
Prokaryotic Communities in the Thalassohaline Tuz Lake, Deep Zone, and Kayacik, Kaldirim and Yavsan Salterns (Turkey) Assessed by 16S rRNA Amplicon Sequencing

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
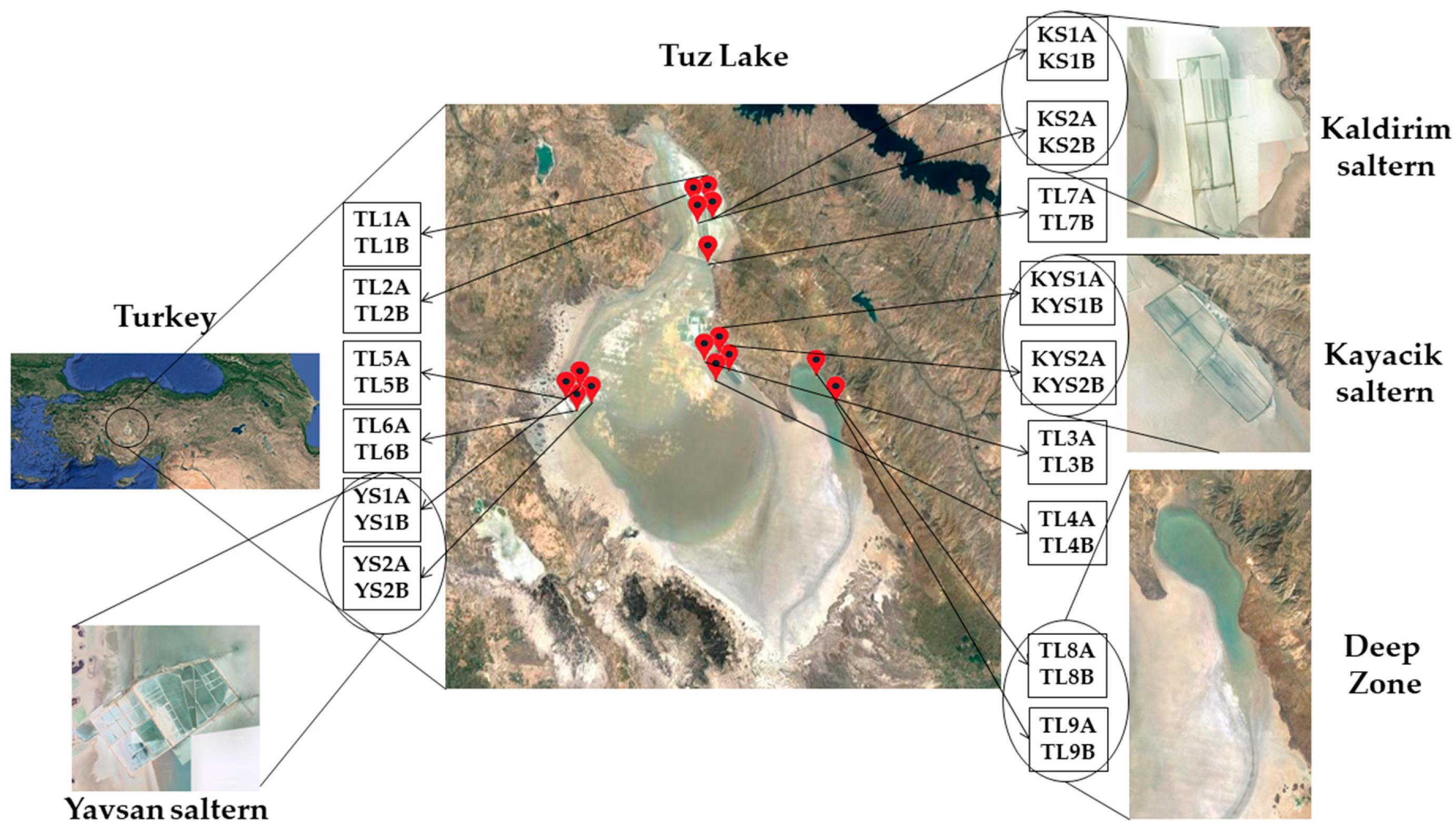
2.1. Brine Sample Collection
2.2. Physico-Chemical Analyses
2.3. DNA Extraction from the Brine Samples
2.4. 16S rRNA Amplicon Processing from the Extracted DNA Samples
2.5. Bioinformatic Analysis
2.6. Statistical Analyses
2.7. Sequencing Data
3. Results
3.1. Physico-Chemical Analyses
3.2. Alpha Diversity and Richness Metrics
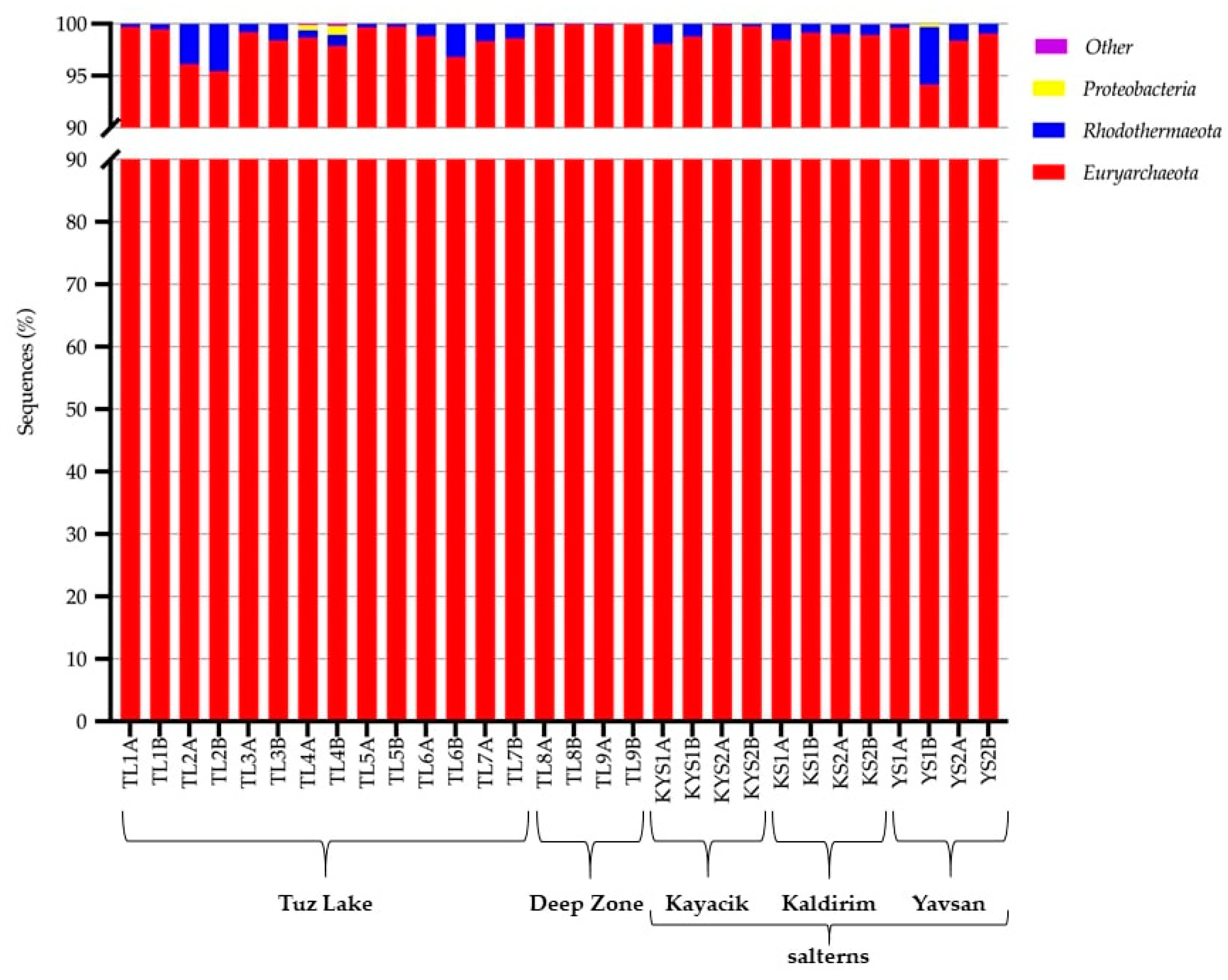
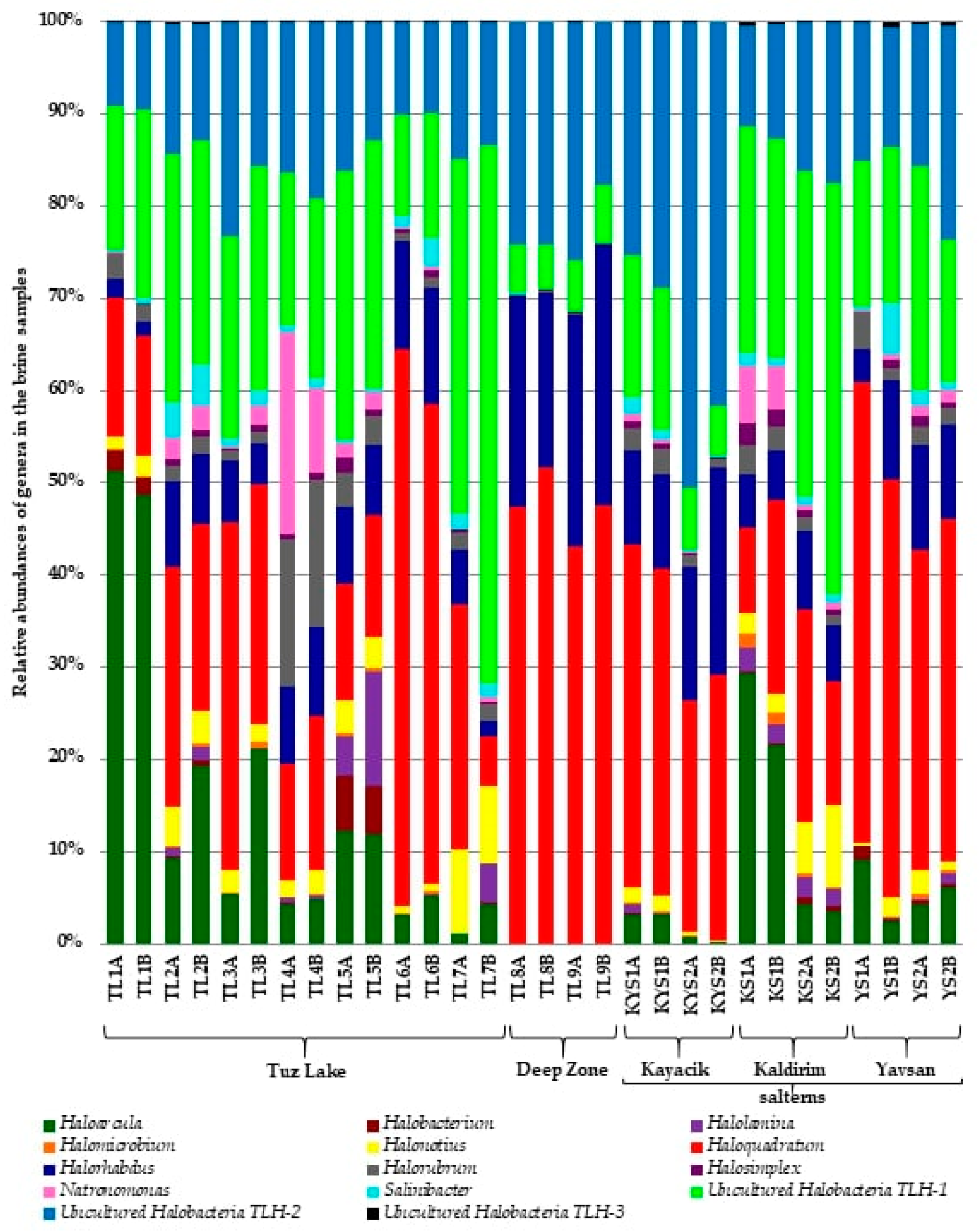
3.3. Prokaryotic Communities in Tuz Lake, Deep Zone, and Kayacik, Kaldirim, and Yavsan Salterns
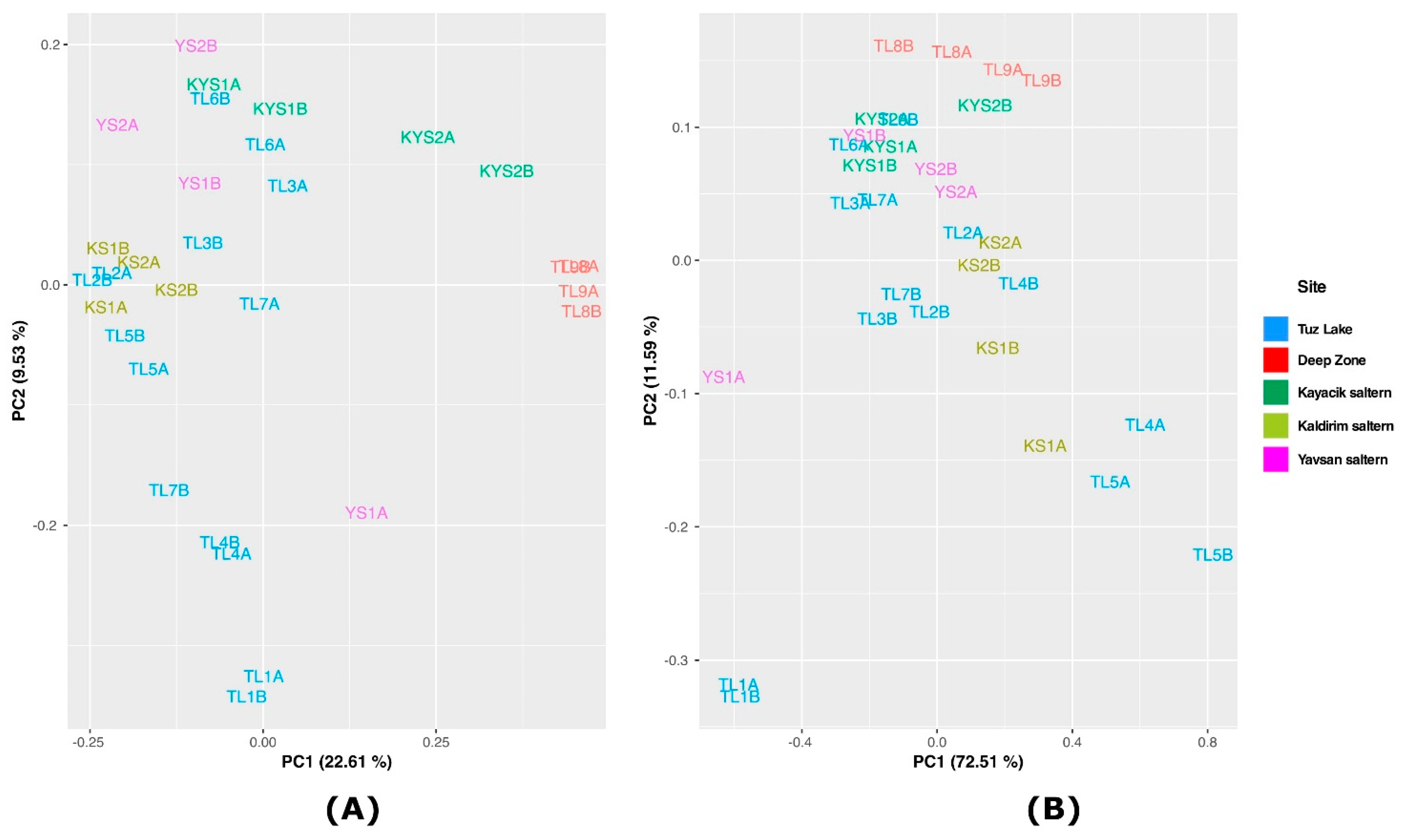
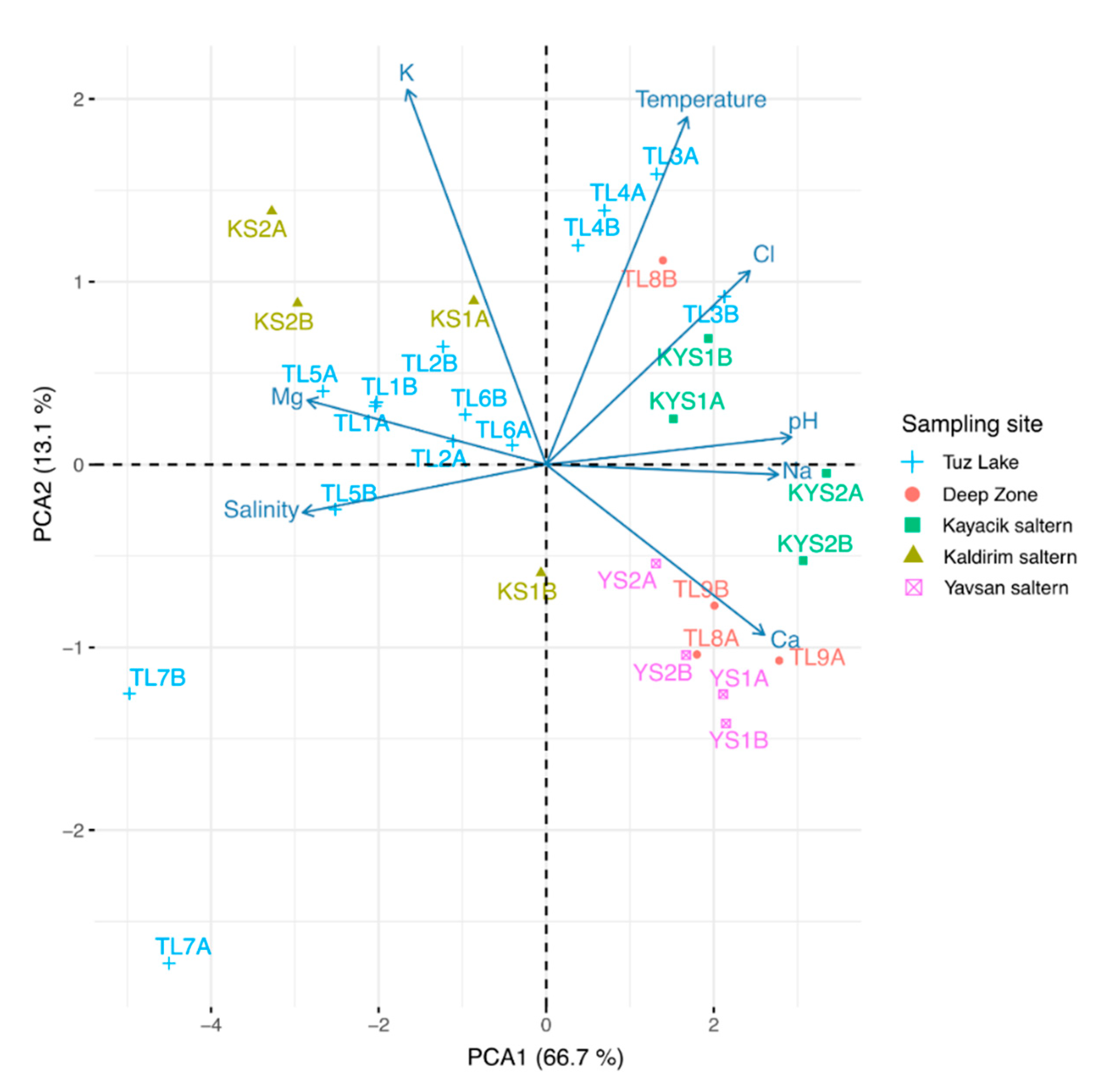
3.4. Principal Components Analysis (PCA)
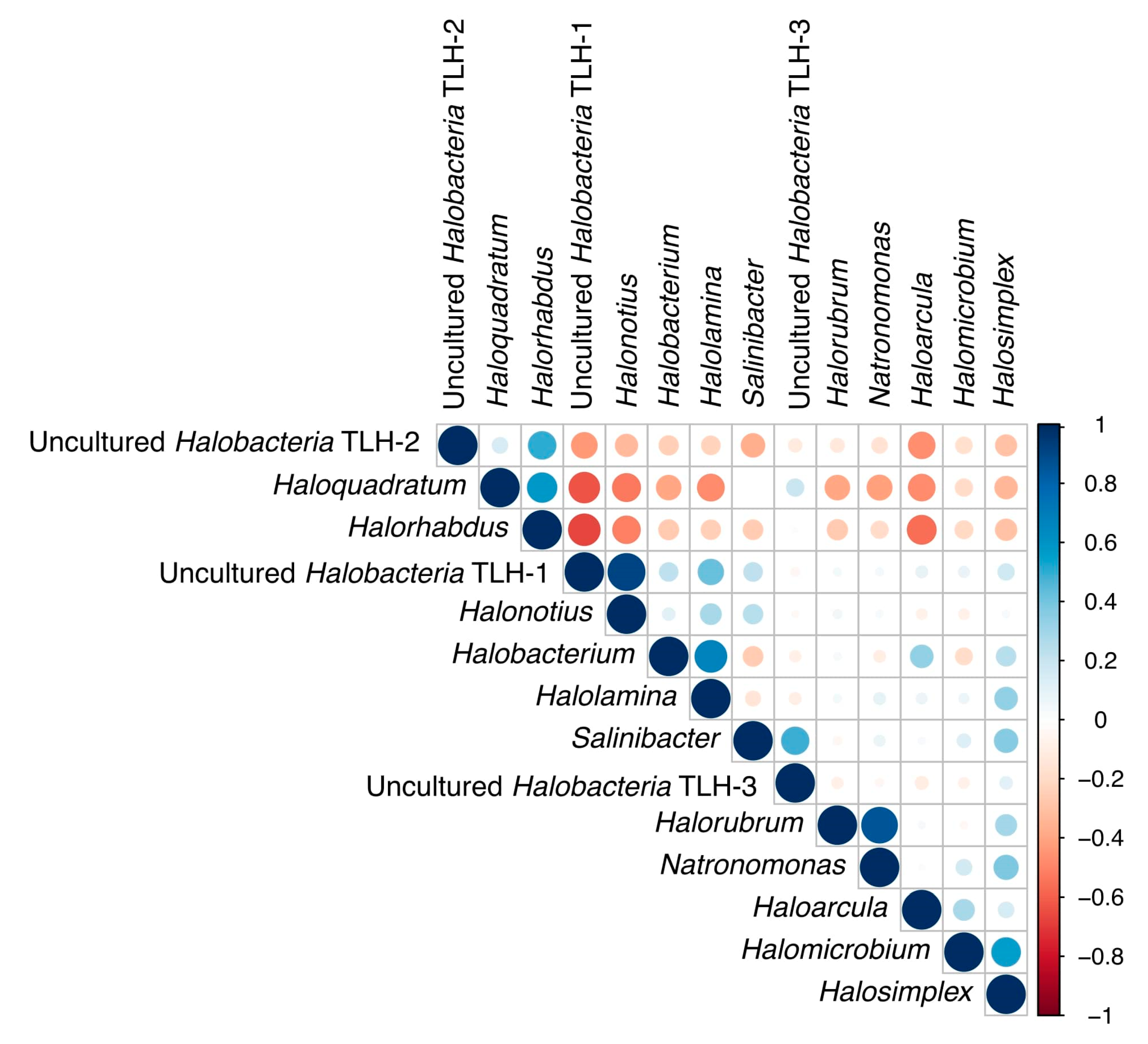
3.5. Correlations among Genera
3.6. Correlations among Physico-Chemical Parameters and Brine Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodríguez-Valera, F. Characteristics and microbial ecology of hypersaline environments. In Halophilic Bacteria; Rodríguez-Valera, F., Ed.; CRC Press: Boca Raton, FL, USA, 1988; Volume 1, pp. 3–30. [Google Scholar]
- Ventosa, A. Unusual micro-organisms from unusual habitats: Hypersaline environments. In Prokaryotic Diversity: Mechanisms and Significance; Society for General Microbiology Symposia, Logan, N.A., Lappin-Scott, H.M., Oyston, P.C.F., Eds.; Cambridge University Press: Cambridge, UK, 2006; Volume 66, pp. 223–254. [Google Scholar]
- Ventosa, A.; Fernández, A.B.; León, M.J.; Sánchez-Porro, C.; Rodríguez-Valera, F. The Santa Pola saltern as a model for studying the microbiota of hypersaline environments. Extremophiles 2014, 18, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Ventosa, A.; de la Haba, R.R.; Sánchez-Porro, C.; Papke, R.T. Microbial diversity of hypersaline environments: A metagenomic approach. Curr. Opin. Microbiol. 2015, 25, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Amoozegar, M.A.; Siroosi, M.; Atashgahi, S.; Smidt, H.; Ventosa, A. Systematics of haloarchaea and biotechnological potential of their hydrolytic enzymes. Microbiology 2017, 163, 623–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vera-Gargallo, B.; Ventosa, A. Metagenomic insights into the phylogenetic and metabolic diversity of the prokaryotic community dwelling in hypersaline soils from the odiel saltmarshes (SW Spain). Genes 2018, 9, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vera-Gargallo, B.; Chowdhury, T.R.; Brown, J.; Fansler, S.J.; Durán-Viseras, A.; Sánchez-Porro, C.; Bailey, V.L.; Jansson, J.K.; Ventosa, A. Spatial distribution of prokaryotic communities in hypersaline soils. Sci. Rep. 2019, 9, 1769. [Google Scholar] [CrossRef]
- La Cono, V.; Bortoluzzi, G.; Messina, E.; La Spada, G.; Smedile, F.; Giuliano, L.; Borghini, M.; Stumpp, C.; Schmitt-Kopplin, P.; Harir, M.; et al. The discovery of Lake Hephaestus, the youngest athalassohaline deep-sea formation on Earth. Sci. Rep. 2019, 9, 8031. [Google Scholar] [CrossRef] [Green Version]
- Javor, B.J. Hypersaline Environments: Microbiology and Biogeochemistry; Springer: Berlin, Germany, 1989. [Google Scholar]
- Grant, W.D. General view of halophiles. In Superbugs. Microorganisms in Extreme Environments; Horikoshi, K., Grant, W.D., Eds.; Springer: Tokyo, Japan, 1990; pp. 15–37. [Google Scholar]
- Oren, A. Halophilic Microorganisms and Their Environments; Kluwer Scientific Publishers: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Pagaling, E.; Wang, H.; Venables, M.; Wallace, A.; Grant, W.D.; Cowan, D.A.; Jones, B.E.; Ma, Y.; Ventosa, A.; Heaphy, S. Microbial biogeography of six salt lakes in Inner Mongolia, China, and a salt lake in Argentina. Appl. Environ. Microbiol. 2009, 75, 5750–5760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podell, S.; Ugalde, J.A.; Narasingarao, P.; Banfield, J.F.; Heidelberg, K.B.; Allen, E.E. Assembly-driven community genomics of a hypersaline microbial ecosystem. PLoS ONE 2013, 8, e61692. [Google Scholar] [CrossRef] [Green Version]
- Cavicchioli, R. Microbial ecology of Antarctic aquatic systems. Nat. Rev. Microbiol. 2015, 13, 691–706. [Google Scholar] [CrossRef]
- Oren, A. Life at high salt concentrations. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 421–440. [Google Scholar]
- Amoozegar, M.A.; Safarpour, A.; Noghabi, K.A.; Bakhtiary, T.; Ventosa, A. Halophiles and their vast potential in biofuel production. Front. Microbiol. 2019, 10, 1895. [Google Scholar] [CrossRef]
- Corral, P.; Amoozegar, M.A.; Ventosa, A. Halophiles and their biomolecules: Recent advances and future applications in biomedicine. Mar. Drugs 2020, 18, 33. [Google Scholar] [CrossRef] [Green Version]
- Çınar, S.; Mutlu, M.B. Prokaryotic community compositions of the hypersaline sediments of Tuz Lake demonstrated by cloning and high-throughput sequencing. Microbiology 2020, 89, 756–768. [Google Scholar] [CrossRef]
- Tuz Lake Special Environmental Protection Area Management Plan (2014–2018); Directorate General for Preservation of Natural Heritage; Tuz Lake Special Environmental Protection Area Management Plan, 2014–2018.
- Erol, O. Tuz Gölü Havzasının jeolojisi ve jeomorfolojisi. Mineral Research and Exploration Institute of Turkey (MTA) Report No. 4220. 1969. (In Turkish) [Google Scholar]
- Kilic, O.; Kilic, A.M. Salt crust mineralogy and geochemical evolution of the Salt Lake (Tuz Gölü), Turkey. Sci. Res. Essays 2010, 5, 1317–1324. [Google Scholar]
- Birbir, M.; Calli, B.; Mertoglu, B.; Bardavid, R.E.; Oren, A.; Ogmen, M.N.; Ogan, A. Extremely halophilic Archaea from Tuz Lake, Turkey, and the adjacent Kaldirim and Kayacik salterns. World J. Microbiol. Biotechnol. 2007, 23, 309–316. [Google Scholar] [CrossRef]
- Karakaya, M.Ç.; Bozdağ, A.; Ercan, H.Ü.; Karakaya, N.; Delikan, A. Origin of Miocene halite from Tuz Gölü basin in Central Anatolia, Turkey: Evidences from the pure halite and fluid inclusion geochemistry. J. Geochem. Explor. 2019, 202, 1–12. [Google Scholar] [CrossRef]
- TVKGM. Official Website of Directorate General for Preservation of Natural Heritage. Available online: https://tvk.csb.gov.tr/ (accessed on 15 February 2021).
- Aksoz, N.; Kolonkaya, N. Ilımlı tuzcul karakterli bir bakterinin Tuz Gölünden (Türkiye) izolasyonu. Mikrobiyol. Bull. 1984, 18, 107–113. [Google Scholar]
- Birbir, M.; Kalli, N.; Johannson, C. Examination of salt quality of Sereflíkoçhísar Lake used in the Turkish leather industry. J. Soc. Leath. Tech. Ch. 2002, 86, 112–117. [Google Scholar]
- Birbir, M.; Sesal, C. Extremely halophilic bacterial communities in Şereflikoçhisar Salt Lake in Turkey. Turk. J. Biol. 2003, 27, 7–22. [Google Scholar]
- Ozcan, B.; Ozcengiz, G.; Coleri, A.; Cokmus, C. Diversity of halophilic archaea from six hypersaline environments in Turkey. J. Microbiol. Biotechnol. 2007, 17, 985–992. [Google Scholar]
- Mutlu, M.B.; Martínez-García, M.; Santos, F.; Peña, A.; Guven, K.; Antón, J. Prokaryotic diversity in Tuz Lake, a hypersaline environment in Inland Turkey. FEMS Microbiol. Ecol. 2008, 65, 474–483. [Google Scholar] [CrossRef]
- Ünal, H.; Kücükyildirim, S. Metabarkodlama Metabarkodlama yaklaşımıyla Tuz Gölü, Türkiye mikroorganizmalarının belirlenmesi için bir pilot çalışma. EJOSAT 2020, 366–374. Available online: https://dergipark.org.tr/en/download/article-file/1153358 (accessed on 13 July 2021).
- Oyewusi, H.A.; Abdul Wahab, R.; Edbeib, M.F.; Mohamad, M.A.N.; Abdul Hamid, A.A.; Kaya, Y.; Huyop, F. Functional profiling of bacterial communities in Lake Tuz using 16S rRNA gene sequences. Biotechnol. Biotechnol. Equip. 2021, 35, 1–10. [Google Scholar] [CrossRef]
- Çınar, S.; Mutlu, M.B. Assessing the diversity of prokaryotic communities and nanohaloarchaeal lineages in various aquatic hypersaline habitats (Turkey) using high-throughput sequencing and cloning. Eskişehir Teknik Üniversitesi Bilim Teknoloji Dergisi-C Yaşam Bilimleri Biyoteknoloji 2021, 10, 57–68. [Google Scholar] [CrossRef]
- Clarke, F.E. Determination of chloride in water improved colorimetric and titrimetric methods. Anal. Chem. 1950, 22, 553–555. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Yesil, H.; Tugtas, A.E. Removal of heavy metals from leaching effluents of sewage sludge via supported liquid membranes. Sci. Total Environ. 2019, 693, 133608. [Google Scholar] [CrossRef] [PubMed]
- Ghai, R.; Pašić, L.; Fernández, A.B.; Martin-Cuadrado, A.B.; Mizuno, C.M.; McMahon, K.D.; Papke, R.T.; Stepanauskas, R.; Rodriguez-Brito, B.; Rohwer, F.; et al. New abundant microbial groups in aquatic hypersaline environments. Sci. Rep. 2011, 1, 135. [Google Scholar] [CrossRef]
- Fernández, A.B.; Ghai, R.; Martin-Cuadrado, A.B.; Sánchez-Porro, C.; Rodríguez-Valera, F.; Ventosa, A. Prokaryotic taxonomic and metabolic diversity of an intermediate salinity hypersaline habitat assessed by metagenomics. FEMS Microbiol. Ecol. 2014, 88, 623–635. [Google Scholar] [CrossRef] [Green Version]
- Vavourakis, C.D.; Ghai, R.; Rodríguez-Valera, F.; Sorokin, D.Y.; Tringe, S.G.; Hugenholtz, P.; Muyzer, G. Metagenomic insights into the uncultured diversity and physiology of microbes in four hypersaline soda lake brines. Front. Microbiol. 2016, 7, 211. [Google Scholar] [CrossRef] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Tremblay, J.; Singh, K.; Fern, A.; Kirton, E.S.; He, S.; Woyke, T.; Lee, J.; Chen, F.; Dangl, J.L.; Tringe, S.G. Primer and platform effects on 16S rRNA tag sequencing. Front. Microbiol. 2015, 6, 771. [Google Scholar] [CrossRef] [Green Version]
- Babraham Bioinformatics FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 15 February 2021).
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [Green Version]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S-Plus, 1994th ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roldán Ahumada, J.A.; Avendaño Garrido, M.L. A commentary on diversity measures UniFrac in very small sample size. Evol. Bioinform. 2019, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Knecht, V.R.; McGinniss, J.E.; Shankar, H.M.; Clarke, E.L.; Kelly, B.J.; Imai, I.; Fitzgerald, A.S.; Bittinger, K.; Bushman, F.D.; Collman, R.G. Molecular analysis of bacterial contamination on stethoscopes in an intensive care unit. Infect. Control Hosp. Epidemiol. 2019, 40, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Grant, W.D. Life at low water activity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 1249–1267. [Google Scholar] [CrossRef] [Green Version]
- Ventosa, A.; Arahal, D.R. Physico-chemical characteristics of hypersaline environments and their biodiversity. Extremophiles 2009, 2, 247–262. [Google Scholar]
- Fernández, A.B.; Ghai, R.; Martin-Cuadrado, A.B.; Sánchez-Porro, C.; Rodríguez-Valera, F.; Ventosa, A. Metagenome sequencing of prokaryotic microbiota from two hypersaline ponds of a marine saltern in Santa Pola, Spain. Genome Announc. 2013, 1, e00933-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Meglio, L.; Santos, F.; Gomariz, M.; Almansa, C.; López, C.; Antón, J.; Nercessian, D. Seasonal dynamics of extremely halophilic microbial communities in three Argentinian salterns. FEMS Microbiol. Ecol. 2016, 92, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Baxter, B.K.; Litchfield, C.D.; Sowers, K.; Griffith, J.D.; Dassarma, P.A.; Dassarma, S. Microbial Diversity of Great Salt Lake. In Adaptation to Life at High Salt Concentrations in Archaea, Bacteria, and Eukarya; Gunde-Cimerman, N., Oren, A., Plemenitaš, A., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 9, pp. 9–25. [Google Scholar]
- Almeida-Dalmet, S.; Sikaroodi, M.; Gillevet, P.M.; Litchfield, C.D.; Baxter, B.K. Temporal study of the microbial diversity of the North Arm of Great Salt Lake, Utah, U.S. Microorganisms 2015, 3, 310–326. [Google Scholar] [CrossRef] [Green Version]
- Baxter, B.K.; Zalar, P. The extremophiles of Great Salt Lake: Complex microbiology in a dynamic hypersaline ecosystem. In Model Ecosystems in Extreme Environments; Elsevier: Amsterdam, The Netherlands, 2019; pp. 57–99. [Google Scholar]
- Narasingarao, P.; Podell, S.; Ugalde, J.A.; Brochier-Armanet, C.; Emerson, J.B.; Brocks, J.J.; Heidelberg, K.B.; Banfield, J.F.; Allen, E.E. De novo metagenomic assembly reveals abundant novel major lineage of Archaea in hypersaline microbial communities. ISME J. 2012, 6, 81–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podell, S.; Emerson, J.B.; Jones, C.M.; Ugalde, J.A.; Welch, S.; Heidelberg, K.B.; Banfield, J.F.; Allen, E.E. Seasonal fluctuations in ionic concentrations drive microbial succession in a hypersaline lake community. ISME J. 2014, 8, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Makhdoumi-Kakhki, A.; Amoozegar, M.A.; Kazemi, B.; Pašić, L.; Ventosa, A. Prokaryotic diversity in Aran-Bidgol salt lake, the largest hypersaline playa in Iran. Microbes Environ. 2012, 27, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naghoni, A.; Emtiazi, G.; Amoozegar, M.A.; Cretoiu, M.S.; Stal, L.J.; Etemadifar, Z.; Shahzadeh Fazeli, S.A.; Bolhuis, H. Microbial diversity in the hypersaline Lake Meyghan, Iran. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, A.B.; Vera-Gargallo, B.; Sánchez-Porro, C.; Ghai, R.; Papke, R.T.; Rodríguez-Valera, F.; Ventosa, A. Comparison of prokaryotic community structure from Mediterranean and Atlantic saltern concentrator ponds by a metagenomic approach. Front. Microbiol. 2014, 5, 1–12. [Google Scholar] [CrossRef]
- Fernández, A.B.; León, M.J.; Vera, B.; Sánchez-Porro, C.; Ventosa, A. Metagenomic sequence of prokaryotic microbiota from an intermediate-salinity pond of a saltern in Isla Cristina, Spain. Genome Announc. 2014, 2, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Pasić, L.; Rodríguez-Mueller, B.; Martin-Cuadrado, A.B.; Mira, A.; Rohwer, F.; Rodríguez-Valera, F. Metagenomic islands of hyperhalophiles: The case of Salinibacter ruber. BMC Genom. 2009, 10, 570. [Google Scholar] [CrossRef] [Green Version]
- Antón, J.; Llobet-Brossa, E.; Rodríguez-Valera, F.; Amann, R. Fluorescence in situ hybridization analysis of the prokaryotic community inhabiting crystallizer ponds. Environ. Microbiol. 1999, 1, 517–523. [Google Scholar] [CrossRef]
- Benlloch, S.; López-López, A.; Casamayor, E.O.; Øvreås, L.; Goddard, V.; Daae, F.L.; Smerdon, G.; Massana, R.; Joint, I.; Thingstad, F.; et al. Prokaryotic genetic diversity throughout the salinity gradient of a coastal solar saltern. Environ. Microbiol. 2002, 4, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Legault, B.A.; Lopez-Lopez, A.; Alba-Casado, J.C.; Doolittle, W.F.; Bolhuis, H.; Rodríguez-Valera, F.; Papke, R.T. Environmental genomics of “Haloquadratum walsbyi” in a saltern crystallizer indicates a large pool of accessory genes in an otherwise coherent species. BMC Genom. 2006, 7, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antón, J.; Peña, A.; Santos, F.; Martínez-García, M.; Schmitt-Kopplin, P.; Rosselló-Mora, R. Distribution, abundance and diversity of the extremely halophilic bacterium Salinibacter ruber. Saline Syst. 2008, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Galinski, E.A.; Grant, W.D.; Oren, A.; Ventosa, A. Halophiles 2010: Life in saline environments. Appl. Environ. Microbiol. 2010, 76, 6971–6981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zenke, R.; von Gronau, S.; Bolhuis, H.; Gruska, M.; Pfeiffer, F.; Oesterhelt, D. Fluorescence microscopy visualization of halomucin, a secreted 927 kDa protein surrounding Haloquadratum walsbyi cells. Front. Microbiol. 2015, 6, 249. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Ventosa, A.; Kamekura, M. Class Halobacteria. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; Whiley & Sons: Hoboken, NJ, USA, 2017; pp. 1–5. [Google Scholar]
- Gaba, S.; Kumari, A.; Medema, M.; Kaushik, R. Pan-genome analysis and ancestral state reconstruction of class halobacteria: Probability of a new super-order. Sci. Rep. 2020, 10, 21205. [Google Scholar] [CrossRef] [PubMed]
- Bolhuis, H.; Martín-Cuadrado, A.B.; Rosselli, R.; Pašić, L.; Rodríguez-Valera, F. Transcriptome analysis of Haloquadratum walsbyi: Vanity is but the surface. BMC Genom. 2017, 18, 510. [Google Scholar] [CrossRef]
- Bolhuis, H.; Poele, E.M.T.; Rodríguez-Valera, F. Isolation and cultivation of Walsby’s square archaeon. Environ. Microbiol. 2004, 6, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- Bolhuis, H.; Palm, P.; Wende, A.; Falb, M.; Rampp, M.; Rodríguez-Valera, F.; Pfeiffer, F.; Oesterhelt, D. The genome of the square archaeon Haloquadratum walsbyi: Life at the limits of water activity. BMC Genom. 2006, 7, 169. [Google Scholar] [CrossRef] [Green Version]
- Uygun, A.; Şen, E. The Salt Lake basin and natural resources I: Geochemistry of the brine of the Salt Lake (Central Anatolia-Turkey). Bull. Geol. Soc. Turk. 1978, 21, 113–120. [Google Scholar]
- Chimileski, S.; Dolas, K.; Naor, A.; Gophna, U.; Papke, R.T. Extracellular DNA metabolism in Haloferax volcanii. Front. Microbiol. 2014, 5, 57. [Google Scholar] [CrossRef]
- Hua, Z.; Ouellette, M.; Makkay, A.M.; Papke, R.T.; Zhaxybayeva, O. Nutrient supplementation experiments with saltern microbial communities implicate utilization of DNA as a source of phosphorus. ISME J. 2021, 1–12. Available online: https://0-www-nature-com.brum.beds.ac.uk/articles/s41396-021-00960-8 (accessed on 13 July 2021).
- Rodríguez-Valera, F.; Juez, G.; Kushner, D.J. Halocins: Salt-dependent bacteriocins produced by extremely halophilic rods. Can. J. Microbiol. 1982, 28, 151–154. [Google Scholar] [CrossRef]
- Torreblanca, M.; Meseguer, I.; Ventosa, A. Production of halocin is a practically universal feature of archaeal halophilic rods. Lett. Appl. Microbiol. 1994, 19, 201–205. [Google Scholar] [CrossRef]
- Birbir, M.; Eryilmaz, S.; Ogan, A. Prevention of halophilic microbial damage on brine cured hides by extremely halophilic halocin producer strains. J. Soc. Leath. Tech. Chem. 2004, 88, 99–104. [Google Scholar]
- Birbir, M.; Eryilmaz, S. Inhibiting lipolytic haloarchaeal damage on brine cured hides with halocin producer strains. J. Soc. Leath. Tech. Chem. 2007, 91, 69–72. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | PD Whole Tree | Chao 1 | Observed OTUs | Shannon | Simpson | Dominance |
|---|---|---|---|---|---|---|
| Tuz Lake | ||||||
| TL1A | 39.02 | 2879.40 | 1517.00 | 6.22 | 0.94 | 0.06 |
| TL1B | 41.00 | 3388.46 | 1753.00 | 6.09 | 0.94 | 0.06 |
| TL2A | 77.94 | 8354.98 | 3799.00 | 7.50 | 0.95 | 0.05 |
| TL2B | 88.63 | 9402.42 | 4539.00 | 7.82 | 0.96 | 0.04 |
| TL3A | 56.38 | 5378.22 | 2756.00 | 6.31 | 0.90 | 0.10 |
| TL3B | 60.63 | 5983.23 | 2881.00 | 7.23 | 0.95 | 0.05 |
| TL4A | 53.08 | 3761.75 | 1936.00 | 6.75 | 0.95 | 0.05 |
| TL4B | 59.47 | 3958.28 | 2357.00 | 7.23 | 0.96 | 0.04 |
| TL5A | 64.97 | 6757.91 | 3134.00 | 8.11 | 0.98 | 0.02 |
| TL5B | 76.81 | 7325.52 | 3930.00 | 8.46 | 0.98 | 0.02 |
| TL6A | 57.09 | 5566.74 | 2674.00 | 5.38 | 0.77 | 0.23 |
| TL6B | 69.76 | 7010.20 | 3450.00 | 6.15 | 0.84 | 0.16 |
| TL7A | 51.98 | 4779.82 | 2134.00 | 5.73 | 0.89 | 0.11 |
| TL7B | 53.02 | 4555.30 | 2241.00 | 5.24 | 0.80 | 0.20 |
| Deep Zone | ||||||
| TL8A | 24.94 | 1805.01 | 914.00 | 4.43 | 0.82 | 0.18 |
| TL8B | 20.61 | 1480.57 | 707.00 | 4.29 | 0.79 | 0.21 |
| TL9A | 23.76 | 1684.02 | 883.00 | 4.65 | 0.85 | 0.15 |
| TL9B | 26.09 | 2007.02 | 1008.00 | 4.55 | 0.83 | 0.17 |
| Kayacik saltern | ||||||
| KYS1A | 71.63 | 8049.00 | 3488.00 | 6.98 | 0.91 | 0.09 |
| KYS1B | 58.10 | 6263.30 | 2768.00 | 6.94 | 0.91 | 0.09 |
| KYS2A | 40.67 | 3763.57 | 1812.00 | 5.52 | 0.90 | 0.10 |
| KYS2B | 34.26 | 2669.73 | 1453.00 | 5.31 | 0.90 | 0.10 |
| Kaldirim saltern | ||||||
| KS1A | 78.75 | 8057.13 | 4026.00 | 8.27 | 0.98 | 0.02 |
| KS1B | 81.74 | 8825.45 | 4242.00 | 8.19 | 0.97 | 0.03 |
| KS2A | 78.01 | 8276.09 | 3692.00 | 7.24 | 0.94 | 0.06 |
| KS2B | 67.12 | 6241.22 | 2900.00 | 6.66 | 0.91 | 0.09 |
| Yavsan saltern | ||||||
| YS1A | 32.29 | 2572.08 | 1321.00 | 5.16 | 0.82 | 0.18 |
| YS1B | 70.16 | 6340.94 | 3174.00 | 5.98 | 0.85 | 0.15 |
| YS2A | 104.00 | 10,259.51 | 5725.00 | 7.17 | 0.92 | 0.08 |
| YS2B | 85.79 | 8352.18 | 4468.00 | 7.28 | 0.92 | 0.08 |
| Genus/Taxon | Phylum | Tuz Lake | Deep Zone | Kayacik Saltern | Kaldirim Saltern | Yavsan Saltern |
|---|---|---|---|---|---|---|
| Uncultured Halobacteria TLH-1 | Euryarchaeota | 24.61 ± 3.20 | 5.43 ± 0.28 | 10.65 ± 2.66 | 31.77 ± 4.86 | 17.93 ± 2.12 |
| Haloquadratum | Euryarchaeota | 23.99 ± 4.25 | 47.32 ± 1.74 | 31.33 ± 2.78 | 16.57 ± 3.18 | 41.30 ± 3.60 |
| Haloarcula | Euryarchaeota | 14.36 ± 4.32 | 0.03 ± 0.00 | 1.80 ± 0.79 | 14.52 ± 6.39 | 5.46 ± 1.42 |
| Uncultured Halobacteria TLH-2 | Euryarchaeota | 13.98 ± 1.04 | 22.99 ± 1.78 | 36.18 ± 5.85 | 14.14 ± 1.51 | 16.40 ± 2.16 |
| Halorhabdus | Euryarchaeota | 6.90 ± 0.93 | 23.76 ± 1.95 | 14.22 ± 2.93 | 6.40 ± 0.69 | 8.82 ± 1.78 |
| Halorubrum | Euryarchaeota | 3.87 ± 1.34 | 0.15 ± 0.02 | 1.80 ± 0.42 | 2.07 ± 0.46 | 2.26 ± 0.60 |
| Halonotius | Euryarchaeota | 3.25 ± 0.66 | 0.02 ± 0.00 | 0.96 ± 0.41 | 4.69 ± 1.65 | 1.50 ± 0.51 |
| Natronomonas | Euryarchaeota | 3.00 ± 1.55 | 0.02 ± 0.00 | 0.30 ± 0.15 | 2.99 ± 1.39 | 0.81 ± 0.24 |
| Halolamina | Euryarchaeota | 1.73 ± 0.91 | <0.01 | 0.27 ± 0.23 | 2.15 ± 0.17 | 0.39 ± 0.24 |
| Salinibacter | Rhodothermaeota | 1.54 ± 0.37 | 0.09 ± 0.03 | 0.88 ± 0.42 | 1.09 ± 0.16 | 2.10 ± 1.13 |
| Halobacterium | Euryarchaeota | 1.19 ± 0.53 | 0.01 ± 0.00 | 0.04 ± 0.02 | 0.44 ± 0.19 | 0.51 ± 0.27 |
| Halosimplex | Euryarchaeota | 0.58 ± 0.11 | 0.08 ± 0.01 | 0.37 ± 0.14 | 1.34 ± 0.43 | 0.67 ± 0.23 |
| Halomicrobium | Euryarchaeota | 0.23 ± 0.05 | 0.01 ± 0.00 | 0.17 ± 0.05 | 0.84 ± 0.36 | 0.30 ± 0.14 |
| Haloplanus | Euryarchaeota | 0.16 ± 0.04 | 0.01 ± 0.00 | 0.46 ± 0.11 | 0.42 ± 0.09 | 0.60 ± 0.11 |
| Uncultured Halobacteria TLH-3 | Euryarchaeota | 0.07 ± 0.02 | 0.00 ± 0.00 | 0.06 ± 0.03 | 0.22 ± 0.05 | 0.34 ± 0.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akpolat, C.; Fernández, A.B.; Caglayan, P.; Calli, B.; Birbir, M.; Ventosa, A. Prokaryotic Communities in the Thalassohaline Tuz Lake, Deep Zone, and Kayacik, Kaldirim and Yavsan Salterns (Turkey) Assessed by 16S rRNA Amplicon Sequencing. Microorganisms 2021, 9, 1525. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071525
Akpolat C, Fernández AB, Caglayan P, Calli B, Birbir M, Ventosa A. Prokaryotic Communities in the Thalassohaline Tuz Lake, Deep Zone, and Kayacik, Kaldirim and Yavsan Salterns (Turkey) Assessed by 16S rRNA Amplicon Sequencing. Microorganisms. 2021; 9(7):1525. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071525
Chicago/Turabian StyleAkpolat, Can, Ana Beatriz Fernández, Pinar Caglayan, Baris Calli, Meral Birbir, and Antonio Ventosa. 2021. "Prokaryotic Communities in the Thalassohaline Tuz Lake, Deep Zone, and Kayacik, Kaldirim and Yavsan Salterns (Turkey) Assessed by 16S rRNA Amplicon Sequencing" Microorganisms 9, no. 7: 1525. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071525