
Composition and Corrosivity of Extracellular Polymeric Substances from the Hydrocarbon-Degrading Sulfate-Reducing Bacterium Desulfoglaeba alkanexedens ALDC

Abstract
:1. Introduction
2. Materials and Methods
2.1. Incubations
2.2. EPS Extraction
2.3. Field Emission Scanning Electron Microscopy (FE-SEM)
2.4. Confocal Microscopy
2.5. Nuclear Magnetic Resonance (NMR) Analysis
2.6. Corrosion Analysis
2.7. Polysaccharide Characterization
3. Results
3.1. D. alkanexedens Aggregation and Extracellular Material
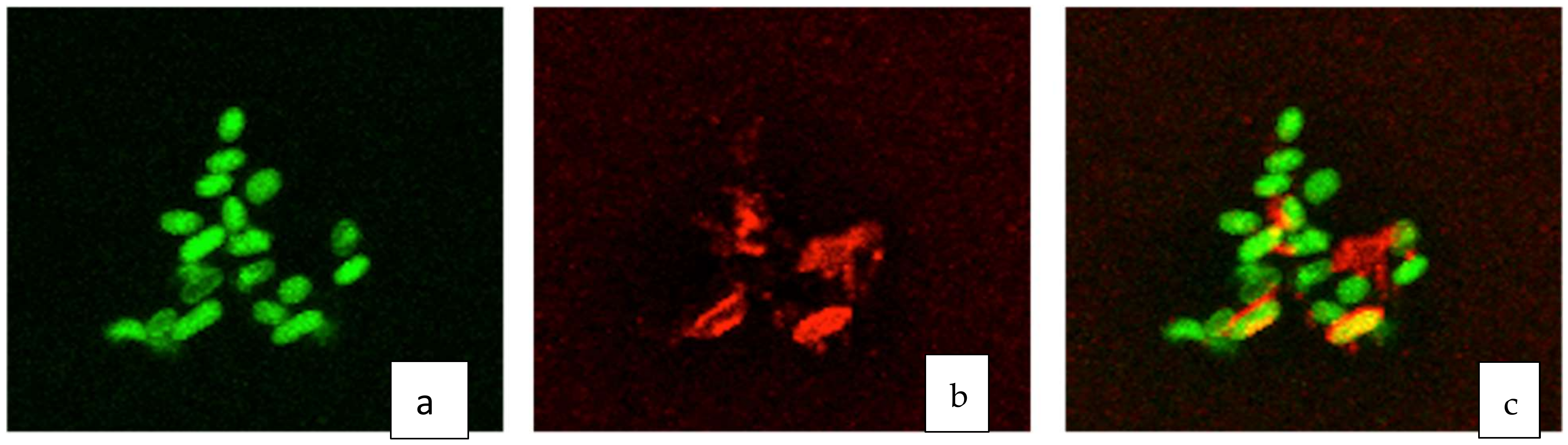
3.2. Fluorescent Microscopic Analysis of D. alkanexedens Aggregates
3.3. EPS Composition and Structure
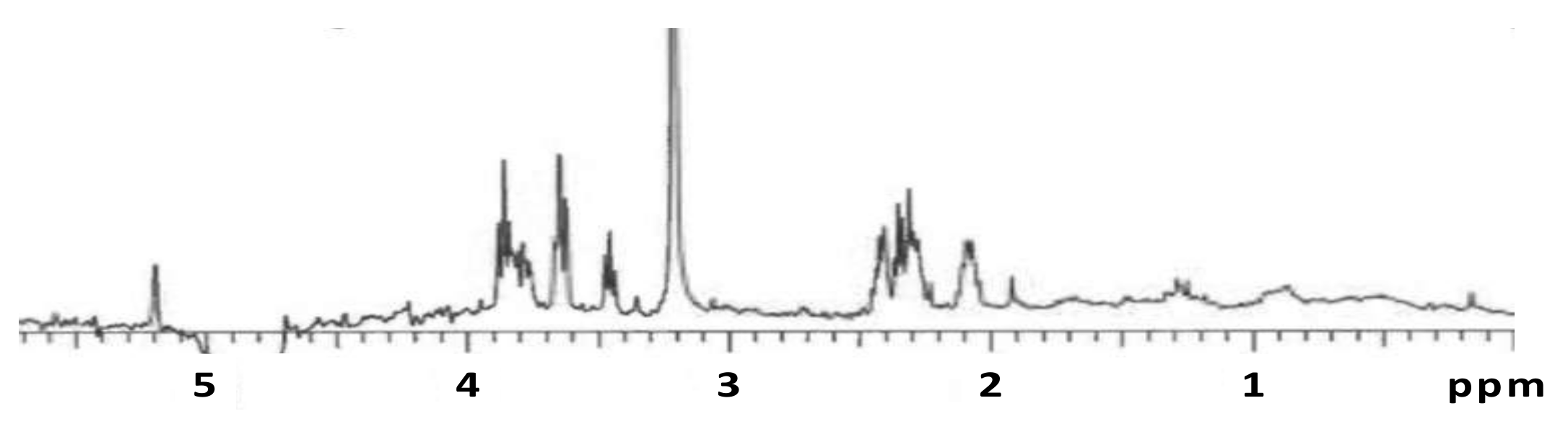
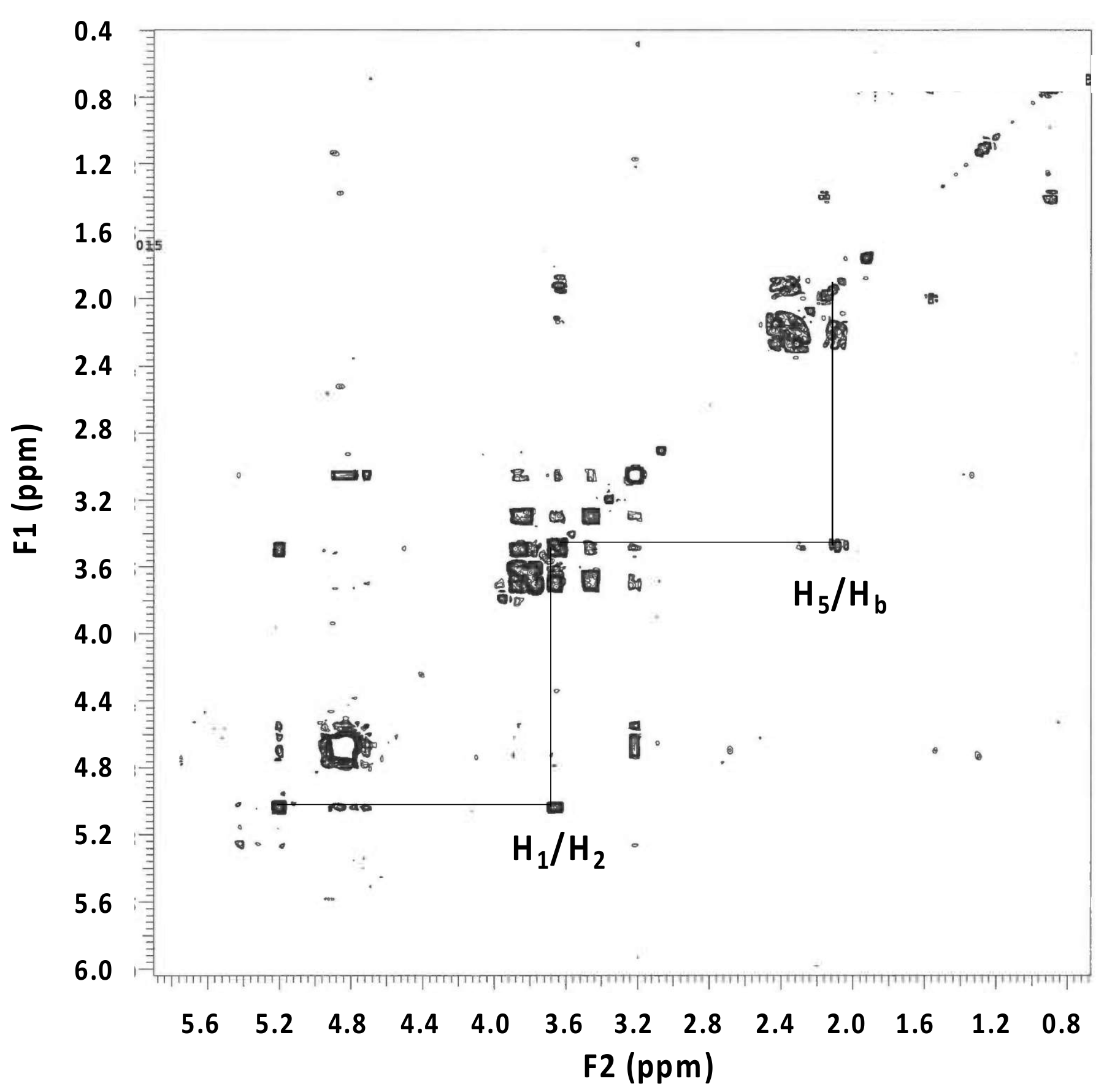
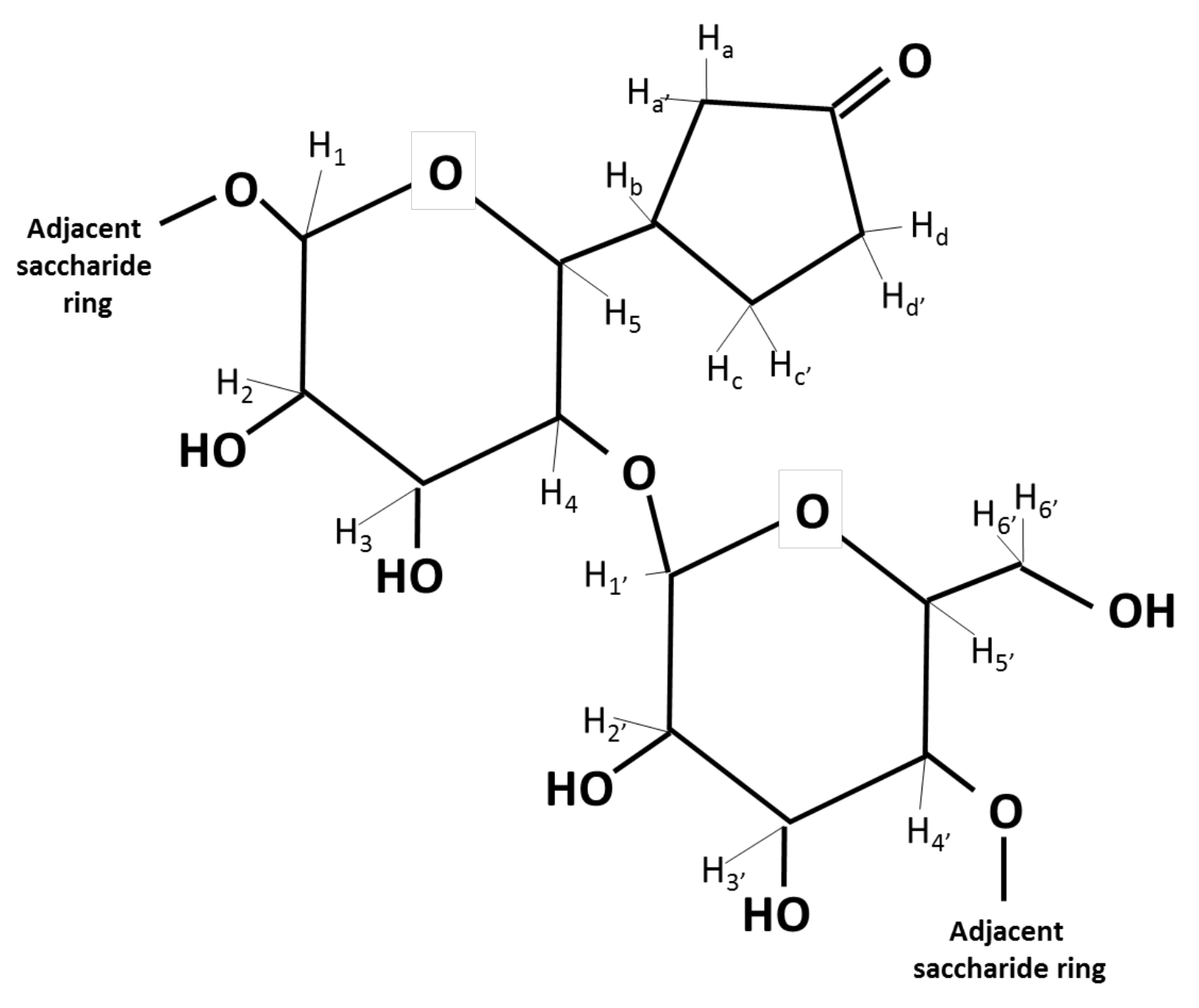
3.4. Nuclear Magnetic Resonance Analysis
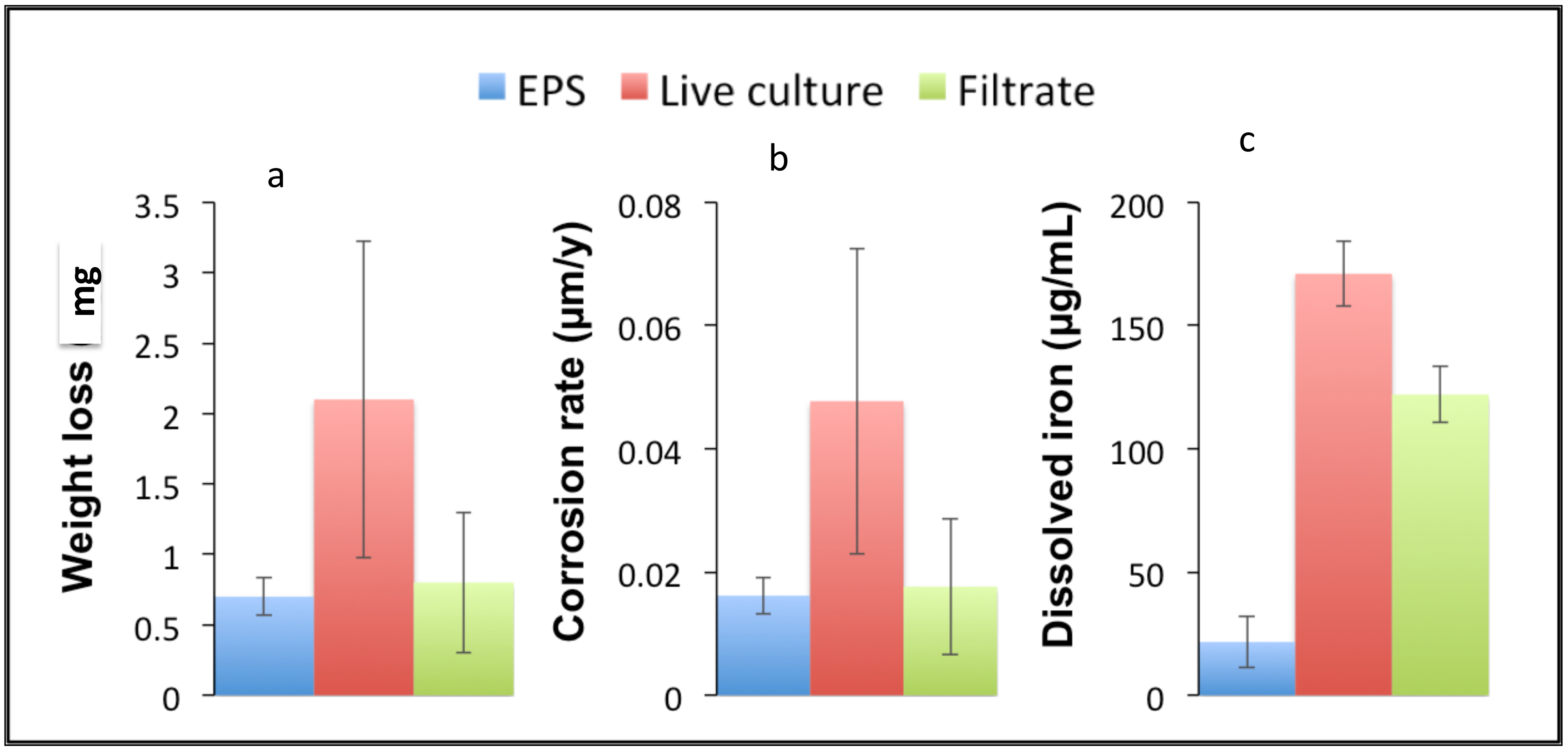
3.5. Assessment of D. alkanexedens EPS Corrosivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beech, I.; Sunner, J. Biocorrosion: Towards understanding interactions between biofilms and metals. Curr. Opin. Biotechnol. 2004, 15, 181–186. [Google Scholar] [CrossRef]
- Hamilton, W.A. Sulphate-reducing bacteria and anaerobic corrosion. Annu. Rev. Microbiol. 1985, 39, 195–217. [Google Scholar] [CrossRef] [PubMed]
- Kilbane, J.; Bogan, B.; Lamb, B. Quantifying the contribution of various bacterial groups to microbiologically influenced corrosion. In Proceedings of the NACE International Corrosion Conference, Houston, TX, USA, 3–7 April 2005. Paper 05491. [Google Scholar]
- Branda, S.S.; Vik, S.; Friedman, L.; Kolter, R. Biofilms: The matrix revisited. Trends. Microbiol. 2005, 13, 20–26. [Google Scholar] [CrossRef]
- Flemming, H.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Wingender, J.; Neu, T.R.; Flemming, H.-C. What are Bacterial Extracellular Polymeric Substances? In Microbial Extracellular Polymeric Substances; Wingender, J., Neu, T.R., Eds.; Springer: Heidelberg, Germany, 1999; pp. 1–19. [Google Scholar] [CrossRef]
- Kolter, R.; Greenberg, E.P. The superficial life of microbes. Nature 2006, 441, 300–302. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, T.; Morris, G.; Green, D.H. Yield and physicochemical properties of EPS from Halomonas sp. strain TG39 identifies a role for protein and anionic residues (sulfate and phosphate) in emulsification of n-hexadecane. Biotechnol. Bioeng. 2009, 103, 207–216. [Google Scholar] [CrossRef]
- More, T.T.; Yadav, J.S.S.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Extracellular polymeric substances of bacteria and their potential environmental applications. J. Environ. Manag. 2014, 144, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Arias, S.; Del Moral, A.; Ferrer, M.R.; Tallon, R.; Quesada, E.; Béjar, V. Mauran, an exopolysaccharide produced by the halophilic bacterium Halomonas maura, with a novel composition and interesting properties for biotechnology. Extremophiles 2003, 7, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, I.W. Biosynthesis and Composition of Gram-Negative Bacterial Extracellular and Wall Polysaccharides. Annu. Rev. Microbiol. 1985, 39, 243–270. [Google Scholar] [CrossRef]
- Gutierrez, T.; Berry, D.; Yang, T.; Mishamandani, S.; McKay, L.; Teske, A.; Aitken, M.D. Role of Bacterial Exopolysaccharides (EPS) in the Fate of the Oil Released during the Deepwater Horizon Oil Spill. PLoS ONE 2013, 8, e67717. [Google Scholar] [CrossRef]
- Gutierrez, T.; Mulloy, B.; Black, K.; Green, D.H. Glycoprotein emulsifiers from two marine Halomonas species: Chemical and physical characterization. J. Appl. Microbiol. 2007, 103, 1716–1727. [Google Scholar] [CrossRef]
- Hauser, L.J.; Land, M.L.; Brown, S.D.; Larimer, F.; Keller, K.L.; Rapp-Giles, B.J.; Price, M.N.; Lin, M.; Bruce, D.C.; Detter, J.C. Complete genome sequence and updated annotation of Desulfovibrio alaskensis G20. J. Bacteriol. 2011, 193, 4268–4269. [Google Scholar] [CrossRef] [Green Version]
- Feio, M.J.; Zinkevich, V.; Beech, I.B.; Llobet-Brossa, E.; Eaton, P.; Schmitt, J.; Guezennec, J. Desulfovibrio alaskensis sp. nov., a sulphate-reducing bacterium from a soured oil reservoir. Int. J. Syst. Evol. Microbiol. 2004, 54, 1747–1752. [Google Scholar] [CrossRef] [Green Version]
- Zinkevich, V.; Bogdarina, I.; Kang, H.; Hill, M.A.W.; Tapper, R.; Beech, I.B. Characterisation of exopolymers produced by different isolates of marine sulphate-reducing bacteria. Int. Biodeterior. Biodegrad. 1996, 37, 163–172. [Google Scholar] [CrossRef]
- Beech, I.B.; Zinkevich, V.; Tapper, R.; Gubner, R. Direct involvement of an extracellular complex produced by a marine sulfate-reducing bacterium in deterioration of steel. Geomicrobiol. J. 1998, 15, 121–134. [Google Scholar] [CrossRef]
- Davidova, I.A.; Duncan, K.E.; Choi, O.K.; Suflita, J.M. Desulfoglaeba alkanexedens gen. nov., sp. nov. an n-alkane-degrading, sulfate-reducing bacterium. Int. J. Syst. Evol. Microbiol. 2006, 56, 2737–2742. [Google Scholar] [CrossRef] [PubMed]
- Lyles, C.N.; Le, H.M.; Beasley, W.H.; McInerney, M.J.; Suflita, J.M. Anaerobic hydrocarbon and fatty acid metabolism by syntrophic bacteria and their impact on carbon steel corrosion. Front. Microbiol. 2014, 5, 114. [Google Scholar] [CrossRef]
- Widdel, F.; Bak, F. Gram-negative mesophilic sulfate-reducing bacteria. In The Prokaryote; Balows, A., Trüper, H.G., Eds.; Springer: New York, NY, USA, 1992; pp. 3352–3378. [Google Scholar] [CrossRef]
- Liang, R.; Aktas, D.F.; Aydin, E.; Bonifay, V.; Sunner, J.; Suflita, J. Anaerobic Biodegradation of Alternative Fuels and Associated Biocorrosion of Carbon Steel in Marine Environments. Environ. Sci. Technol. 2016, 50, 4844–4853. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Phillips, E.J.P. Organic matter mineralization with reduction of ferric iron in anaerobic sediments. Appl. Environ. Microbiol. 1986, 51, 683–689. [Google Scholar] [CrossRef] [Green Version]
- ASTM. G1-03 Standard Practice for Preparing, Cleaning and Evaluating Corrosion Test Specimens; ASTM: West Conshohocken, PA, USA, 2003. [Google Scholar]
- Herbert, D.; Phipps, P.J. Chemical Analysis of Microbial Cells. In Methods in Microbiology; Norris, J.R., Ribbons, D.W., Eds.; Academic Press: New York, NY, USA, 1971; Volume 5B, pp. 209–344. [Google Scholar] [CrossRef]
- Bubb, W.A. NMR Spectroscopy in the Study of Carbohydrates: Characterizing the Structural Complexity. Concepts Magn. Reson. Part A Educ. J. 2003, 19, 1–19. [Google Scholar] [CrossRef]
- Data Organic Chemistry. Available online: https://organicchemistrydata.org/hansreich/resources/nmr/?index=nmr_index%2F1H_shift (accessed on 15 March 2021).
- Nilsson, G.S.; Gorton, L.; Bergquist, K.-E.; Nilsson, U. Determination of the Degree of Branching in Normal and Amylopectin Potato Starch with 1H-NMR Spectroscopy Improved Resolution and Two-Dimensional Spectroscopy. Starch 1996, 48, 352–357. [Google Scholar] [CrossRef]
- Bouchez-Naitali, M.; Blanchet, D.; Bardin, V.; Vandecasteele, J.-P. Evidence for interfacial uptake in hexadecane degradation by Rhodococcus equi: The importance of cell flocculation. Microbiology 2001, 147, 2537–2543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, R.S.; Finnerty, W.R.; Sudarsanan, K.; Young, R.A. Microbial assimilation of hydrocarbons. I. The fine structure of a hydrocarbon oxidizing Acinetobacter sp. Arch. Microbiol. 1975, 102, 75–78. [Google Scholar] [CrossRef]
- Müller-Hurtig, R.; Wagner, F. Biosurfactants for environmental control. In Biosurfactant: Production, Properties, Applications. Surfactant Science Series; Kosaric, N., Ed.; Marcel Dekker: New York, NY, USA, 1993; Volume 48, pp. 447–469. [Google Scholar]
- Reddy, P.G.; Singh, H.D.; Roy, P.K.; Baruah, J.N. Predominant role of hydrocarbon solubilization in the microbial uptake of hydrocarbons. Biotechol. Bioeng. 1982, 24, 1241–1269. [Google Scholar] [CrossRef]
- Baumgarten, T.; Sperling, S.; Seifert, J.; Von Bergen, M.; Steiniger, F.; Wick, L.Y.; Heipieper, H.J. Membrane vesicle formation as a multiple-stress response mechanism enhances Pseudomonas putida DOT-T1E cell surface hydrophoibicity and biofilm formation. Appl. Environ. Microbiol. 2012, 78, 6217–6224. [Google Scholar] [CrossRef] [Green Version]
- Gonzales-Gil, G.; Thomas, L.; Emwas, A.-H.; Lens, P.N.L.; Saikaly, P.E. NMR and MALDI-TOF MS based characterization of exopolysaccharides in anaerobic microbial aggregates from full-scale reactors. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Shaw, D.R.; Zhang, L.; Haroon, M.F.; Narita, Y.; Emwas, A.-H.; Saikaly, P.E.; Okabe, S. Aggregation ability of three phylogenetically distant anammox bacterial species. Water Res. 2018, 143, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Videla, H.A.; Herrera, L.K. Understanding of microbial inhibition of corrosion: A comprehensive overview. Int. Biodeter. Biodegrad. 2009, 63, 896–900. [Google Scholar] [CrossRef]
- Vigneron, A.; Alsop, E.B.; Chambers, B.; Lomans, B.; Head, I.M.; Tsesmetzis, N. Complementary microorganisms in highly corrosive biofilms from offshore oil production facility. Appl. Environ. Microbiol. 2016, 82, 2545–2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kip, N.; Van Veen, J.A. The dual role of microbes in corrosion. ISME J. 2015, 9, 542–551. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1H NMR Chemical Shift Values (ppm) | |||
|---|---|---|---|
| Experimental NMR | Predicted NMR | Starch in D2O | |
| H1 | 5.19 | 4.58 | 5.35 |
| H2 | 3.66 | 3.24 | 3.63 |
| H3 | 3.87 | 3.65 | 3.94 |
| H4 | 3.47 | 3.49 | 3.62 |
| H5 | 3.62 | 3.52 | 3.82 |
| Hb | 2.10 | 2.32 | |
| Ha Ha′ | 2.36 | 2.35, 2.45 | |
| Hd Hd′ | 2.42 | 2.42, 2.35 | |
| Hc Hc′ | 2.34 | 2.48, 2.27 | |
| H1′ | 5.19 | 4.49 | 5.35 |
| H2′ | 3.66 | 3.20 | 3.63 |
| H3′ | 3.62 | 3.60 | 3.94 |
| H4′ | 3.47 | 3.16 | 3.62 |
| H5′ | 3.87 | 3.46 | 3.82 |
| H6′ H6′ | 3.76 | 3.81, 3.81 | 3.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davidova, I.A.; Lenhart, T.R.; Nanny, M.A.; Suflita, J.M. Composition and Corrosivity of Extracellular Polymeric Substances from the Hydrocarbon-Degrading Sulfate-Reducing Bacterium Desulfoglaeba alkanexedens ALDC. Microorganisms 2021, 9, 1994. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091994
Davidova IA, Lenhart TR, Nanny MA, Suflita JM. Composition and Corrosivity of Extracellular Polymeric Substances from the Hydrocarbon-Degrading Sulfate-Reducing Bacterium Desulfoglaeba alkanexedens ALDC. Microorganisms. 2021; 9(9):1994. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091994
Chicago/Turabian StyleDavidova, Irene A., Tiffany R. Lenhart, Mark A. Nanny, and Joseph M. Suflita. 2021. "Composition and Corrosivity of Extracellular Polymeric Substances from the Hydrocarbon-Degrading Sulfate-Reducing Bacterium Desulfoglaeba alkanexedens ALDC" Microorganisms 9, no. 9: 1994. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091994