
The Two-Component System 09 of Streptococcus pneumoniae Is Important for Metabolic Fitness and Resistance during Dissemination in the Host

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
- μ = growth rate
- X (t1) = cell density at time t1
- X (t2) = cell density at time t2
- g = doubling time
2.2. Generation of Pneumococcal Mutants
2.3. Immunoblot Analysis of Whole Bacterial Lysates Using the LI-COR Technology
2.4. RNA Purification and Quantitative Real-Time PCR (qPCR)
2.5. Field Emission Scanning Electron Microscopy (FESEM) and Transmission Electron Microscopy (TEM)
2.6. Infection of Epithelial Cells for Pneumococcal Adherence Analysis by Immunofluorescence Microscopy
2.7. Quantification of Phagocytosed Pneumococci by the Antibiotic Protection Assay
2.8. Quantification of Phagocyte-Associated and Intracellular Pneumococci by Immunofluorescence Microscopy
2.9. Acute Pneumonia Infection Model
2.10. Systemic Infection Model
2.11. Co-Infection Model
2.12. Statistical Analysis
3. Results
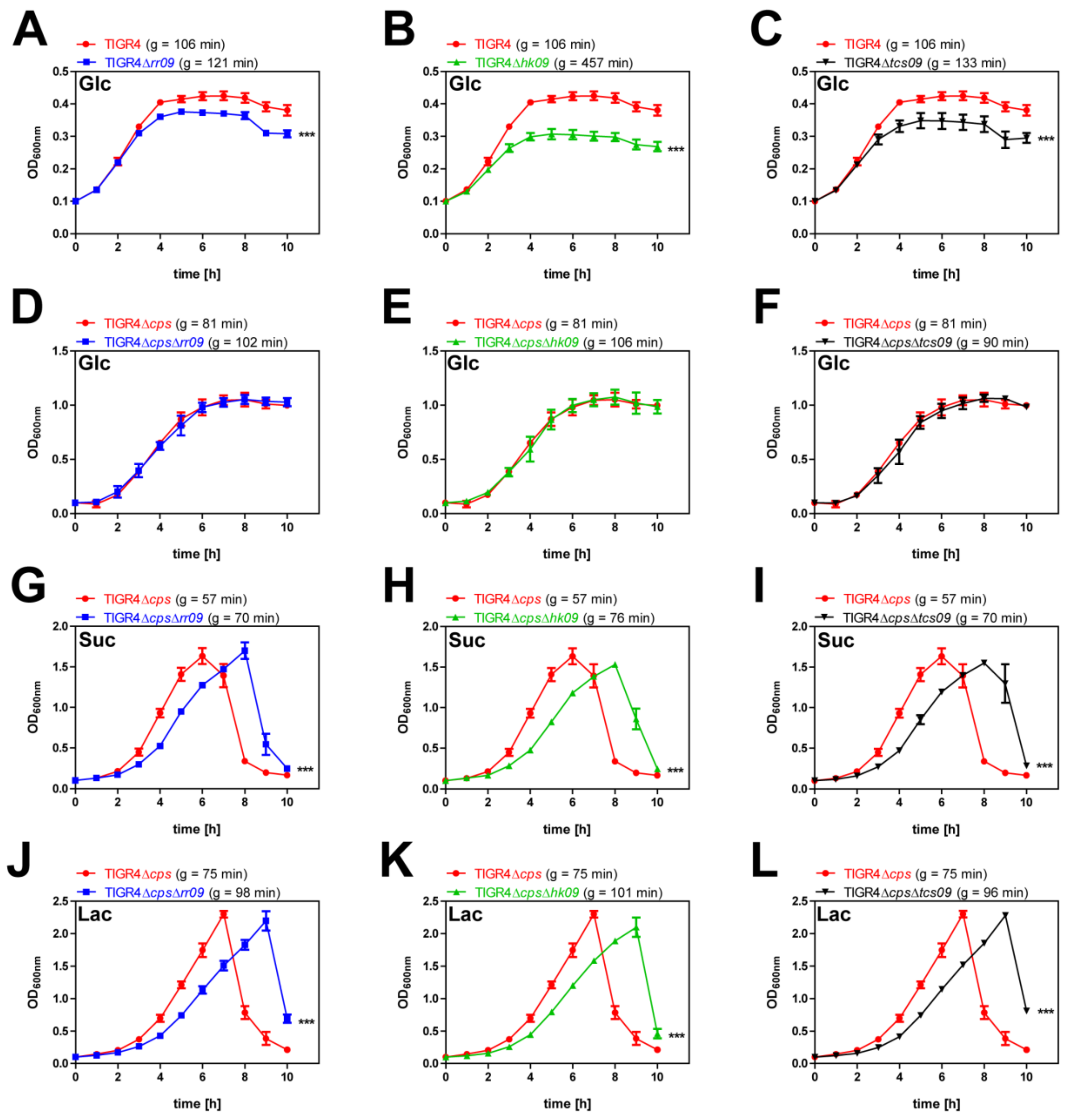
3.1. Growth Analysis of Encapsulated and Nonencapsulated Wild-Type TIGR4 and tcs09-Mutants under Nutrient-Defined Conditions
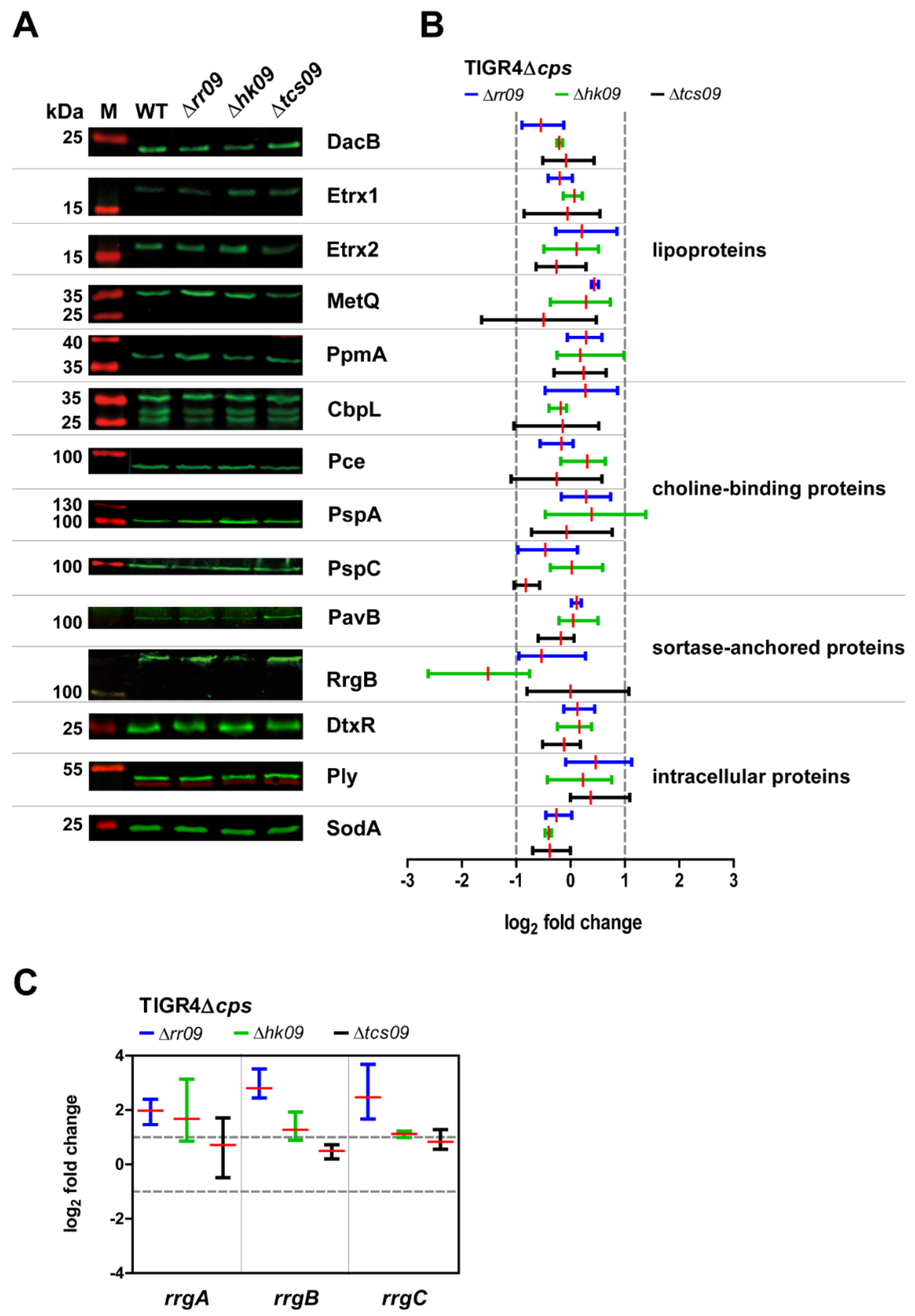
3.2. Expression of Important Virulence Factors in TIGR4Δcps Wild-Type and Isogenic tcs09-Mutants
3.3. Analysis of Pneumococcal Morphology and Capsule Content
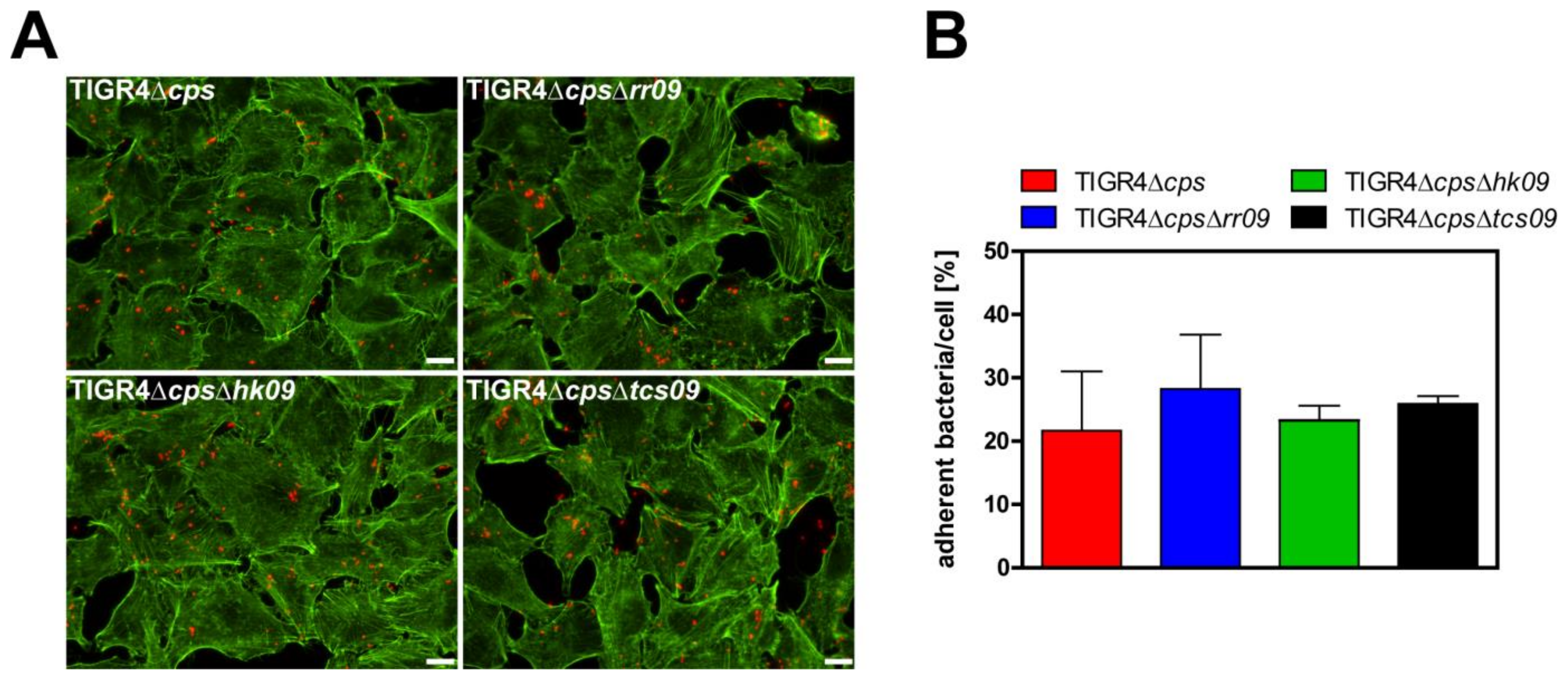
3.4. Role of the TCS09 on Pneumococcal Adherence
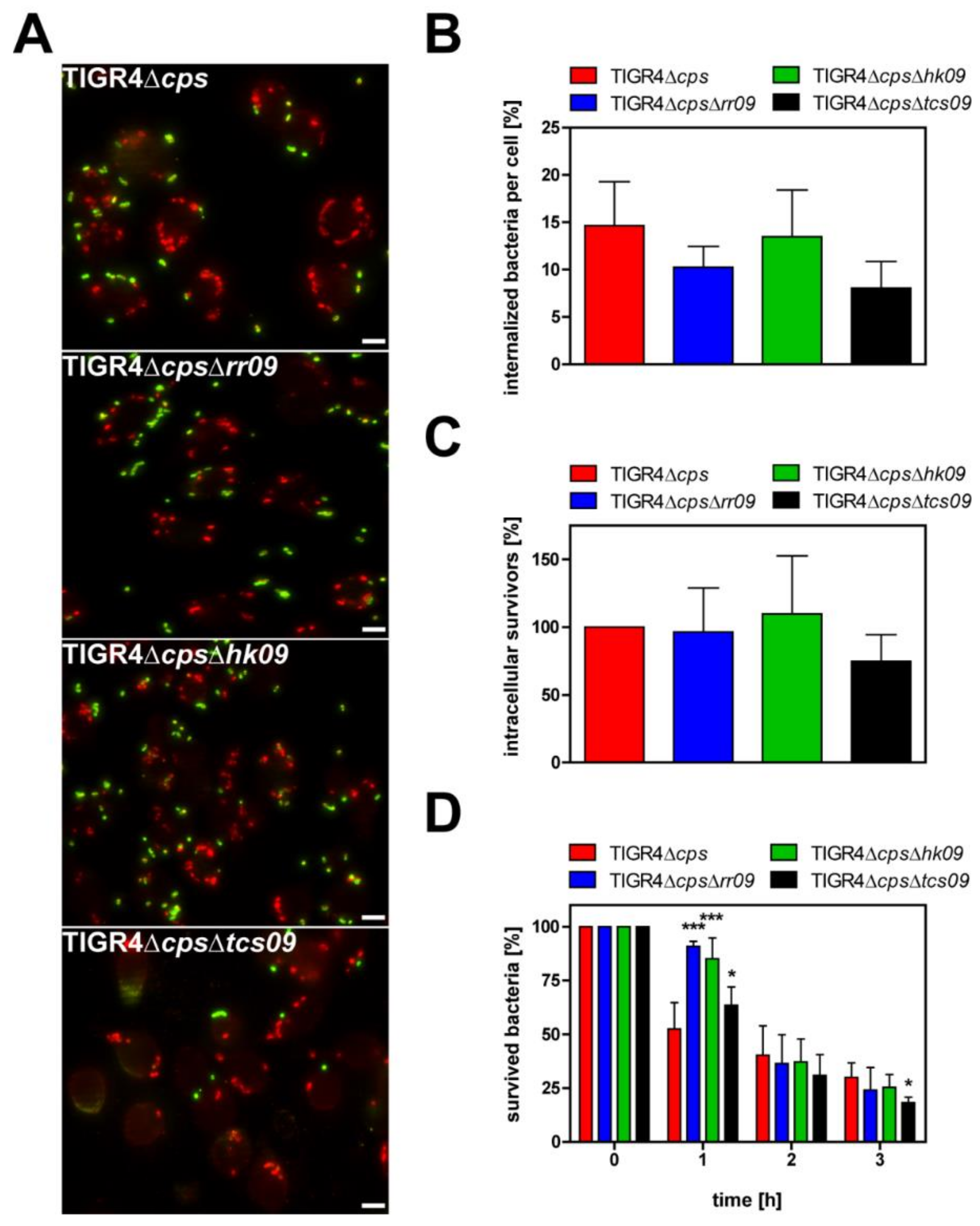
3.5. Impact of the Pneumococcal TCS09 on Uptake by Phagocytes
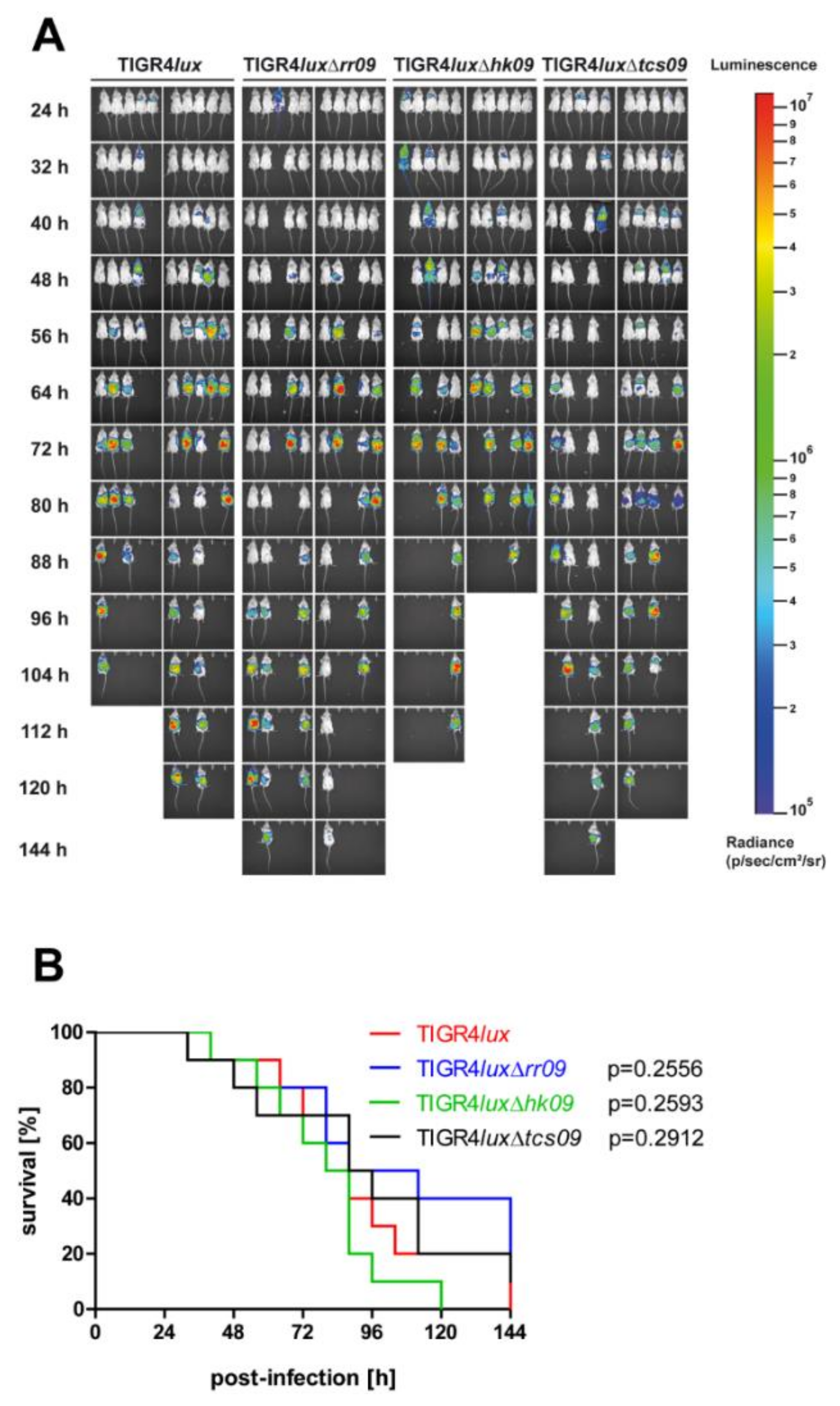
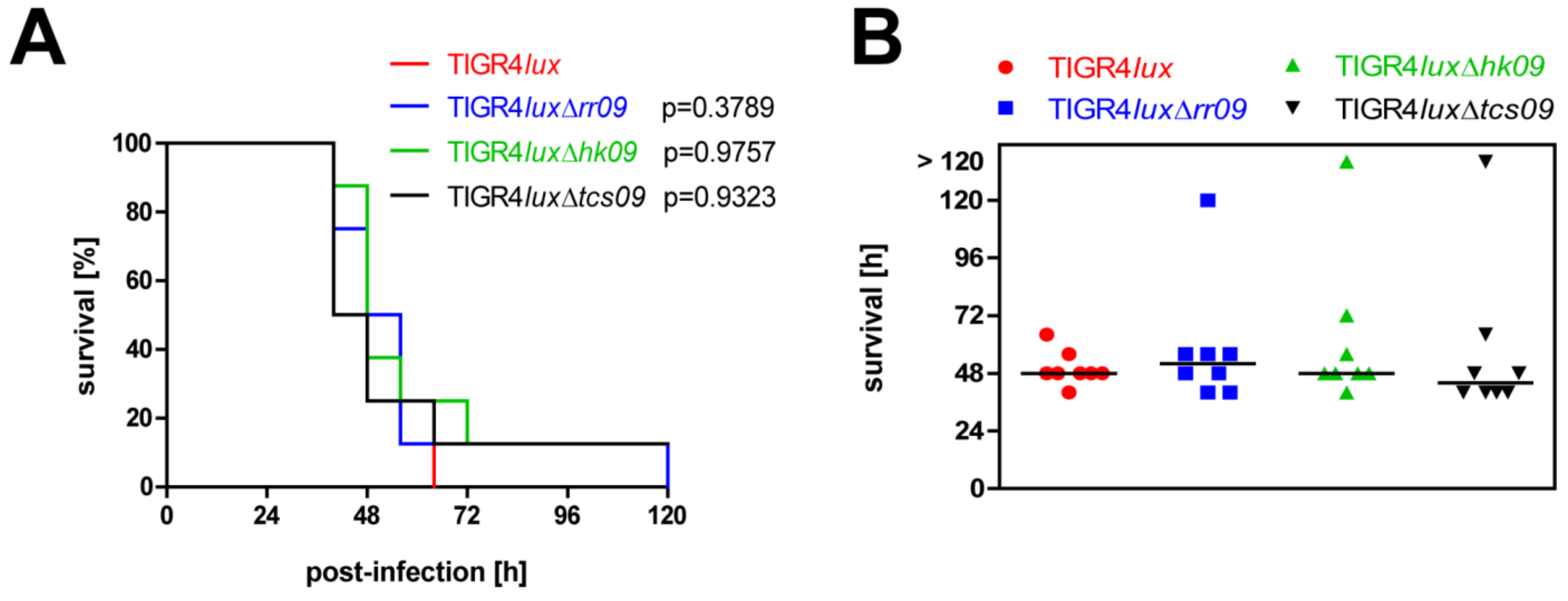
3.6. Influence of TCS09 on Lung Infections Caused by S. pneumoniae TIGR4
3.7. Impact of TCS09 on Virulence in the Systemic Mouse Infection Model
3.8. Importance of TCS09 for the Bacterial Load in the Respiratory Tract after Co-Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Statement
References
- Austrian, R. Some aspects of the pneumococcal carrier state. J. Antimicrob. Chemother. 1986, 18 (Suppl. A), 35–45. [Google Scholar] [CrossRef]
- Bogaert, D.; De Groot, R.; Hermans, P.W. Streptococcus pneumoniae colonisation: The key to pneumococcal disease. Lancet Infect. Dis. 2004, 4, 144–154. [Google Scholar] [CrossRef]
- Kadioglu, A.; Weiser, J.N.; Paton, J.C.; Andrew, P.W. The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat. Rev. Microbiol. 2008, 6, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Tuomanen, E.I.; Austrian, R.; Masure, H.R. Pathogenesis of pneumococcal infection. N. Engl. J. Med. 1995, 332, 1280–1284. [Google Scholar] [CrossRef] [PubMed]
- Wahl, B.; O’Brien, K.L.; Greenbaum, A.; Majumder, A.; Liu, L.; Chu, Y.; Lukšić, I.; Nair, H.; McAllister, D.A.; Campbell, H.; et al. Burden of Streptococcus pneumoniae and Haemophilus influenzae type b disease in children in the era of conjugate vaccines: Global, regional, and national estimates for 2000-15. Lancet Glob. Health 2018, 6, e744–e757. [Google Scholar] [CrossRef] [Green Version]
- Cooper, V.S.; Honsa, E.; Rowe, H.; Deitrick, C.; Iverson, A.R.; Whittall, J.J.; Neville, S.L.; McDevitt, C.A.; Kietzman, C.; Rosch, J.W. Experimental Evolution In Vivo To Identify Selective Pressures during Pneumococcal Colonization. mSystems 2020, 5, e00352-20. [Google Scholar] [CrossRef]
- Rowe, H.M.; Karlsson, E.; Echlin, H.; Chang, T.-C.; Wang, L.; van Opijnen, T.; Pounds, S.B.; Schultz-Cherry, S.; Rosch, J.W. Bacterial Factors Required for Transmission of Streptococcus pneumoniae in Mammalian Hosts. Cell Host Microbe 2019, 25, 884–891.e6. [Google Scholar] [CrossRef] [PubMed]
- Bermpohl, D.; Halle, A.; Freyer, D.; Dagand, E.; Braun, J.S.; Bechmann, I.; Schröder, N.W.; Weber, J.R. Bacterial programmed cell death of cerebral endothelial cells involves dual death pathways. J. Clin. Investig. 2005, 115, 1607–1615. [Google Scholar] [CrossRef] [Green Version]
- Iovino, F.; Seinen, J.; Henriques-Normark, B.; van Dijl, J.M. How Does Streptococcus pneumoniae Invade the Brain? Trends Microbiol. 2016, 24, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.R.; Tuomanen, E.I. Cellular damage in bacterial meningitis: An interplay of bacterial and host driven toxicity. J. Neuroimmunol. 2007, 184, 45–52. [Google Scholar] [CrossRef]
- Hava, D.L.; Camilli, A. Large-scale identification of serotype 4 Streptococcus pneumoniae virulence factors. Mol. Microbiol. 2002, 45, 1389–1406. [Google Scholar] [CrossRef] [Green Version]
- Kohler, S.; Voß, F.; Gómez Mejia, A.; Brown, J.S.; Hammerschmidt, S. Pneumococcal lipoproteins involved in bacterial fitness, virulence, and immune evasion. FEBS Lett. 2016, 590, 3820–3839. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Mejia, A.; Gámez, G.; Hirschmann, S.; Kluger, V.; Rath, H.; Böhm, S.; Voss, F.; Kakar, N.; Petruschka, L.; Völker, U.; et al. Pneumococcal Metabolic Adaptation and Colonization Are Regulated by the Two-Component Regulatory System 08. mSphere 2018, 3, e00165-18. [Google Scholar] [CrossRef]
- Schulz, C.; Gierok, P.; Petruschka, L.; Lalk, M.; Mäder, U.; Hammerschmidt, S. Regulation of the Arginine Deiminase System by ArgR2 Interferes with Arginine Metabolism and Fitness of Streptococcus pneumoniae. mBio 2014, 5, e01858-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, F.; Kakar, N.; Meyer, T.C.; Depke, M.; Masouris, I.; Burchhardt, G.; Gómez-Mejia, A.; Dhople, V.; Håvarstein, L.S.; Sun, Z.; et al. In vivo proteomics identifies the competence regulon and AliB oligopeptide transporter as pathogenic factors in pneumococcal meningitis. PLoS Pathog. 2019, 15, e1007987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, S.; Hammerschmidt, S. Versatility of pneumococcal surface proteins. Microbiology (Read.) 2006, 152, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Mejia, A.; Gámez, G.; Hammerschmidt, S. Streptococcus pneumoniae two-component regulatory systems: The interplay of the pneumococcus with its environment. Int. J. Med. Microbiol. 2018, 308, 722–737. [Google Scholar] [CrossRef]
- Laub, M.T.; Goulian, M. Specificity in two-component signal transduction pathways. Annu. Rev. Genet. 2007, 41, 121–145. [Google Scholar] [CrossRef] [Green Version]
- Stock, A.M.; Robinson, V.L.; Goudreau, P.N. Two-component signal transduction. Annu. Rev. Biochem. 2000, 69, 183–215. [Google Scholar] [CrossRef] [Green Version]
- Glanville, D.G.; Han, L.; Maule, A.F.; Woodacre, A.; Thanki, D.; Abdullah, I.T.; Morrissey, J.A.; Clarke, T.B.; Yesilkaya, H.; Silvaggi, N.R.; et al. RitR is an archetype for a novel family of redox sensors in the streptococci that has evolved from two-component response regulators and is required for pneumococcal colonization. PLoS Pathog. 2018, 14, e1007052. [Google Scholar] [CrossRef]
- Paterson, G.K.; Blue, C.E.; Mitchell, T.J. Role of two-component systems in the virulence of Streptococcus pneumoniae. J. Med. Microbiol. 2006, 55, 355–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinoso-Vizcaíno, N.M.; Cian, M.B.; Cortes, P.R.; Olivero, N.B.; Hernandez-Morfa, M.; Piñas, G.E.; Badapanda, C.; Rathore, A.; Perez, D.R.; Echenique, J. The pneumococcal two-component system SirRH is linked to enhanced intracellular survival of Streptococcus pneumoniae in influenza-infected pulmonary cells. PLoS Pathog. 2020, 16, e1008761. [Google Scholar] [CrossRef]
- Hirschmann, S.; Gómez-Mejia, A.; Mäder, U.; Karsunke, J.; Driesch, D.; Rohde, M.; Häussler, S.; Burchhardt, G.; Hammerschmidt, S. The Two-Component System 09 Regulates Pneumococcal Carbohydrate Metabolism and Capsule Expression. Microorganisms 2021, 9, 468. [Google Scholar] [CrossRef]
- Blue, C.E.; Mitchell, T.J. Contribution of a response regulator to the virulence of Streptococcus pneumoniae is strain dependent. Infect. Immun. 2003, 71, 4405–4413. [Google Scholar] [CrossRef] [Green Version]
- Hendriksen, W.T.; Silva, N.; Bootsma, H.J.; Blue, C.E.; Paterson, G.K.; Kerr, A.R.; de Jong, A.; Kuipers, O.P.; Hermans, P.W.; Mitchell, T.J. Regulation of gene expression in Streptococcus pneumoniae by response regulator 09 is strain dependent. J. Bacteriol. 2007, 189, 1382–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Throup, J.P.; Koretke, K.K.; Bryant, A.P.; Ingraham, K.A.; Chalker, A.F.; Ge, Y.; Marra, A.; Wallis, N.G.; Brown, J.R.; Holmes, D.J.; et al. A genomic analysis of two-component signal transduction in Streptococcus pneumoniae. Mol. Microbiol. 2000, 35, 566–576. [Google Scholar] [CrossRef]
- Tettelin, H.; Nelson, K.E.; Paulsen, I.T.; Eisen, J.A.; Read, T.D.; Peterson, S.; Heidelberg, J.; DeBoy, R.T.; Haft, D.H.; Dodson, R.J.; et al. Complete Genome Sequence of a Virulent Isolate of Streptococcus pneumoniae. Science 2001, 293, 498. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, M.R.; Gutiérrez-Fernández, J.; Pribyl, T.; Gisch, N.; Saleh, M.; Rohde, M.; Petruschka, L.; Burchhardt, G.; Schwudke, D.; Hermoso, J.A.; et al. Structure of the pneumococcal l,d-carboxypeptidase DacB and pathophysiological effects of disabled cell wall hydrolases DacA and DacB. Mol. Microbiol. 2014, 93, 1183–1206. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.; Bartual, S.G.; Abdullah, M.R.; Jensch, I.; Asmat, T.M.; Petruschka, L.; Pribyl, T.; Gellert, M.; Lillig, C.H.; Antelmann, H.; et al. Molecular architecture of Streptococcus pneumoniae surface thioredoxin-fold lipoproteins crucial for extracellular oxidative stress resistance and maintenance of virulence. EMBO Mol. Med. 2013, 5, 1852–1870. [Google Scholar] [CrossRef]
- Krebs, H.A. Chemical Composition of Blood Plasma and Serum. Annu. Rev. Biochem. 1950, 19, 409–430. [Google Scholar] [CrossRef]
- Nanduri, B.; Shah, P.; Ramkumar, M.; Allen, E.B.; Swiatlo, E.; Burgess, S.C.; Lawrence, M.L. Quantitative analysis of Streptococcus pneumoniae TIGR4 response to in vitro iron restriction by 2-D LC ESI MS/MS. Proteomics 2008, 8, 2104–2114. [Google Scholar] [CrossRef]
- Bidossi, A.; Mulas, L.; Decorosi, F.; Colomba, L.; Ricci, S.; Pozzi, G.; Deutscher, J.; Viti, C.; Oggioni, M.R. A Functional Genomics Approach to Establish the Complement of Carbohydrate Transporters in Streptococcus pneumoniae. PLoS ONE 2012, 7, e33320. [Google Scholar] [CrossRef] [Green Version]
- Buckwalter, C.M.; King, S.J. Pneumococcal carbohydrate transport: Food for thought. Trends Microbiol. 2012, 20, 517–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trappetti, C.; Kadioglu, A.; Carter, M.; Hayre, J.; Iannelli, F.; Pozzi, G.; Andrew, P.W.; Oggioni, M.R. Sialic Acid: A Preventable Signal for Pneumococcal Biofilm Formation, Colonization, and Invasion of the Host. J. Infect. Dis. 2009, 199, 1497–1505. [Google Scholar] [CrossRef]
- Abranches, J.; Chen, Y.-Y.M.; Burne, R.A. Galactose metabolism by Streptococcus mutans. Appl. Environ. Microbiol. 2004, 70, 6047–6052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afzal, M.; Shafeeq, S.; Kuipers, O.P. LacR is a repressor of lacABCD and LacT is an activator of lacTFEG, constituting the lac gene cluster in Streptococcus pneumoniae. Appl. Environ. Microbiol. 2014, 80, 5349–5358. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, S.M.; Kloosterman, T.G.; Kuipers, O.P.; Neves, A.R. CcpA Ensures Optimal Metabolic Fitness of Streptococcus pneumoniae. PLoS ONE 2011, 6, e26707. [Google Scholar] [CrossRef] [Green Version]
- Iskandar, C.F.; Cailliez-Grimal, C.; Borges, F.; Revol-Junelles, A.-M. Review of lactose and galactose metabolism in Lactic Acid Bacteria dedicated to expert genomic annotation. Trends Food Sci. Technol. 2019, 88, 121–132. [Google Scholar] [CrossRef]
- Iyer, R.; Camilli, A. Sucrose metabolism contributes to in vivo fitness of Streptococcus pneumoniae. Mol. Microbiol. 2007, 66, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Jensch, I.; Gámez, G.; Rothe, M.; Ebert, S.; Fulde, M.; Somplatzki, D.; Bergmann, S.; Petruschka, L.; Rohde, M.; Nau, R.; et al. PavB is a surface-exposed adhesin of Streptococcus pneumoniae contributing to nasopharyngeal colonization and airways infections. Mol. Microbiol. 2010, 77, 22–43. [Google Scholar] [CrossRef] [PubMed]
- Voß, S.; Gámez, G.; Hammerschmidt, S. Impact of pneumococcal microbial surface components recognizing adhesive matrix molecules on colonization. Mol. Oral Microbiol. 2012, 27, 246–256. [Google Scholar] [CrossRef]
- Kreikemeyer, B.; Gámez, G.; Margarit, I.; Giard, J.C.; Hammerschmidt, S.; Hartke, A.; Podbielski, A. Genomic organization, structure, regulation and pathogenic role of pilus constituents in major pathogenic Streptococci and Enterococci. Int. J. Med. Microbiol. 2011, 301, 240–251. [Google Scholar] [CrossRef]
- Hilleringmann, M.; Giusti, F.; Baudner, B.C.; Masignani, V.; Covacci, A.; Rappuoli, R.; Barocchi, M.A.; Ferlenghi, I. Pneumococcal pili are composed of protofilaments exposing adhesive clusters of Rrg A. PLoS Pathog. 2008, 4, e1000026. [Google Scholar] [CrossRef]
- Ng, W.-L.; Tsui, H.-C.T.; Winkler, M.E. Regulation of the pspA Virulence Factor and Essential pcsB Murein Biosynthetic Genes by the Phosphorylated VicR (YycF) Response Regulator in Streptococcus pneumoniae. J. Bacteriol. 2005, 187, 7444. [Google Scholar] [CrossRef] [Green Version]
- Standish, A.J.; Stroeher, U.H.; Paton, J.C. The two-component signal transduction system RR06/HK06 regulates expression of cbpA in Streptococcus pneumoniae. Proc. Natl. Acad. Sci. USA 2005, 102, 7701. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Fernández, J.; Saleh, M.; Alcorlo, M.; Gómez-Mejía, A.; Pantoja-Uceda, D.; Treviño, M.A.; Voß, F.; Abdullah, M.R.; Galán-Bartual, S.; Seinen, J.; et al. Modular Architecture and Unique Teichoic Acid Recognition Features of Choline-Binding Protein L (CbpL) Contributing to Pneumococcal Pathogenesis. Sci. Rep. 2016, 6, 38094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bewley, M.A.; Naughton, M.; Preston, J.; Mitchell, A.; Holmes, A.; Marriott, H.M.; Read, R.C.; Mitchell, T.J.; Whyte, M.K.B.; Dockrell, D.H. Pneumolysin Activates Macrophage Lysosomal Membrane Permeabilization and Executes Apoptosis by Distinct Mechanisms without Membrane Pore Formation. mBio 2014, 5, e01710-14. [Google Scholar] [CrossRef] [Green Version]
- Hammerschmidt, S.; Talay, S.R.; Brandtzaeg, P.; Chhatwal, G.S. SpsA, a novel pneumococcal surface protein with specific binding to secretory Immunoglobulin A and secretory component. Mol. Microbiol. 1997, 25, 1113–1124. [Google Scholar] [CrossRef]
- Dave, S.; Brooks-Walter, A.; Pangburn, M.K.; McDaniel, L.S. PspC, a Pneumococcal Surface Protein, Binds Human Factor H. Infect. Immun. 2001, 69, 3435. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Gong, Y.; Dong, S.; Zhang, L.; Lai, X.; Zhang, X.; Yin, Y. Pneumococcal ClpP modulates the maturation and activation of human dendritic cells: Implications for pneumococcal infections. J. Leukoc. Biol. 2013, 93, 737–749. [Google Scholar] [CrossRef]
- Surewaard, B.G.J.; Trzciński, K.; Jacobino, S.R.; Hansen, I.S.; Vughs, M.M.; Sanders, E.A.M.; van der Ende, A.; van Strijp, J.A.G.; de Haas, C.J.C. Pneumococcal immune evasion: ZmpC inhibits neutrophil influx. Cel. Microbiol. 2013, 15, 1753–1765. [Google Scholar] [CrossRef]
- Beiter, K.; Wartha, F.; Albiger, B.; Normark, S.; Zychlinsky, A.; Henriques-Normark, B. An endonuclease allows Streptococcus pneumoniae to escape from neutrophil extracellular traps. Curr. Biol. 2006, 16, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Ohkuma, S.; Poole, B. Fluorescence probe measurement of the intralysosomal pH in living cells and the perturbation of pH by various agents. Proc. Natl. Acad. Sci. USA 1978, 75, 3327–3331. [Google Scholar] [CrossRef] [Green Version]
- Casiano-Colón, A.; Marquis, R.E. Role of the arginine deiminase system in protecting oral bacteria and an enzymatic basis for acid tolerance. Appl. Environ. Microbiol. 1988, 54, 1318–1324. [Google Scholar] [CrossRef] [Green Version]
- Gennaris, A.; Collet, J.-F. The ‘captain of the men of death’, Streptococcus pneumoniae, fights oxidative stress outside the ‘city wall’. EMBO Mol. Med. 2013, 5, 1798–1800. [Google Scholar] [CrossRef]
- Tseng, H.-J.; McEwan, A.G.; Paton, J.C.; Jennings, M.P. Virulence of Streptococcus pneumoniae: PsaA Mutants Are Hypersensitive to Oxidative Stress. Infect. Immun. 2002, 70, 1635. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, Y.M.; Kerr, A.R.; McCluskey, J.; Mitchell, T.J. Role of HtrA in the virulence and competence of Streptococcus pneumoniae. Infect. Immun. 2004, 72, 3584–3591. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Capsule Type | Resistance | Knockout Genes | Reference |
|---|---|---|---|---|
| TIGR4 | 4 | - | - | [27] |
| TIGR4Δrr09 | 4 | erythromycin | sp_0661 | This study |
| TIGR4Δhk09 | 4 | erythromycin | sp_0662 | This study |
| TIGR4Δtcs09 | 4 | erythromycin | sp_0661, sp_0662 | This study |
| TIGR4lux | 4 | kanamycin | - | [14] |
| TIGR4luxΔrr09 | 4 | kanamycin, erythromycin | sp_0661 | This study |
| TIGR4luxΔhk09 | 4 | kanamycin, erythromycin | sp_0662 | This study |
| TIGR4luxΔtcs09 | 4 | kanamycin, erythromycin | sp_0661, sp_0662 | This study |
| TIGR4Δcps | 4 | kanamycin | sp_0343–sp_0360 | [14] |
| TIGR4ΔcpsΔrr09 | 4 | kanamycin, erythromycin | sp_0343–sp_0360, sp_0661 | This study |
| TIGR4ΔcpsΔhk09 | 4 | kanamycin, erythromycin | sp_0343–sp_0360, sp_0662 | This study |
| TIGR4ΔcpsΔtcs09 | 4 | kanamycin, erythromycin | sp_0343–sp_0360, sp_0661, sp_0662, | This study |
| Target Gene | Primer | Sequence 5′–3′ |
|---|---|---|
| enolase (sp_1128) | enoRT_F enoRT_R | CGGACGTGGTATGGTTCCA TAGCCAATGATAGCTTCAGCA |
| rrgA (sp_0462) | RT_rrgA_F RT_rrgA_R | GTGATTAAGGAGACAGGCGA GGTGTATGTCCCAGGTTTTAT |
| rrgB (sp_0463) | rrgBF2 rrgBmut2 | TGGGACGACAACAACATC GATCAATATTCACTCCTAGAG |
| rrgC (sp_0464) | RT_rrgC_F RT_rrgC_R | CGGTTGCAAGTATGGAAGTT TACTTCAATCTGATTCTCAAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirschmann, S.; Gómez-Mejia, A.; Kohler, T.P.; Voß, F.; Rohde, M.; Brendel, M.; Hammerschmidt, S. The Two-Component System 09 of Streptococcus pneumoniae Is Important for Metabolic Fitness and Resistance during Dissemination in the Host. Microorganisms 2021, 9, 1365. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071365
Hirschmann S, Gómez-Mejia A, Kohler TP, Voß F, Rohde M, Brendel M, Hammerschmidt S. The Two-Component System 09 of Streptococcus pneumoniae Is Important for Metabolic Fitness and Resistance during Dissemination in the Host. Microorganisms. 2021; 9(7):1365. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071365
Chicago/Turabian StyleHirschmann, Stephanie, Alejandro Gómez-Mejia, Thomas P. Kohler, Franziska Voß, Manfred Rohde, Max Brendel, and Sven Hammerschmidt. 2021. "The Two-Component System 09 of Streptococcus pneumoniae Is Important for Metabolic Fitness and Resistance during Dissemination in the Host" Microorganisms 9, no. 7: 1365. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071365