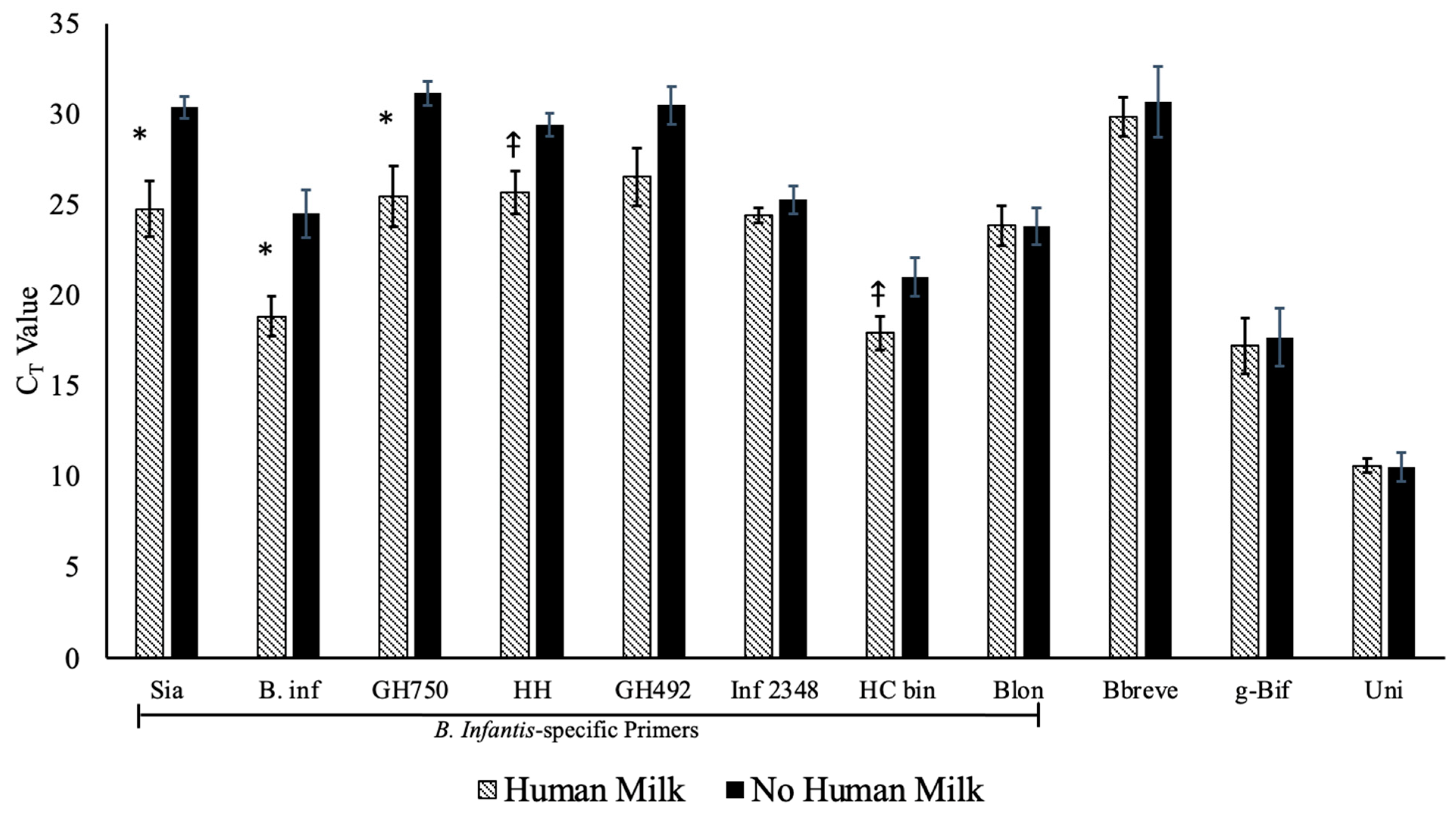
3.2. Gene Prevalence and Abundance
3.3. Gastrointestinal Bacterial Communities
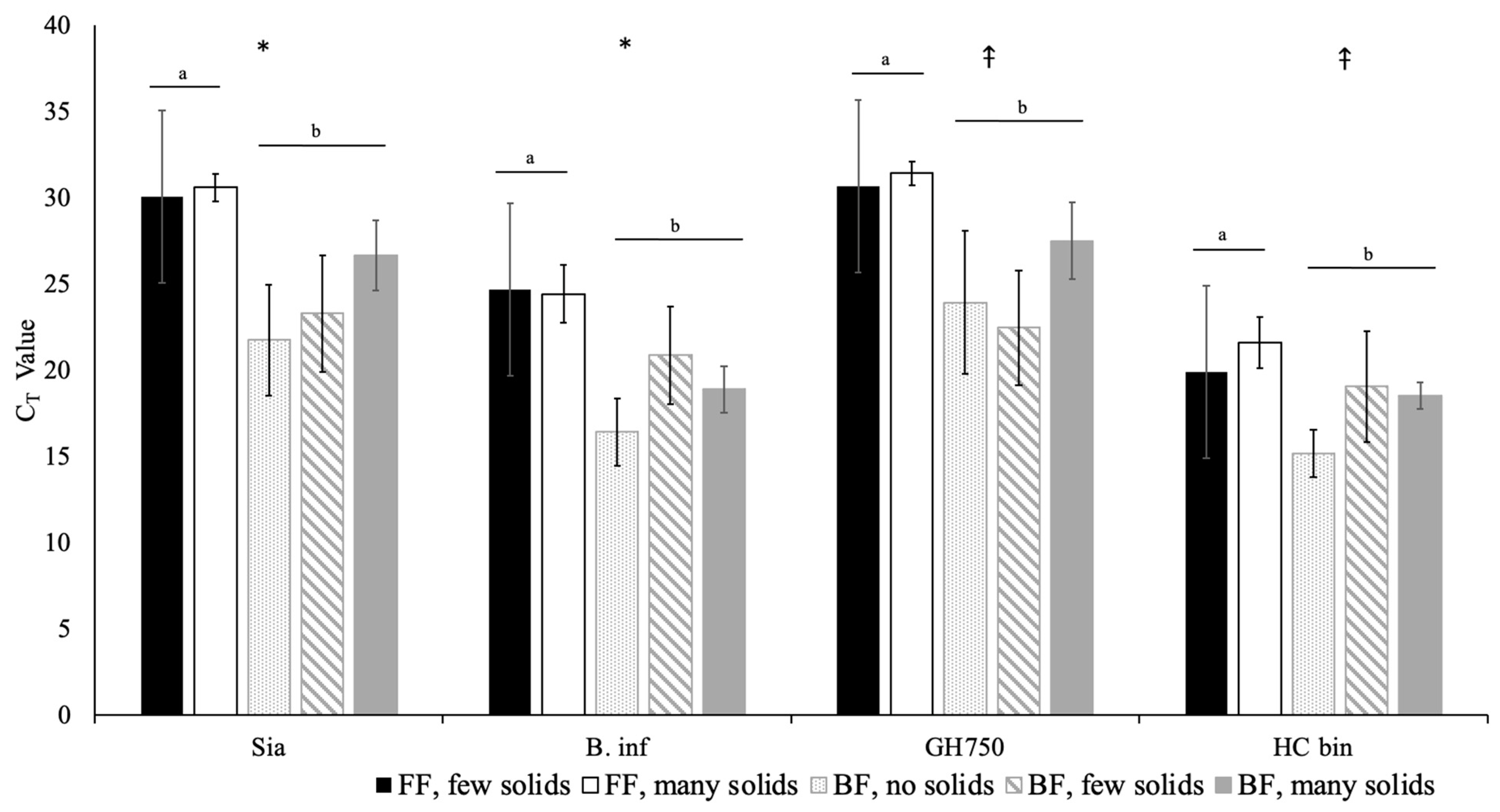
3.4. Univariate Models
3.5. Multivariate Model
4. Discussion
4.1. Limitations
4.2. Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin, C.R.; Ling, P.-R.; Blackburn, G.L. Review of Infant Feeding: Key Features of Breast Milk and Infant Formula. Nutrients 2016, 8, 279. [Google Scholar] [CrossRef] [Green Version]
- Zeuner, B.; Teze, D.; Muschiol, J.; Meyer, A.S. Synthesis of Human Milk Oligosaccharides: Protein Engineering Strategies for Improved Enzymatic Transglycosylation. Molecules 2019, 24, 2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reverri, E.J.; Devitt, A.A.; Kajzer, J.A.; Baggs, G.E.; Borschel, M.W. Review of the Clinical Experiences of Feeding Infants Formula Containing the Human Milk Oligosaccharide 2′-Fucosyllactose. Nutrients 2018, 10, 1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatric. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [Green Version]
- Hernell, O. Human Milk vs. Cow’s Milk and the Evolution of Infant Formulas. In Issues Complementary Feeding; Karger: Basel, Switzerland, 2011; pp. 17–28. [Google Scholar]
- Petherick, A. Development: Mother’s milk: A rich opportunity. Nature 2010, 468, S5–S7. [Google Scholar] [CrossRef] [PubMed]
- Donovan, S.M.; Comstock, S.S. Human Milk Oligosaccharides Influence Neonatal Mucosal and Systemic Immunity. Ann. Nutr. Metab. 2016, 69, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underwood, M.A.; Kalanetra, K.M.; Bokulich, N.A.; Mirmiran, M.; Barile, D.; Tancredi, D.J.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Prebiotic Oligosaccharides in Premature Infants. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 352–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arboleya, S.; Salazar, N.; Solís, G.; Fernández, N.; Gueimonde, M.; Reyes-Gavilán, C.G.D.L. In Vitro evaluation of the impact of human background microbiota on the response to Bifidobacterium strains and fructo-oligosaccharides. Br. J. Nutr. 2013, 110, 2030–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezirtzoglou, E. The Intestinal Microflora During the First Weeks of Life. Anaerobe 1997, 3, 173–177. [Google Scholar] [CrossRef]
- Sela, D.A.; Mills, D.A. Nursing our microbiota: Molecular linkages between bifidobacteria and milk oligosaccharides. Trends Microbiol. 2010, 18, 298–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özcan, E.; Sela, D.A. Inefficient Metabolism of the Human Milk Oligosaccharides Lacto-N-tetraose and Lacto-N-neotetraose Shifts Bifidobacterium Longum Subsp. Infantis Physiology. Front. Nutr. 2018, 5, 46. [Google Scholar] [CrossRef] [PubMed]
- Marcobal, A.; Sonnenburg, J.L. Human milk oligosaccharide consumption by intestinal microbiota. Clinical microbiology and infection: The official publication of the European Society of Clinical Microbiology and Infectious Diseases. Clin. Microbiol. Infect. 2012, 18, 12–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugino, K.Y.; Ma, T.; Kerver, J.M.; Paneth, N.; Comstock, S.S. Human Milk Feeding Patterns at 6 Months of Age are a Major Determinant of Fecal Bacterial Diversity in Infants. J. Hum. Lact. 2020, 890334420957571. [Google Scholar] [CrossRef] [PubMed]
- Bonuck, K.A.; Trombley, M.; Freeman, B.K.; McKee, D. Randomized, Controlled Trial of a Prenatal and Postnatal Lactation Consultant Intervention on Duration and Intensity of Breastfeeding up to 12 Months. Pediatrics 2005, 116, 1413–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugino, K.Y.; Paneth, N.; Comstock, S.S. Michigan cohorts to determine associations of maternal pre-pregnancy body mass index with pregnancy and infant gastrointestinal microbial communities: Late pregnancy and early infancy. PLoS ONE 2019, 14, e0213733. [Google Scholar] [CrossRef] [Green Version]
- Tso, L. Targeted high-resolution taxonomic identification of Bifidobacterium Longum Subsp. Infantis using human milk oligosaccharide metabolizing genes. bioRxiv 2021. [Google Scholar] [CrossRef]
- Tannock, G.W. Comparison of the compositions of the stool microbiotas of infants fed goat milk formula, cow milk-based formula, or breast milk. Appl. Environ. Microbiol. 2013, 79, 3040–3048. [Google Scholar] [CrossRef] [Green Version]
- Lawley, B. Differentiation of Bifidobacterium Longum subspecies Longum and Infantis by quantitative PCR using functional gene targets. Peer J 2017, 5, e3375. [Google Scholar] [CrossRef] [Green Version]
- Hong, W.-S.; Chen, M.-J. Rapid Identification of Bifidobacteria in Dairy Products by Gene-targeted Species-specific PCR Technique and DGGE. Asian-Australas. J. Anim. Sci. 2007, 20, 1887–1894. [Google Scholar] [CrossRef]
- Tannock, G.W. Identification of Lactobacillus isolates from the gastrointestinal tract, silage, and yoghurt by 16S-23S rRNA gene intergenic spacer region sequence comparisons. Appl. Environ. Microbiol. 1999, 65, 4264–4267. [Google Scholar] [CrossRef] [Green Version]
- Frese, S.A. Persistence of supplemented Bifidobacterium longum subsp infantis EVC001 in breasted infants. mSphere 2017, 2, e00501-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuki, T. Rapid identification of human intestinal bifidobacteria by 16S rRNA-targeted species- and group-specific primers. FEMS Microbiol. Lett. 1998, 167, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Grönlund, M.M. Fecal microflora in healthy infants born by different methods of delivery: Permanent changes in intestinal flora after cesarean delivery. J. Pediatric. Gastroenterol. Nutr. 1999, 28, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Hesla, H.M. Impact of lifestyle on the gut microbiota of healthy infants and their mothers—The ALADDIN birth cohort. FEMS Microbiol. Ecol. 2014, 90, 791–801. [Google Scholar] [CrossRef] [Green Version]
- Dogra, S.; Sakwinska, O.; Soh, S.-E.; Ngom-Bru, C.; Brück, W.M.; Berger, B.; Brüssow, H.; Lee, Y.S.; Yap, F.; Chong, Y.-S.; et al. Dynamics of Infant Gut Microbiota Are Influenced by Delivery Mode and Gestational Duration and Are Associated with Subsequent Adiposity. mBio 2015, 6, e02419-14. [Google Scholar] [CrossRef] [Green Version]
- Huurre, A.; Kalliomäki, M.; Rautava, S.; Rinne, M.; Salminen, S.; Isolauri, E. Mode of Delivery Effects on Gut Microbiota and Humoral Immunity. Neonatology 2008, 93, 236–240. [Google Scholar] [CrossRef]
- Mitsou, E.K.; Kirtzalidou, E.; Oikonomou, I.; Liosis, G.; Kyriacou, A. Fecal microflora of Greek healthy neonates. Anaerobe 2008, 14, 94–101. [Google Scholar] [CrossRef]
- Ebihara, K.; Tachibe, M.; Kaneko, N.; Kishida, T.; Kabeerdoss, J.; Ferdous, S.; Balamurugan, R.; Mechenro, J.; Vidya, R.; Santhanam, S.; et al. Hydroxypropylation of high-amylose maize starch changes digestion and fermentation-dependent parameters in rats Development of the gut microbiota in southern Indian infants from birth to 6 months: A molecular analysis ERRATUM. J. Nutr. Sci. 2013, 2, 18. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.J.; Ajami, N.J.; O’Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nat. Cell Biol. 2018, 562, 583–588. [Google Scholar] [CrossRef]
- Ferretti, P.; Pasolli, E.; Tett, A.; Asnicar, F.; Gorfer, V.; Fedi, S.; Armanini, F.; Truong, D.T.; Manara, S.; Zolfo, M.; et al. Mother-to-Infant Microbial Transmission from Different Body Sites Shapes the Developing Infant Gut Microbiome. Cell Host Microbe 2018, 24, 133–145.e5. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [Green Version]
- Korpela, K.; Helve, O.; Kolho, K.-L.; Saisto, T.; Skogberg, K.; Dikareva, E.; Stefanovic, V.; Salonen, A.; Andersson, S.; de Vos, W.M. Maternal Fecal Microbiota Transplantation in Cesarean-Born Infants Rapidly Restores Normal Gut Microbial Development: A Proof-of-Concept Study. Cell 2020, 183, 324–334. [Google Scholar] [CrossRef]
- Lawson, M.A.E.; O’Neill, I.; Kujawska, M.; Javvadi, S.G.; Wijeyesekera, A.; Flegg, Z.; Chalklen, L.; Hall, L.J. Breast milk-derived human milk oligosaccharides promote Bifidobacterium interactions within a single ecosystem. ISME J. 2019, 14, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Sakanaka, M.; Gotoh, A.; Yoshida, K.; Odamaki, T.; Koguchi, H.; Xiao, J.-Z.; Kitaoka, M.; Katayama, T. Varied Pathways of Infant Gut-Associated Bifidobacterium to Assimilate Human Milk Oligosaccharides: Prevalence of the Gene Set and Its Correlation with Bifidobacteria-Rich Microbiota Formation. Nutrients 2019, 12, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, A.L.; Alves, R.; Figueiredo, A.; Alves-Santos, N.; Freitas-Costa, N.; Batalha, M.; Yonemitsu, C.; Manivong, N.; Furst, A.; Bode, L.; et al. Human Milk Oligosaccharide Profile Variation Throughout Postpartum in Healthy Women in a Brazilian Cohort. Nutrients 2020, 12, 790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuel, T.M.; Binia, A.; De Castro, C.A.; Thakkar, S.K.; Billeaud, C.; Agosti, M.; Al-Jashi, I.; Costeira, M.J.; Marchini, G.; Martínez-Costa, C.; et al. Impact of maternal characteristics on human milk oligosaccharide composition over the first 4 months of lactation in a cohort of healthy European mothers. Sci. Rep. 2019, 9, 11767. [Google Scholar] [CrossRef] [PubMed]
- Alderete, T.L.; Autran, C.; Brekke, B.E.; Knight, R.; Bode, L.; Goran, M.I.; Fields, D.A. Associations between human milk oligosaccharides and infant body composition in the first 6 mo of life. Am. J. Clin. Nutr. 2015, 102, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Garrido, D.; Dallas, D.C.; Mills, D.A. Consumption of human milk glycoconjugates by infant-associated bifidobacteria: Mechanisms and implications. Microbiology 2013, 159, 649–664. [Google Scholar] [CrossRef] [PubMed]
- Dessì, A.; Marincola, F.C.; Masili, A.; Gazzolo, D.; Fanos, V. Clinical Metabolomics and Nutrition: The New Frontier in Neonatology and Pediatrics. BioMed. Res. Int. 2014, 2014, 1–8. [Google Scholar] [CrossRef]
- Brink, L.R.; Mercer, K.E.; Piccolo, B.D.; Chintapalli, S.V.; Elolimy, A.; Bowlin, A.K.; Matazel, K.S.; Pack, L.; Adams, S.H.; Shankar, K.; et al. Neonatal diet alters fecal microbiota and metabolome profiles at different ages in infants fed breast milk or formula. Am. J. Clin. Nutr. 2020, 111, 1190–1202. [Google Scholar] [CrossRef]
- Laurens, M.L.L.; Kraus-Friedberg, C.; Kar, W.; Sanfilippo, D.; Rajasekaran, S.; Comstock, S.S. Dietary Intake Influences Metabolites in Healthy Infants: A Scoping Review. Nutrients 2020, 12, 2073. [Google Scholar] [CrossRef] [PubMed]
- Jian, C.; Luukkonen, P.; Yki-Järvinen, H.; Salonen, A.; Korpela, K. Quantitative PCR provides a simple and accessible method for quantitative microbiota profiling. PLoS ONE 2020, 15, e0227285. [Google Scholar] [CrossRef] [Green Version]
- Kralik, P.; Ricchi, M. A Basic Guide to Real Time PCR in Microbial Diagnostics: Definitions, Parameters, and Everything. Front. Microbiol. 2017, 8, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezirtzoglou, E.; Maipa, V.; Chotoura, N.; Apazidou, E.; Tsiotsias, A.; Voidarou, C.; Kostakis, D.; Alexopoulos, A. Occurrence of Bifidobacterium in the intestine of newborns by fluorescence in situ hybridization. Comp. Immunol. Microbiol. Infect. Dis. 2006, 29, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Bezirtzoglou, E.; Tsiotsias, A.; Welling, G.W. Microbiota profile in feces of breast- and formula-fed newborns by using fluorescence in situ hybridization (FISH). Anaerobe 2011, 17, 478–482. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primer Set | Forward and Reverse Primers (5′–3′) | Gene Targeted | Organism Targeted | Annealing Temperature (°C) | Primer Concentration | Reference |
|---|---|---|---|---|---|---|
| Sia | F: GACGAGGAGGAATACAGCAG R: CACGAACAGCGAATCATGGATT | Sialidase (Blon_2348) | B. longum infantis | 58 | 1 μM | Klepac-Ceraj (unpublished) |
| B. inf | F: CCATCTCTGGGATCGTCGG R: TATCGGGGAGCAAGCGTGA | 16S rRNA | B. longum infantis, B. longum longum, B. indicum | 57 | 300 nM | Tannock et al. (2013) [19] |
| g-Bif | F: CTCCTGGAAACGGGTGG R: GGTGTTCTTCCCGATATCTACA | 16S rRNA | Bifidobacterium genus | 55 | 1 μM | Tannock et al. (2013) [19] |
| GH-492 | F: CGATGATGTGCTGGATTCGTTC R: CTCGACCATTCCAAGATGCTA | Glycoside hydrolase (Blon_2358) | B. longum infantis | 60 | 300 nM | Klepac-Ceraj (unpublished) |
| Inf 2348 | F: ATACAGCAGAACCTTGGCCT R: GTTCTCGTCCATGTGATCGC | Sialidase | B. longum infantis | 60 | 5 μM | Lawley et al. (2017) [20] |
| HC bin | F: AGGATACGTTCGGCGTC R: CGCAAGATTCCTCTAGCA | 16S rRNA | B. longum infantis | 60 | 5 μM | Hong and Chen (2007) [21] |
| Uni | F349: ACTCCTACGGGAGGCAGCAGT R528:ATTACCGCGGCTGCTGGC | 16S rRNA | Targets all bacteria | 60 | 1 μM | Tannock et al. (1999) [22] |
| GH750 | F: GCGCCATCCTGGTGATGTTATT R: CTACGTGATCTGGGAGAGTTTC | Glycoside hydrolase (Blon_2355) | B. longum infantis | 59 | 5 μM | Klepac-Ceraj (unpublished) |
| HH | F: CCACAATGTCATCGACCATCTG R: CCGAAGTATTCGGATGCCTATG | Haloacid dehalogenase-like hydrolase domain-containing protein (Blon_2356) | B. longum infantis | 59 | 5 μM | Klepac-Ceraj (unpublished) |
| Blon | F: CGTATTGGCTTTGTACGCATTT R: ATCGTGCCGGTGAGATTTAC | Major facilitator superfamily | B. longum infantis | 50 | 1 μM | Frese et al. (2017) [23] |
| Bbreve | F: CCGGATGTCCATCACAC R: ACAAAGTGCCTTGCTCCCT | 16S rRNA | Bifidobacterium breve | 55 | 5 μM | Matsuki et al. (1998) [24] |
| Total n = 40 | |
|---|---|
| Maternal age, years | 31.79 (min, 22; max, 39) |
| Vaginal birth, % (n) | 62.5 (25) |
| Infant age, days | 202.53 (min, 161; max, 292) |
| Male, % (n) | 70 (28) |
| Human milk exposure, % (n) | |
| 100% | 17.5 (7) |
| 20–80% | 27.5 (11) |
| >0 but <20% | 25 (10) |
| 0% | 30 (12) |
| Human milk, new | |
| Any HM, % | 70 (28) |
| No HM, % | 30 (12) |
| Solid food intake, % (n), yes | |
| No solids | 15 (6) |
| Some solids | 27.5 (11) |
| Significant solids | 57.5 (23) |
| Solid food intake, food-specific, % (n), yes | |
| Rice | 27.5 (11) |
| Fruit | 57.5 (23) |
| Vegetables | 60 (24) |
| Meat | 5 (2) |
| Oats | 20 (8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferro, L.E.; Sugino, K.Y.; Klepac-Ceraj, V.; Comstock, S.S. The Abundance of Human Milk Oligosaccharide (HMO)-Metabolizing Genes in Fecal Samples from Six-Month-Old Human Infants. Microorganisms 2021, 9, 1352. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071352
Ferro LE, Sugino KY, Klepac-Ceraj V, Comstock SS. The Abundance of Human Milk Oligosaccharide (HMO)-Metabolizing Genes in Fecal Samples from Six-Month-Old Human Infants. Microorganisms. 2021; 9(7):1352. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071352
Chicago/Turabian StyleFerro, Lynn E., Kameron Y. Sugino, Vanja Klepac-Ceraj, and Sarah S. Comstock. 2021. "The Abundance of Human Milk Oligosaccharide (HMO)-Metabolizing Genes in Fecal Samples from Six-Month-Old Human Infants" Microorganisms 9, no. 7: 1352. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071352