
Invasive Species as Hosts of Zoonotic Infections: The Case of American Mink (Neovison vison) and Leishmania infantum


Abstract
:1. Introduction
2. Materials and Methods
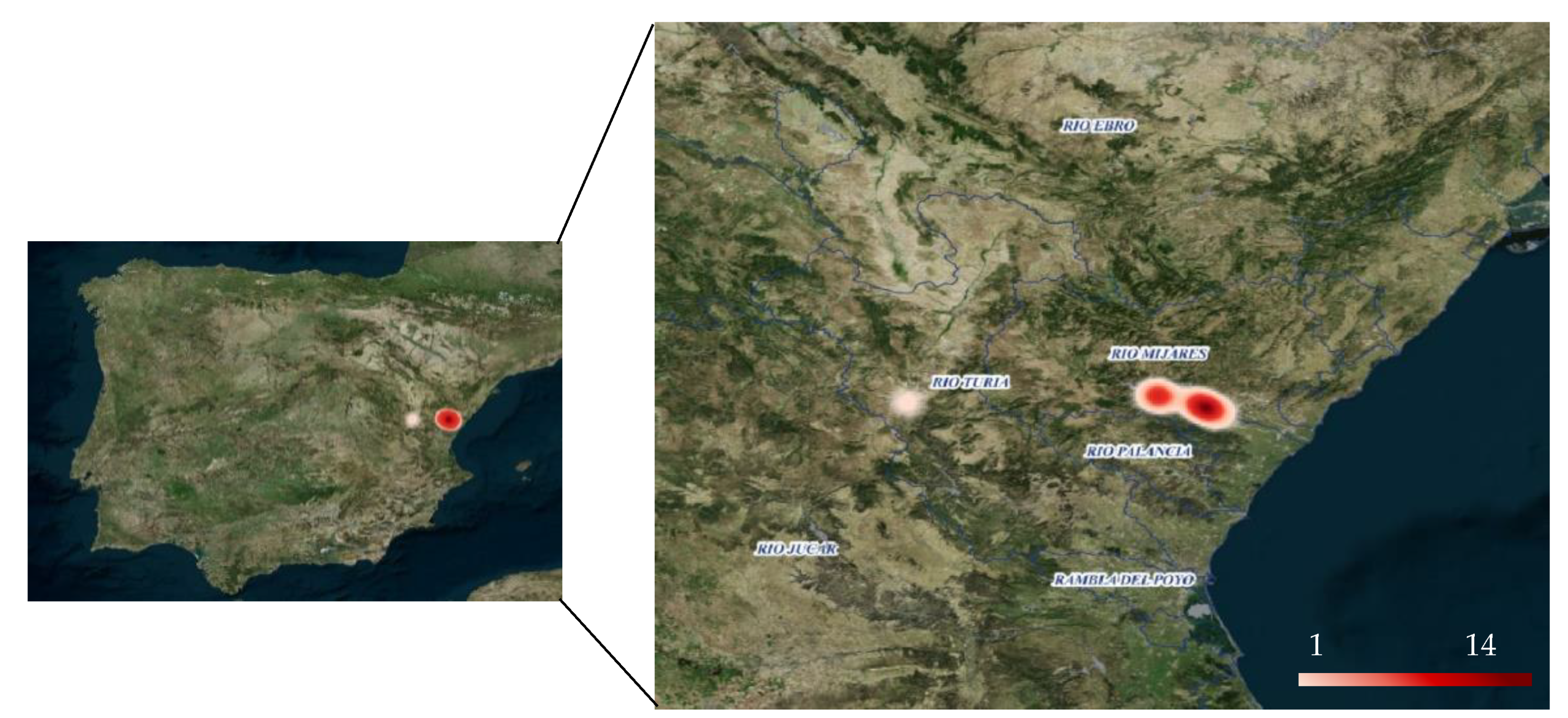
2.1. Samples and Area of Study
2.2. Culture of Leishmania and Determination of the Detection Limit
2.3. DNA Isolation of Spleen Samples and PCR
2.4. Sequencing
3. Results
3.1. Samples
3.2. PCR Detection Limit
3.2.1. PCR Detection Limit Using a Reference Strain
3.2.2. PCR Detection Limit Using DNA from the Bone Marrow of an Infected Dog
3.3. PCR Results from Mink Spleen Samples
3.4. Sequences Obtained
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruíz-Olmo, J.; Palazón, S.; Bueno, F.; Bravo, C.; Munilla, I.; Romero, R. Distribution, status and colonization of the American mink (Mustela vison) in Spain. J. Wildl. Res. 1997, 2, 30–36. [Google Scholar]
- Sidorovich, V.; Kruuk, H.; Macdonald, D.W. Body size, and interactions between European and American mink (Mustela lutreola and M. vison) in Eastern Europe. J. Zool. 1999, 248, 521–527. [Google Scholar] [CrossRef]
- Bonesi, L.; Palazón, S. The American mink in Europe: Status, impacts, and control. Biol. Conserv. 2007, 134, 470–483. [Google Scholar] [CrossRef]
- Martínez-Rondán, F.J.; Ruiz de Ybáñez, M.R.; Tizzani, P.; López-Beceiro, A.M.; Fidalgo, L.E.; Martínez-Carrasco, C. The American mink (Neovison vison) is a competent host for native European parasites. Vet. Parasitol. 2017, 247, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Ribas, M.P.; Almería, S.; Fernández-Aguilar, X.; De Pedro, G.; Lizarraga, P.; Alarcia-Alejos, O.; Molina-López, R.; Obón, E.; Gholipour, H.; Temiño, C.; et al. Tracking Toxoplasma gondii in freshwater ecosystems: Interaction with the invasive American mink (Neovison vison) in Spain. Parasitol. Res. 2018, 117, 2275–2281. [Google Scholar] [CrossRef] [Green Version]
- Gholipour, H.; Busquets, N.; Fernández-Aguilar, X.; Sánchez, A.; Ribas, M.P.; De Pedro, G.; Lizarraga, P.; Alarcia-Alejos, O.; Temiño, C.; Cabezón, O. Influenza A Virus surveillance in the invasive American Mink (Neovison vison) from freshwater ecosystems, Northern Spain. Zoonoses Public Health 2017, 64, 363–369. [Google Scholar] [CrossRef]
- Mahdy, M.A.A.; Younis, W.; Ewaida, Z. An overview of SARS-CoV-2 and animal infection. Front. Vet. Sci. 2020, 7, 596391. [Google Scholar] [CrossRef]
- Molina, R.; Jiménez, M.; García-Martínez, J.; San Martín, J.V.; Carrillo, E.; Sánchez, C.; Moreno, J.; Alves, F.; Alvar, J. Role of asymptomatic and symptomatic humans as reservoirs of visceral leishmaniasis in a Mediterranean context. PLoS. Negl. Trop. Dis. 2020, 14, e0008253. [Google Scholar] [CrossRef] [PubMed]
- Naucke, T.J.; Lorentz, S. First report of venereal and vertical transmission of canine leishmaniosis from naturally infected dogs in Germany. Parasit. Vectors 2012, 5, 1–5. [Google Scholar] [CrossRef] [Green Version]
- WHO. Health Topics: Leishmaniasis. Available online: https://www.who.int/health-topics/leishmaniasis (accessed on 4 April 2021).
- Millán, J.; Ferroglio, E.; Solano-Gallego, L. Role of wildlife in the epidemiology of Leishmania infantum infection in Europe. Parasitol. Res. 2014, 113, 2005–2014. [Google Scholar] [CrossRef]
- Molina, R.; Jiménez, M.I.; Cruz, I.; Iriso, A.; Martín- Martín, I.; Sevillano, O.; Melero, S.; Bernal, J. The hare (Lepus granatensis) as potential sylvatic reservoir of Leishmania infantum in Spain. Vet. Parasitol. 2012, 190, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Madrid, R.; Belinchón-Lorenzo, S.; Iniesta, V.; Fernández-Cotrina, J.; Parejo, J.C.; Serrano, F.J.; Monroy, I.; Baz, V.; Gómez-Luque, A.; Gómez-Nieto, L.C. First detection of Leishmania infantum kinetoplast DNA in hair of wild mammals: Application of qPCR method to determine potential parasite reservoirs. Acta Trop. 2013, 128, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Risueño, J.; Ortuño, M.; Pérez-cutillas, P.; Goyena, E.; Maia, C.; Cortes, S.; Campino, L.; Bernal, L.J.; Muñoz, C.; Arcenillas, I.; et al. Epidemiological and genetic studies suggest a common Leishmania infantum transmission cycle in wildlife, dogs and humans associated to vector abundance in Southeast Spain. Vet. Parasitol. 2018, 259, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Oleaga, A.; Zanet, S.; Espí, A.; Pegoraro de Macedo, M.R.; Gortázar, C.; Ferroglio, E. Leishmania in wolves in northern Spain: A spreading zoonosis evidenced by wildlife sanitary surveillance. Vet. Parasitol. 2018, 255, 26–31. [Google Scholar] [CrossRef]
- Azami-Conesa, I.; Martínez-Díaz, R.A.; González, F.; Gómez-Muñoz, M.T. First detection of Leishmania infantum in common urban bats Pipistrellus pipistrellus in Europe. Res. Vet. Sci. 2020, 132, 172–176. [Google Scholar] [CrossRef]
- Alcover, M.M.; Ribas, A.; Guillén, M.C.; Berenguer, D.; Tomás-Pérez, M.; Riera, C.; Fisa, R. Wild mammals as potential silent reservoirs of Leishmania infantum in a Mediterranean area. Prev. Vet. Med. 2020, 175, 104874. [Google Scholar] [CrossRef]
- Navea-Pérez, H.M.; Díaz- Sáez, V.; Corpas-López, V.; Merino-Espinosa, G.; Morillas-Márquez, F.; Martín- Sánchez, J. Leishmania infantum in wild rodents: Reservoirs or just irrelevant incidental hosts? Parasitol. Res. 2015, 114, 2363–2370. [Google Scholar] [CrossRef]
- Otranto, D.; Iatta, R.; Baneth, G.; Cavalera, M.A.; Bianco, A.; Parisi, A.; Dantas-Torres, F.; Colella, V.; McMillan-Cole, A.C.; Chomel, B. High prevalence of vector-borne pathogens in domestic and wild carnivores in Iraq. Acta Trop. 2019, 197, 105058. [Google Scholar] [CrossRef] [PubMed]
- Filioussis, G.; Petridou, E.; Papadopoulos, D.; Karavanis, E.; Morgan, E.; Billinis, C.; Papadopoulos, E. Hemorrhagic pneumonia in neonatal minks in Greece concomitant with Leismania infantum detection. Pol. J. Vet. Sci. 2018, 21, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Tsakmakidis, Ι.; Pavlou, C.; Tamvakis, A.; Papadopoulos, T.; Christodoulou, V.; Angelopoulou, K.; Dovas, C.I.; Antoniou, M.; Anastasakis, C.; Diakou, A. Leishmania infection in lagomorphs and minks in Greece. Vet. Parasitol. Reg. Stud. Reports. 2019, 16, 100279. [Google Scholar] [CrossRef]
- Regulation (EU) 2016/1141 of 13 July 2016 Adopting a List of Invasive Alien Species of Union Concern Pursuant to Regulation (EU) No 1143/2014 of the European Parliament and of the Council. Available online: http://data.europa.eu/eli/reg/2014/1143/oj (accessed on 25 June 2021).
- Regulation (EU) No 1143/2014 of the European Parliament and of the Council of 22 October 2014 on the Prevention and Management of the Introduction and Spread of Invasive Alien Species. Available online: http://data.europa.eu/eli/reg_impl/2016/1141/oj (accessed on 25 June 2021).
- Real Decreto 630/2013, de 2 de Agosto, por el que se Regula el Catálogo Español de Especies Exóticas Invasoras. Available online: https://www.boe.es/eli/es/rd/2013/08/02/630 (accessed on 25 June 2021).
- Decreto 213/2009, de 20 de Noviembre, del Consell, por el que se Aprueban Medidas Para el Control de Especies Exóticas Invasoras en la Comunitat Valenciana. [2009/13396]. Available online: https://dogv.gva.es/es/eli/es-vc/d/2009/11/20/213/ (accessed on 25 June 2021).
- Melero, Y.; Santulli, G.; Gómez, A.; Gosàlbez, J.; Rodríguez-Refojos, C.; Palazón, S. Morphological variation of introduced species: The case of American mink (Neovison vison) in Spain. Mamm. Biol. 2012, 77, 345–350. [Google Scholar] [CrossRef]
- Leary, S.; Underwood, W.; Anthony, R.; Cartner, S.; Grandin, T.; Greenacre, C.; Gwaltney-Brant, S.; McCrackin, M.A.; Meyer, R.; Miller, D.; et al. AVMA Guidelines for the Euthanasia of Animals: 2020 Edition; AVMA: Schaumburg, IL, USA, 2020; pp. 97–99. [Google Scholar]
- Lachaud, L.; Marchergui-Hammami, S.; Chabbert, E.; Dereure, J.; Dedet, J.P.; Bastien, P. Comparison of six PCR methods using peripheral blood for detection of canine visceral leishmaniasis. J. Clin. Microbiol. 2002, 40, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Souguir-Omrani, H.; Chemkhi, J.; Fathallah-Mili, A.; Saadi-BenAoun, Y.; BelHadjAli, I.; Guizani, I.; Guerbouj, S. Paraechinus aethiopicus (Ehrenberg 1832) and Atelerix algirus (Lereboullet 1842) hedgehogs: Possible reservoirs of endemic leishmaniases in Tunisia. Infect. Genet. Evol. 2018, 63, 219–230. [Google Scholar] [CrossRef] [PubMed]
- El Tai, N.O.; Osman, O.F.; El Fari, M.; Presber, W.; Schönian, G. Genetic heterogeneity of ribosomal internal transcribed spacer (ITS) in clinical samples of Leishmania donovani spotted on filter paper as revealed by single-strand conformation polymorphisms (sscp) and sequencing. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 1–5. [Google Scholar] [CrossRef]
- Van Eys, G.J.; Schoone, G.J.; Kroon, N.C.; Ebeling, S.B. Sequence analysis of small subunit ribosomal RNA genes and its use for detection and identification of Leishmania parasites. Mol. Biochem. Parasitol. 1992, 51, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Cruz, I.; Cañavate, C.; Rubio, J.M.; Morales, M.A.; Chicharro, C.; Laguna, F.; Jiménez- Mejías, M.; Sirera, G.; Videla, S.; Alvar, J. A nested polymerase chain reaction (Ln-PCR) for diagnosing and monitoring Leishmania infantum infection in coinfected patients with human immunodeficiency virus. Trans. R. Soc. Trop. Med. Hyg. 2002, 96, 185–189. [Google Scholar] [CrossRef]
- Piarroux, R.; Fontes, M.; Perasso, R.; Gambarelli, F.; Joblet, C.; Dumon, H.; Quilici, M. Phylogenetic relationships between Old World Leishmania strains revealed by analysis of a repetitive DNA sequence. Mol. Biochem. Parasitol. 1995, 73, 249–252. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control and European Food Safety Authority. Mosquito Maps. Stockholm: ECDC. 2018. Available online: https://www.ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data/phlebotomine-maps (accessed on 5 July 2021).
- Cardoso, L.; Schallig, H.; Persichetti, M.F.; Pennisi, M.G. New Epidemiological Aspects of Animal Leishmaniosis in Europe: The Role of Vertebrate Hosts Other than Dogs. Pathogens 2021, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Battisti, E.; Zanet, S.; Khalili, S.; Trisciuoglio, A.; Hertel, B.; Ferroglio, E. Molecular Survey on Vector-Borne Pathogens in Alpine Wild Carnivorans. Front. Vet. Sci. 2020, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Del Río, L.; Chitimia, L.; Cubas, A.; Victoriano, I.; De la Rúa, P.; Gerrikagoitia, X.; Barral, M.; Muñoz-García, C.I.; Goyena, E.; García- Martínez, D.; et al. Evidence for widespread Leishmania infantum infection among wild carnivores in L. infantum periendemic northern Spain. Prev. Vet. Med. 2014, 113, 430–435. [Google Scholar] [CrossRef]
- Giner, J.; Villanueva-Saz, S.; Alcover, M.M.; Riera, C.; Fisa, R.; Basurco, A.; Yzuel, A.; Trotta, M.; Fani, C.; Verde, M.T.; et al. Treatment and follow-up of a domestic ferret (Mustela putorius furo) with clinical leishmaniosis caused by Leishmania infantum. Vet. Parasitol. Reg. Stud. Rep. 2020, 21, 100423. [Google Scholar] [CrossRef]
- Chemkhi, J.; Souguir, H.; BelHadjAli, I.; Driss, M.; Guizani, I.; Guerbouj, S. Natural infection of Algerian hedgehog, Atelerix algirus (Lereboullet 1842) with Leishmania parasites in Tunisia. Acta Trop. 2015, 150, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Akhoundi, M.; Downing, T.; Votýpka, J.; Kuhls, K.; Lukeš, J.; Cannet, A.; Ravel, C.; Marty, P.; Delaunay, P.; Kasbari, M.; et al. Leishmania infections: Molecular targets and diagnosis. Mol. Aspects Med. 2017, 57, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, A.; Campino, L.; Cardoso, L.; Cortes, S. Evaluation of four molecular methods to detect Leishmania infection in dogs. Parasit. Vectors 2017, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, D.M.; Lonardoni, M.V.C.; Theodoro, U.; Silveira, T.G.V. Comparison of different primes for PCR-based diagnosis of cutaneous leishmaniasis. Braz. J. Infect. Dis. 2011, 15, 204–210. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Target | Oligonucleotides (5′-3′) | Expected Size of the Amplicon | PCR Conditions | N° Cycles | Reference |
|---|---|---|---|---|---|
| Repeat region | T2 (CGGCTTCGCACCATGCGGTG) B4 (ACATCCCTGCCCACATACGC) | 250 bp | 94 °C—30″ 61 °C—30″ 72 °C—15″ | 35 | [33] |
| kDNA | RV1 (CTTTTCTGGTCCCGCGGGTAGG) RV2 (CCACCTGGCCTATTTTACACCA) | 140 bp | 94 °C—30″ 62 °C—30″ 72 °C—20″ | 35 | [28] |
| kDNA | 13A (GTGGGGGAGGGGCGTTCT) 13B (ATTTTACACCAACCCCCAGTT) | 120 bp | 94 °C—30″ 60 °C—30″ 72 °C—20″ | 30 | [29] |
| ITS1 | LITSR (CTGGATCATTTTCCGATG) L5.8S (TGATACCACTTATCGCACTT) | 320 bp | 94 °C—30″ 53 °C—30″ 72 °C—15″ | 35 | [30] |
| SSU | Ext R221 (GGTTCCTTTCCTGATTTACG) Ext R332 (GGCCGGTAAAGGCCGAATAG) | 603 bp | 94 °C—30″ 60 °C—30″ 72 °C—30″ | 35 | [31] |
| Int R223 (TCCATCGCAACCTCGGTT) Int R333 (AAAGCGGGCGCGGTGCTG) | 358 bp | 94 °C—30″ 65 °C—30″ 72 °C—30″ | 32 | [32] |
| Animal ID | Date | Sex | Age | Weight (g) | Length (cm) | River |
|---|---|---|---|---|---|---|
| 68/16 | 8 March 2016 | F | Ad | 680 | 57 | Mijares |
| 26/16 | 10 February 2016 | F | Ad | 689 | 58 | Mijares |
| 192/16 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| 56/16 | 1 March 2016 | M | Ad | 1180 | 62 | Mijares |
| 23/16 | 10 February 2016 | F | Ad | 793 | 58.9 | Mijares |
| 197/16 | 20 October 2016 | M | Ad | 950 | 63 | Palancia |
| 66/16 | 6 March 2016 | F | Ad | 600 | 51 | Mijares |
| 201/16 | 22 October 2016 | F | Ad | 660 | 55.5 | Palancia |
| 55/16 | 1 March 2016 | M | Ad | 980 | 56 | Mijares |
| 41/16 | 18 February 2016 | M | Ad | 901 | 59 | Mijares |
| 62/16 | 4 March 2016 | F | Ad | 640 | 48 | Mijares |
| 1023/16 | 22 May 2016 | M | Juv | 878 | 62 | Turia |
| 77/16 | 14 March 2016 | M | Ad | 1180 | 64 | Mijares |
| 212/16 | 26 October 2016 | F | Ad | 650 | 58 | Palancia |
| 213/16 | 27 October 2016 | M | Ad | 810 | 58 | Palancia |
| 35/16 | 16 February 2016 | M | Ad | 1109 | 63.5 | Mijares |
| 29/16 | 14 February 2016 | F * | Ad | 721 | 57 | Mijares |
| 37/16 | 17 February 2016 | M | Ad | 1604 | 69 | Mijares |
| 30/16 | 14 February 2016 | F * | Ad | 583 | 52 | Mijares |
| 28/16 | 14 February 2016 | F | Ad | 637 | 56 | Mijares |
| 331/16 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| 1001/16 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Ten-Fold Dilutions from the Original DNA Sample | ||||||||
|---|---|---|---|---|---|---|---|---|
| PCR Protocol | DNA from Culture (20 µg/mL) | 10−1 | 10−2 | 10−3 | 10−4 | 10−5 | 10−6 | 10−7 |
| PCR Repeat region | + | + | + | + | + | + | − | − |
| PCR kDNA(RV1/RV2) | + | + | + | + | + | + | − | − |
| LnPCR SSU | + | + | + | + | + | + | − | − |
| PCR kDNA(13A/13B) | + | + | + | + | + | − | − | − |
| PCR ITS1 | + | + | + | + | + | − | − | − |
| Ten-Fold Dilutions from the Original DNA Sample | ||||||||
|---|---|---|---|---|---|---|---|---|
| PCR Protocol | Bone Marrow DNA | 10−1 | 10−2 | 10−3 | 10−4 | 10−5 | 10−6 | 10−7 |
| PCR Repeat region | + | + | + | + | + | − | − | − |
| PCR kDNA(RV1/RV2) | + | + | + | + | + | − | − | − |
| LnPCR SSU | + | + | + | − | − | − | − | − |
| PCR kDNA(13A/13B) | + | + | + | − | − | − | − | − |
| PCR ITS1 | + | + | + | + | − | − | − | − |
| Animal ID | PCR Repeat Region | LnPCR SSU | PCR kDNA (RV1/RV2) | PCR kDNA (13A/13B) | PCR ITS |
|---|---|---|---|---|---|
| 68/16 | + | − | − | − | − |
| 26/16 | + | − | − | − | − |
| 192/16 | + | − | − | − | − |
| 56/16 | + * | − | − | − | − |
| 23/16 | + | − | − | − | − |
| 197/16 | + | − | − | − | − |
| 66/16 | + | − | − | − | − |
| 201/16 | + | − | − | − | − |
| 55/16 | + | − | − | − | − |
| 41/16 | + | − | − | − | − |
| 62/16 | + | − | − | − | − |
| 1023/16 | + | − | − | − | − |
| 77/16 | + * | − | − | − | − |
| 212/16 | − | − | − | − | − |
| 213/16 | + * | − | − | − | − |
| 35/16 | − | − | − | − | − |
| 29/16 | + | + | − | − | − |
| 37/16 | + | − | − | − | − |
| 30/16 | + * | + | − | − | − |
| 28/16 | + | − | − | − | − |
| 331/16 | + | − | − | − | − |
| 1001/16 | + * | − | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azami-Conesa, I.; Sansano-Maestre, J.; Martínez-Díaz, R.A.; Gómez-Muñoz, M.T. Invasive Species as Hosts of Zoonotic Infections: The Case of American Mink (Neovison vison) and Leishmania infantum. Microorganisms 2021, 9, 1531. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071531
Azami-Conesa I, Sansano-Maestre J, Martínez-Díaz RA, Gómez-Muñoz MT. Invasive Species as Hosts of Zoonotic Infections: The Case of American Mink (Neovison vison) and Leishmania infantum. Microorganisms. 2021; 9(7):1531. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071531
Chicago/Turabian StyleAzami-Conesa, Iris, Jose Sansano-Maestre, Rafael Alberto Martínez-Díaz, and María Teresa Gómez-Muñoz. 2021. "Invasive Species as Hosts of Zoonotic Infections: The Case of American Mink (Neovison vison) and Leishmania infantum" Microorganisms 9, no. 7: 1531. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071531