
Geographical Relationships between Long-Tailed Goral (Naemorhedus caudatus) Populations Based on Gut Microbiome Analysis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Microbiota Analysis
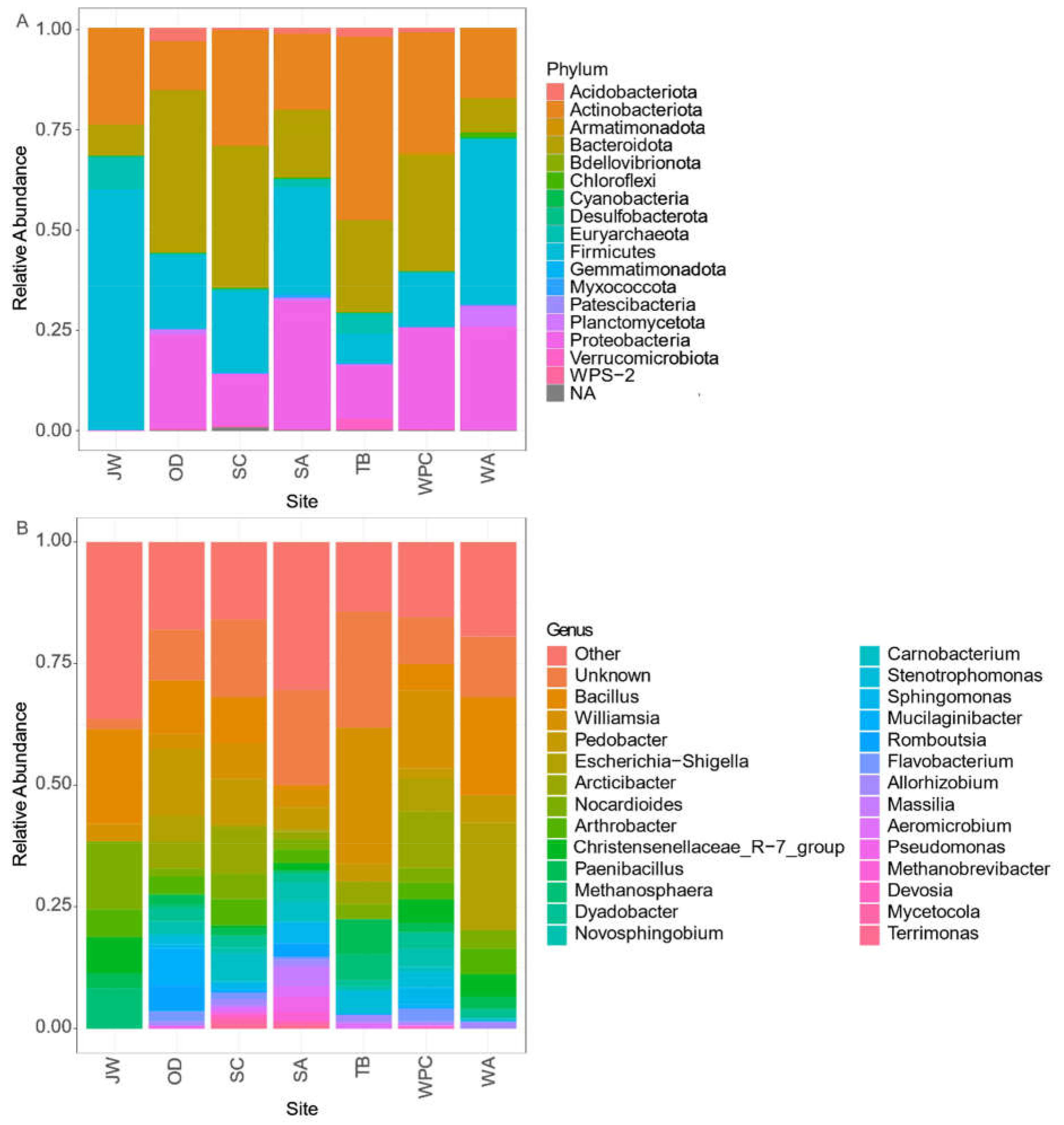
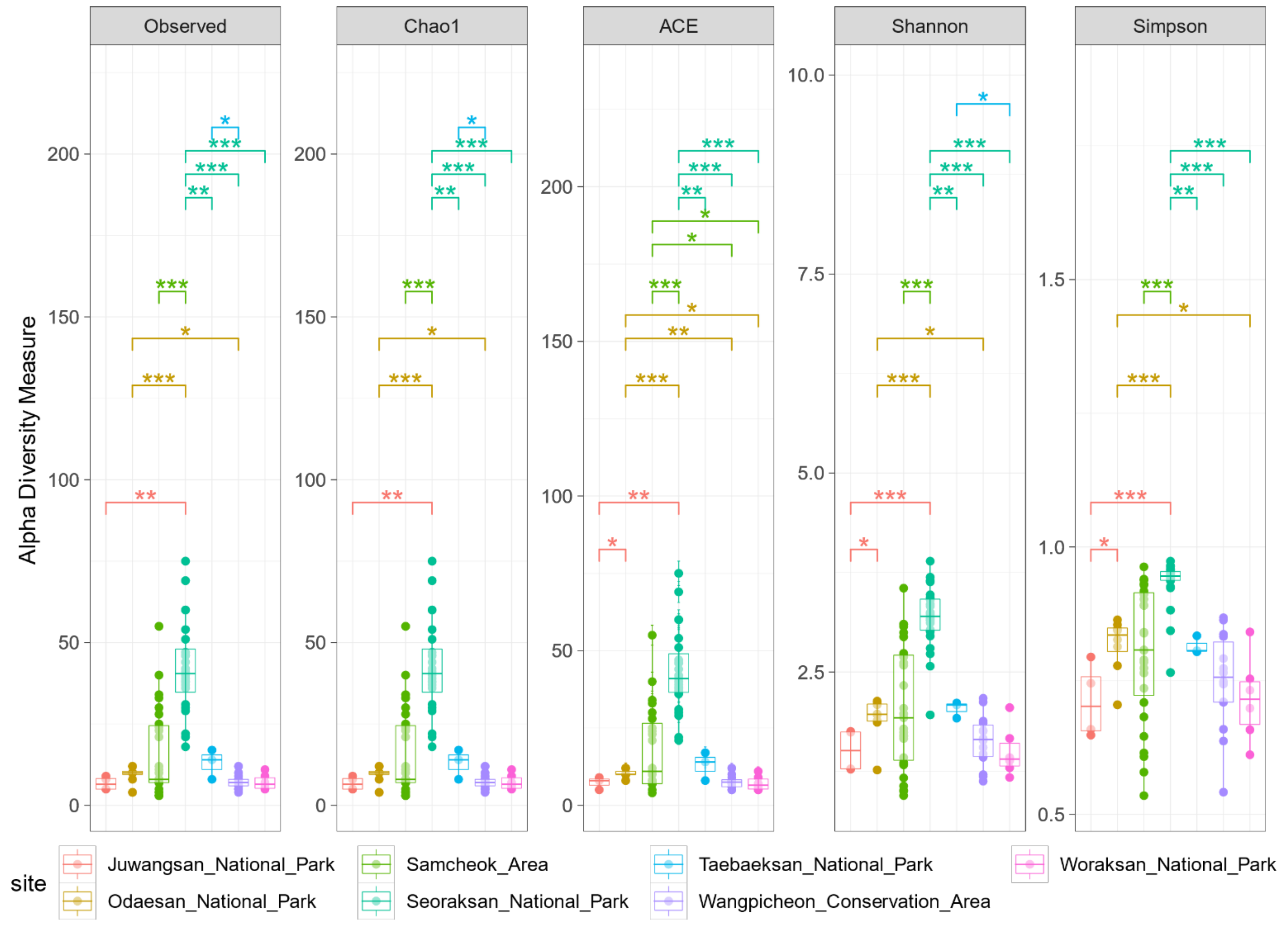
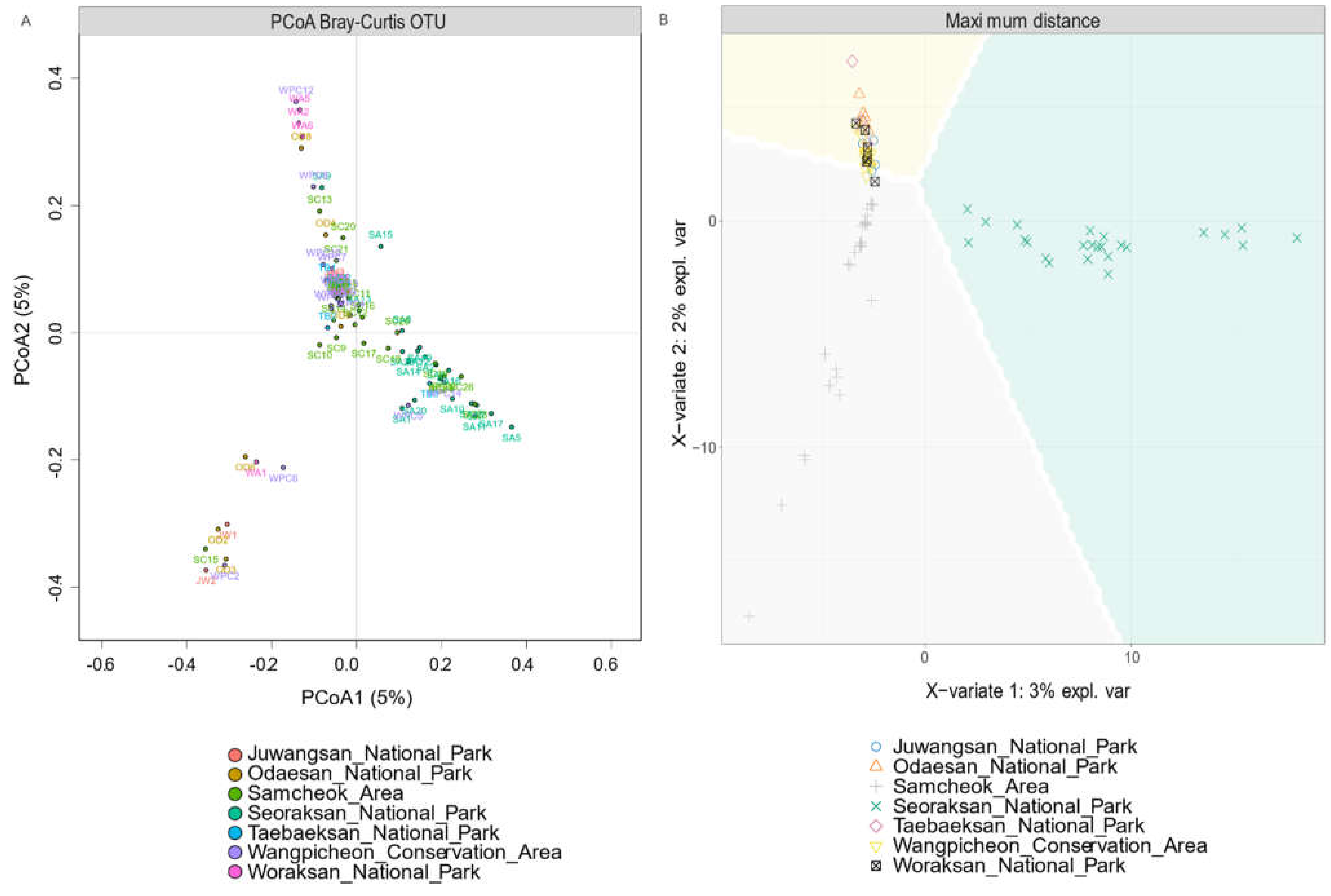
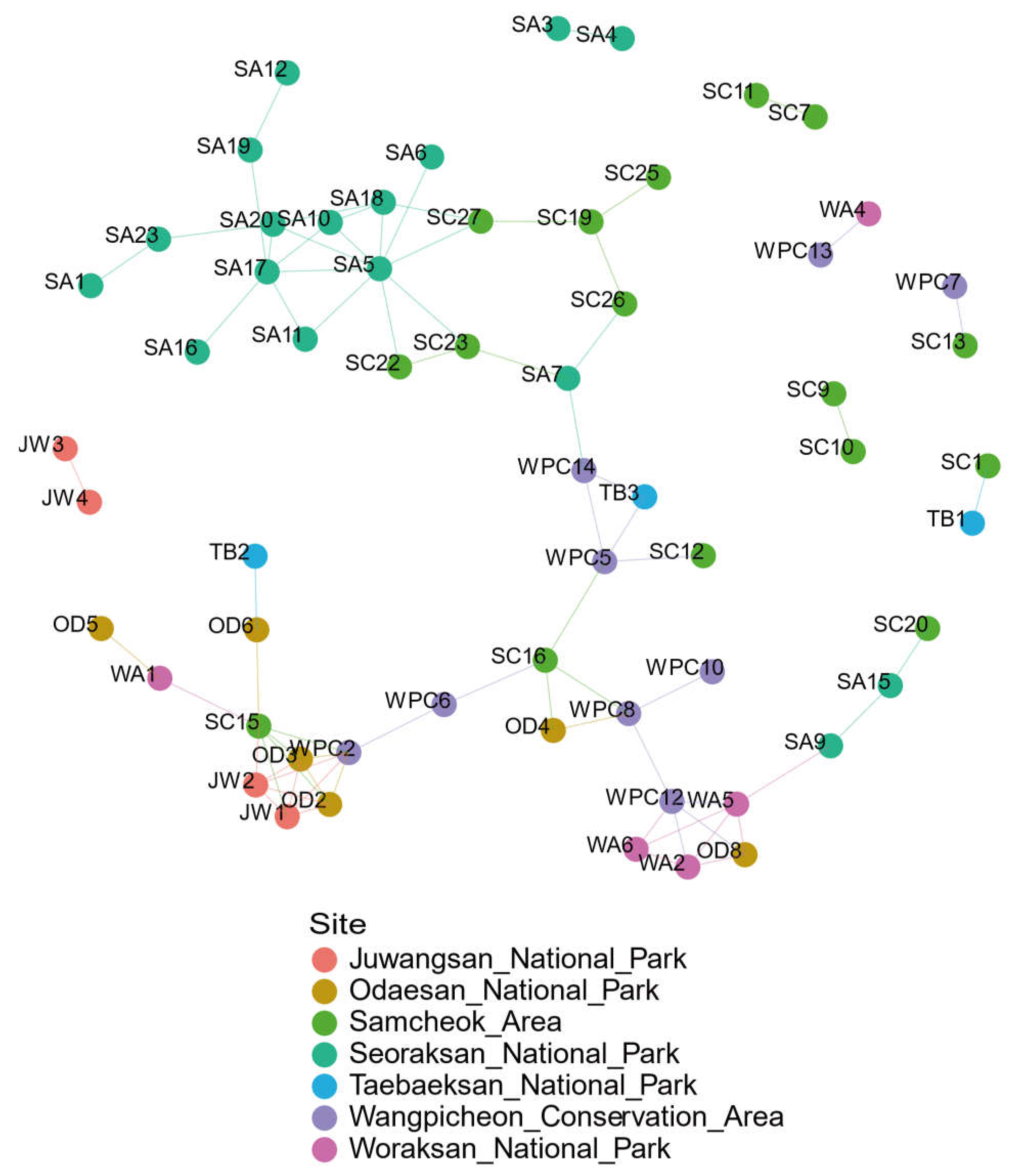
3. Results
3.1. Biodiversity Patterns
3.2. Site-Relational Network Analysis
4. Discussion
4.1. Biodiversity and Regional Relationships
4.2. Home Range, Dispersal, and Migration
4.3. Site-Relational Network Analysis and Regional Migration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bragina, E.; Kim, S.; Zaumyslova, O.; Park, Y.S.; Lee, W. Naemorhedus caudatus. The IUCN Red List of Threatened Species. 2020, 2020: E.T14295A22150540. Available online: https://www.iucnredlist.org/species/14295/22150540 (accessed on 4 August 2021).
- Wilson, D.E.; Reeder, D.M. (Eds.) Mammal Species of the World: A Taxonomic and Geographic Reference; The Johns Hopkins University Press: Baltimore, MD, USA, 2005; Volume 1. [Google Scholar]
- Choi, S.K.; Chun, S.; An, J.; Lee, M.Y.; Kim, H.J.; Min, M.S.; Kwon, S.W.; Choi, T.Y.; Lee, H.; Kim, K.S. Genetic diversity and population structure of the long-tailed goral, Naemorhedus caudatus, in South Korea. Genes Genet. Syst. 2015, 90, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Park, H.B.; Hong, S. Habitat characteristics coincidence of dead and living long-tailed gorals (Naemorhedus caudatus) according to extreme snowfall. Animals 2021, 11, 997. [Google Scholar] [CrossRef]
- Rawls, J.F.; Mahowald, M.A.; Ley, R.E.; Gordon, J.I. Reciprocal gut microbiota transplants from zebrafish and mice to germ-free recipients reveal host habitat selection. Cell 2006, 127, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Li, J.; Yu, Y.; Wang, J.; He, Z.; Van Nostrand, J.D.; Kempher, M.L.; Wu, L.; Wang, Y.; Liao, L.; et al. Environmental filtering decreases with fish development for the assembly of gut microbiota. Environ. Microbiol. 2016, 18, 4739–4754. [Google Scholar] [CrossRef]
- Burns, A.R.; Stephens, W.Z.; Stagaman, K.; Wong, S.; Rawls, J.F.; Guillemin, K.; Bohannan, B.J. Contribution of neutral processes to the assembly of gut microbial communities in the zebrafish over host development. ISME J. 2016, 10, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Goertz, S.; de Menezes, A.B.; Birtles, R.J.; Fenn, J.; Lowe, A.E.; MacColl, A.D.; Poulin, B.; Young, S.; Bradley, J.E.; Taylor, C.H. Geographical location influences the composition of the gut microbiota in wild house mice (Mus musculus domesticus) at a fine spatial scale. PLoS ONE 2019, 14, e0222501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.B.; Han, C.W.; Hong, S. Camera trapping of long-tailed goral (Naemorhedus caudatus) in BaekAm and Geumjong Mountains, South Korea. J. For. Environ. Sci. 2018, 34, 71–76. [Google Scholar]
- Cho, C.U.; Kim, K.C.; Kwon, G.H.; Kim, K.Y.; Lee, B.K.; Son, J.I. Habitat use of reintroduced long-tailed gorals (Naemorhedus caudatus) in Woraksan (Mt.) National Park in Korea. Korean J. Environ. Ecol. 2015, 29, 184–191. [Google Scholar] [CrossRef]
- Cho, C.U.; Gyun, G.H.; Yang, J.J.; Lim, S.J.; Lee, A.N.; Park, H.B.; Lee, B.K. Home range and behavioral characteristics of the endangered Korea Gorals (Naemorhedus caudatus) with GPS collar. Korean J. Environ. Ecol. 2014, 28, 1–9. [Google Scholar] [CrossRef]
- Yang, B.; Wang, Y.; Qian, P.Y. Sensitivity and correlation of hypervariable regions in 16S rRNA genes in phylogenetic analysis. BMC Bioinform. 2016, 17, 135. [Google Scholar] [CrossRef] [Green Version]
- Churchill, J.D.; King, J.L.; Chakraborty, R.; Budowle, B. Effects of the Ion PGM™ Hi-Q™ sequencing chemistry on sequence data quality. Int. J. Leg. Med. 2016, 130, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- De Summa, S.; Lasorella, A.; Strippoli, S.; Giudice, G.; Guida, G.; Elia, R.; Nacchiero, E.; Azzariti, A.; Silvestris, N.; Guida, M.; et al. The genetic germline background of single and multiple primary melanomas. Front. Mol. Biosci. 2020, 7, 555630. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.; Beiko, R.G. 16S rRNA gene analysis with QIIME2. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2018; Volume 1849, pp. 113–129. [Google Scholar]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Mcmurdie, P.J.; Holmes, S. Phyloseq: A bioconductor package for handling and analysis of high-throughput phylogenetic sequence data. In Pacific Symposium on Biocomputing. Pac. Symp. Biocomput. 2012, 2012, 235–246. [Google Scholar]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.A. MixOmics: An R package for omics feature selection and multiple data integration. PLOS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [Green Version]
- Couch, C.E.; Stagaman, K.; Spaan, R.S.; Combrink, H.J.; Sharpton, T.J.; Beechler, B.R.; Jolles, A.E. Diet and gut microbiome enterotype are associated at the population level in African buffalo. Nat. Commun. 2021, 12, 2267. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hundertmark, K.J. Home Range, Dispersal and Migration. Ecology and Management of the North American Moose; Smithsonian Institution Press: Washington, DC, USA, 1998; pp. 303–335. [Google Scholar]
- Cho, C.; Kim, K.; Kwon, G. Habitat altitude and home range of the endangered long-tailed goral (Naemorhedus caudatus): Seasonal and monthly home range and altitude change. Mammalia 2016, 80, 481–489. [Google Scholar] [CrossRef]
- Jianping, W.; Yong, Z.; Dongfeng, F.; Wenxu, L. Comparison of home range of Siberian musk deer and goral in winter. J. Northeast. For. Uuniversity-Chin. Edition 2008, 36, 58. [Google Scholar]
- Shupei, T.A.N.G.; Wu, L.; Zhiqing, H.A.N.; Yingying, H.A.N.; Weidong, B.A.O. A Pilot Study on Home Range and Habitat Use of Chinese Goral (Naemorhedus Griseus): Exploring GPS Tracking Data in Cliff Landscape by Three Estimation Methods. Res. Sq. 2021. preprint. [Google Scholar] [CrossRef]
- Cho, C.U.; Kim, K.Y.; Kim, K.C.; Kim, H.M.; An, J.Y.; Lee, B.K.; Park, J.G. Home range analysis of a pair of gorals (Naemorhedus caudatus) using GPS collar according to the elevation change, in the North Gyeongbuk Province (Uljin) of Korea. J. Korean Assoc. Geogr. Inf. Stud. 2015, 18, 135–146. [Google Scholar] [CrossRef]
- Cho, C.U.; Kim, K.C.; Kim, K.Y.; Lee, Y.H.; Son, J.I.; Jung, S.J.; Song, D.J.; Jung, D.H. Analysis of home range change of female goral (Naemorhedus caudatus) during the period of birth and management plan for their protection in the natural habitat in Koreas. Korean J. Environ. Ecol. 2016, 30, 857–864. [Google Scholar] [CrossRef]
- Kropil, R.; Smolko, P.; Garaj, P. Home range and migration patterns of male red deer Cervus elaphus in Western Carpathians. Eur. J. Wildl. Res. 2015, 61, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Takii, A.; Izumiyama, S.; Mochizuki, T.; Okumura, T.; Sato, S. Seasonal migration of sika deer in the Oku-Chichibu Mountains, central Japan. Mammal Study 2012, 37, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Ramanzin, M.; Sturaro, E.; Zanon, D. Seasonal migration and home range of roe deer (Capreolus capreolus) in the Italian eastern Alps. Can. J. Zool. 2007, 85, 280–289. [Google Scholar] [CrossRef]
- Wahlström, L.K.; Liberg, O. Patterns of dispersal and seasonal migration in roe deer (Capreolus capreolus). J. Zool. 1995, 235, 455–467. [Google Scholar] [CrossRef]
- Sah, P.; Méndez, J.D.; Bansal, S. A multi-species repository of social networks. Sci. Data 2019, 6, 44. [Google Scholar] [CrossRef] [Green Version]
- Franks, D.W.; Ruxton, G.D.; James, R. Sampling animal association networks with the gambit of the group. Behav. Ecol. Sociobiol. 2010, 64, 493–503. [Google Scholar] [CrossRef]
- Kim, B.J. Simple assessment of taxonomic status and genetic diversity of Korean long-tailed goral (Naemorhedus caudatus) based on partial mitochondrial cytochrome b gene using non-invasive fecal samples. Proc. Natl. Inst. Ecol. Repub. Korea 2021, 2, 32–41. [Google Scholar]
- Yang, J.; Zhu, G.F.; Jiang, J.; Xiang, C.L.; Gao, F.L.; Bao, W.D. Non-invasive genetic analysis indicates low population connectivity in vulnerable Chinese gorals: Concerns for segregated population management. Zool. Res. 2019, 40, 439–448. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, C.E.; Cho, B.-J.; Kim, M.-J.; Park, H.C.; Shin, J.-H. Geographical Relationships between Long-Tailed Goral (Naemorhedus caudatus) Populations Based on Gut Microbiome Analysis. Microorganisms 2021, 9, 2002. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9092002
Park CE, Cho B-J, Kim M-J, Park HC, Shin J-H. Geographical Relationships between Long-Tailed Goral (Naemorhedus caudatus) Populations Based on Gut Microbiome Analysis. Microorganisms. 2021; 9(9):2002. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9092002
Chicago/Turabian StylePark, Chang Eon, Bum-Joon Cho, Min-Ji Kim, Hee Cheon Park, and Jae-Ho Shin. 2021. "Geographical Relationships between Long-Tailed Goral (Naemorhedus caudatus) Populations Based on Gut Microbiome Analysis" Microorganisms 9, no. 9: 2002. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9092002