
Sperm Nuclei Analysis and Nuclear Organization of a Fertile Boar–Pig Hybrid by 2D FISH on Both Total and Motile Sperm Fractions

 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sampling
2.2. Freezing Protocol of Epididymal Spermatozoa
2.3. Percoll Gradient
2.4. Sperm DNA Integrity
2.5. Slide Preparation and Decondensation
2.6. Probe Preparation and Sperm FISH Analysis
2.7. Statistical Analysis
3. Results
3.1. Sperm DNA Integrity
3.2. FISH Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Halbert, N.D.; Derr, J.N. A comprehensive evaluation of cattle introgression into US federal bison herds. J. Hered. 2007, 98, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pauciullo, A.; Knorr, C.; Perucatti, A.; Iannuzzi, A.; Iannuzzi, L.; Erhardt, G. Characterization of a very rare case of living ewe-buck hybrid using classical and molecular cytogenetics. Sci. Rep. 2016, 6, 34781. [Google Scholar] [CrossRef] [Green Version]
- Iannuzzi, A.; Pereira, J.; Iannuzzi, C.; Fu, B.; Ferguson-Smith, M. Pooling strategy and chromosome painting characterize a living zebroid for the first time. PLoS ONE 2017, 12, e0180158. [Google Scholar] [CrossRef] [Green Version]
- Ryder, O.A.; Chemnick, L.G.; Bowling, A.T.; Benirschke, K. Male Mule Foal Qualifies as the Offspring of a Female Mule and Jack Donkey. J. Hered. 1985, 76, 379–381. [Google Scholar] [PubMed]
- Iacolina, L.; Pertoldi, C.; Amills, M.; Kusza, S.; Megens, H.-J.; Bâlteanu, V.A.; Bakan, J.; Cubric-Curik, V.; Oja, R.; Saarma, U.; et al. Hotspots of recent hybridization between pigs and wild boars in Europe. Sci. Rep. 2018, 8, 17372. [Google Scholar] [CrossRef] [PubMed]
- Jori, F.; Relun, A.; Trabucco, B.; Charrier, F.; Maestrini, O.; Chavernac, D.; Cornelis, D.; Casabianca, F.; Etter, E.M.C. Questionnaire-Based Assessment o Wild Boar/Domestic Pig Interactions and Implications for Disease Risk Management in Corsica. Front. Vet. Sci. 2017, 4, 198. [Google Scholar] [CrossRef] [Green Version]
- Tanihara, F.; Hirata, M.; Iizuka, S.; Sairiki, S.; Nii, M.; Nguyen, N.T.; Le, Q.A.; Hirano, T.; Otoi, T. Relationship among ovarian follicular status, developmental competence of oocytes, and anti-Mullerian hormone levels: A comparative study in Japanese wild boar crossbred gilts and Large White gilts. Anim. Sci. J. 2019, 90, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.P.; Jost, L. Sperm chromatin structure assay is useful for fertility assessment. Methods Cell Sci. 2000, 22, 169–189. [Google Scholar] [CrossRef] [PubMed]
- Kawase, Y.; Wada, N.A.; Jishage, K. Evaluation of DNA fragmentation of freeze-dried mouse sperm using a modified sperm chromatin structure assay. Theriogenology 2009, 72, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.P. The Sperm Chromatin Structure Assay (SCSA®) and other sperm DNA fragmentation tests for evaluation of sperm nuclear DNA integrity as related to fertility. Anim. Reprod. Sci. 2016, 169, 56–75. [Google Scholar] [CrossRef] [Green Version]
- Di Dio, C.; Longobardi, V.; Zullo, G.; Parma, P.; Pauciullo, A.; Perucatti, A.; Higgins, J.; Iannuzzi, A. Analysis of meiotic segregation by triple-color fish on both total and motile sperm fractions in a t(1p;18) river buffalo bull. PLoS ONE 2020, 15, e0232592. [Google Scholar] [CrossRef] [PubMed]
- Massei, G.; Kindberg, J.; Licoppe, A.; Gačić, D.; Šprem, N.; Kamler, J.; Baubet, E.; Hohmann, U.; Monaco, A.; Ozoliņš, J.; et al. Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Manag. Sci. 2015, 71, 492–500. [Google Scholar] [CrossRef]
- Canu, A.; Costa, S.; Iacolina, L.; Piatti, P.; Apollonio, M.; Scandura, M. Are captive wild boar more introgressed than free-ranging wild boar? Two case studies in Italy. Eur. J. Wildl. Res. 2014, 60, 459–467. [Google Scholar] [CrossRef]
- Pittiglio, C.; Khomenko, S.; Beltran-Alcrudo, D. Wild boar mapping using population-density statistics: From polygons to high resolution raster maps. PLoS ONE 2018, 13, e0193295. [Google Scholar] [CrossRef] [Green Version]
- Rivero, M.J.; Rodríguez-Estévez, V.; Pietrosemoli, S.; Carballo, C.; Cooke, A.S.; Kongsted, A.G. Forage Consumption and Its Effects on the Performance of Growing Swine-Discussed in Relation to European Wild Boar (Sus scrofa L.) in Semi-Extensive Systems: A Review. Animals 2019, 9, 457. [Google Scholar] [CrossRef] [Green Version]
- Ciotola, F.; Albarella, S.; Scopino, G.; Carpino, S.; Monaco, F.; Peretti, V. Crossbreeding effect on genome stability in pig (Sus scrofa scrofa). Folia Biol. 2014, 62, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Guastella, A.M.; Criscione, A.; Marletta, D.; Zuccaro, A.; Chies, L.; Bordonaro, S. Molecular characterization and genetic structure of the Nero Siciliano pig breed. Genet. Mol. Biol. 2010, 33, 650–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzi, F.; Gliozzi, T.; Cerolini, S.; Maldjian, A.; Zaniboni, L.; Parodi, L.; Gandini, G. Semen quality of Italian local pig breeds. Ital. J. Anim. Sci. 2005, 4, 482–484. [Google Scholar] [CrossRef]
- Turri, F.; Gliozzi, T.M.; Gandini, G.; Pizzi, F. Constitution of Nero Siciliano pig breed genetic reserve: Flow cytometry evaluation of epididymal sperm cells. Ital. J. Anim Sci. 2015, 14, 150. [Google Scholar]
- Genualdo, V.; Perucatti, A.; Marletta, D.; Castiglioni, B.; Bordonaro, S.; Iannaccone, M.; Ciotola, F.; Peretti, V.; Iannuzzi, A. Cytogenetic investigation in two endangered pig breeds raised in Southern-Italy: Clinical and environmental aspects. Livest. Sci. 2018, 216, 36–43. [Google Scholar] [CrossRef]
- Suzuki, K.; Nagai, T. In vitro fertility and motility characteristics of frozen–thawed boar epididymal spermatozoa separated by Percoll. Theriogenology 2003, 60, 1481–1494. [Google Scholar] [CrossRef]
- Evenson, D.P.; Darzynkiewicz, Z.; Melamed, M.R. Relation of mammalian sperm chromatin heterogeneity to fertility. Science 1980, 210, 1131–1133. [Google Scholar] [CrossRef] [PubMed]
- Sellem, E.; Broekhuijse, M.; Chevrier, L.; Camugli, S.; Schmitt, E.; Schibler, L.; Koenen, E. Use of combinations of in vitro quality assessments to predict fertility of bovine semen. Theriogenology 2015, 84, 1447–1454.e5. [Google Scholar] [CrossRef]
- Gliozzi, T.M.; Turri, F.; Manes, S.; Cassinelli, C.; Pizzi, F. The combination of kinetic and flow cytometric semen parameters as a tool to predict fertility in cryopreserved bull semen. Animals 2017, 11, 1975–1982. [Google Scholar] [CrossRef] [PubMed]
- Toschi, P.; Capra, E.; A Anzalone, D.; Lazzari, B.; Turri, F.; Pizzi, F.; A Scapolo, P.; Stella, A.; Williams, J.L.; Marsan, P.A.; et al. Maternal peri-conceptional undernourishment perturbs offspring sperm methylome. Reproduction 2020, 159, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Eggen, A.; Gautier, M.; Billaut, A.; Petit, É.; Hayes, H.; Laurent, P.; Urban, C.; Pfister-Genskow, M.; Eilertsen, K.; Bishop, M.D. Construction and characterization of a bovine BAC library with four genome-equivalent coverage. Genet. Sel. Evol. 2001, 33, 543–548. [Google Scholar] [CrossRef]
- Iannuzzi, A.; Braun, M.; Genualdo, V.; Perucatti, A.; Reinartz, S.; Proios, I.; Heppelmann, M.; Rehage, J.; Hülskötter, K.; Beineke, A.; et al. Clinical, cytogenetic and molecular genetic characterization of a tandem fusion translocation in a male Holstein cattle with congenital hypospadias and a ventricular septal defect. PLoS ONE 2020, 15, e0227117. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2007. [Google Scholar]
- Ahmadi, A.; Ng, S.-C. Developmental capacity of damaged spermatozoa. Hum. Reprod. 1999, 14, 2279–2285. [Google Scholar] [CrossRef]
- Bollwein, H.; Fuchs, I.; Koess, C. Interrelationship Between Plasma Membrane Integrity, Mitochondrial Membrane Potential and DNA Fragmentation in Cryopreserved Bovine Spermatozoa. Reprod. Domest. Anim. 2008, 43, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Didion, B.A.; Kasperson, K.M.; Wixon, R.L.; Evenson, D.P. Boar Fertility and Sperm Chromatin Structure Status: A Retrospective Report. J. Androl. 2009, 30, 655–660. [Google Scholar] [CrossRef] [Green Version]
- Hernández, M.; Roca, J.; Ballester, J.; Vazquez, J.M.; Martínez, E.A.; Johannisson, A.; Saravia, F.; Rodriguez-Martinez, H. Differences in SCSA outcome among boars with different sperm freezability. Int. J. Androl. 2006, 29, 583–591. [Google Scholar] [CrossRef]
- Iannuzzi, A.; Della Valle, G.; Russo, M.; Longobardi, V.; Albero, G.; De Canditiis, C.; Kosior, M.A.; Pistucci, R.; Gasparrini, B. Evaluation of bovine sperm telomere length and association with semen quality. Theriogenology 2020, 158, 227–232. [Google Scholar] [CrossRef]
- Noguchi, M.; Yoshioka, K.; Hikono, H.; Iwagami, G.; Suzuki, C.; Kikuchi, K. Centrifugation on Percoll density gradient enhances motility, membrane integrity and in vitro fertilizing ability of frozen-thawed boar sperm. Zygote 2015, 23, 68–75. [Google Scholar] [CrossRef]
- Matás, C.; Vieira, L.; García-Vázquez, F.; Avilés-López, K.; López-Úbeda, R.; Carvajal, J.; Gadea, J. Effects of centrifugation through three different discontinuous Percoll gradients on boar sperm function. Anim. Reprod. Sci. 2011, 127, 62–72. [Google Scholar] [CrossRef]
- Martecikova, S.; Hulinska, P.; Reckova, Z.; Pavlik, A.; Jeseta, M.; Machatkova, M. Effect of acrosome reaction progress in frozen-thawed boar spermatozoa on the efficiency of in vitro oocyte fertilization. Veterinární Medicína 2010, 55, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Larson, K.; Brannian, J.; Timm, B.; Jost, L.; Evenson, D. Density gradient centrifugation and glass wool filtration of semen remove spermatozoa with damaged chromatin structure. Hum. Reprod. 1999, 14, 2015–2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, S.; Johannisson, A.; Wahlsten, T.; Ijas, R.; Andersson, M.; Rodriguez-Martinez, H. Sperm chromatin structure and sperm morphology: Their association with fertility in AI-dairy Ayrshire sires. Theriogenology 2013, 79, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Vozdová, M.; Kasikova, K.; Oracova, E.; Prinosilova, P.; Rybar, R.; Horinova, V.; Gaillyová, R.; Rubes, J. The effect of the swim-up and hyaluronan-binding methods on the frequency of abnormal spermatozoa detected by FISH and SCSA in carriers of balanced chromosomal translocations. Hum. Reprod. 2012, 27, 930–937. [Google Scholar] [CrossRef] [Green Version]
- Ducos, A.; Pinton, A.; Yerle, M.; Séguéla, A.; Berland, H.M.; Brun-Baronnat, C.; Bonnet, N.; Darré, R. Cytogenetic and molecular characterization of eight new reciprocal translocations in the pig species. Estimation of their incidence in French populations. GSE 2002, 34, 389–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubeš, J.; Vozdová, M.; Kubíčková, S. Aneuploidy in pig sperm: Multicolor fluorescence in situ hybridization using probes for chromosomes 1, 10, and Y. Cytogenet. Genome Res. 1999, 85, 200–204. [Google Scholar] [CrossRef] [PubMed]
- O Zalensky, A.; Allen, M.J.; Kobayashi, A.; A Zalenskaya, I.; Balhórn, R.; Bradbury, E.M. Well-defined genome architecture in the human sperm nucleus. Chromosoma 1995, 103, 577–590. [Google Scholar] [CrossRef]
- Cremer, T.; Cremer, C. Chromosome territories, nuclear architecture and gene regulation in mammalian cells. Nat. Rev. Genet. 2001, 2, 292–301. [Google Scholar] [CrossRef]
- Parada, L.; Misteli, T. Chromosome positioning in the interphase nucleus. Trends Cell Biol. 2002, 12, 425–432. [Google Scholar] [CrossRef]
- Parada, L.A.; McQueen, P.G.; Misteli, T. Tissue-specific spatial organization of genomes. Genome Biol. 2004, 5, R44. [Google Scholar] [CrossRef] [Green Version]
- Foster, H.A.; Abeydeera, L.R.; Griffin, D.K.; Bridger, J.M. Non-random chromosome positioning in mammalian sperm nuclei, with migration of the sex chromosomes during late spermatogenesis. J. Cell Sci. 2005, 118, 1811–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acloque, H.; Bonnet-Garnier, A.; Mompart, F.; Pinton, A.; Yerle-Bouissou, M. Sperm nuclear architecture is locally modified in presence of a Robertsonian translocation t(13;17). PLoS ONE 2013, 8, e78005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiland, E.; Zegalo, M.; Kurpisz, M. Interindividual differences and alterations in the topology of chromosomes in human sperm nuclei of fertile donors and carriers of reciprocal translocations. Chromosome Res. 2008, 16, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Parada, L.A.; McQueen, P.G.; Munson, P.J.; Misteli, T. Conservation of relative chromosome positioning in normal and cancer cells. Mol. Biol. Cell. 2002, 13, 246a-a. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
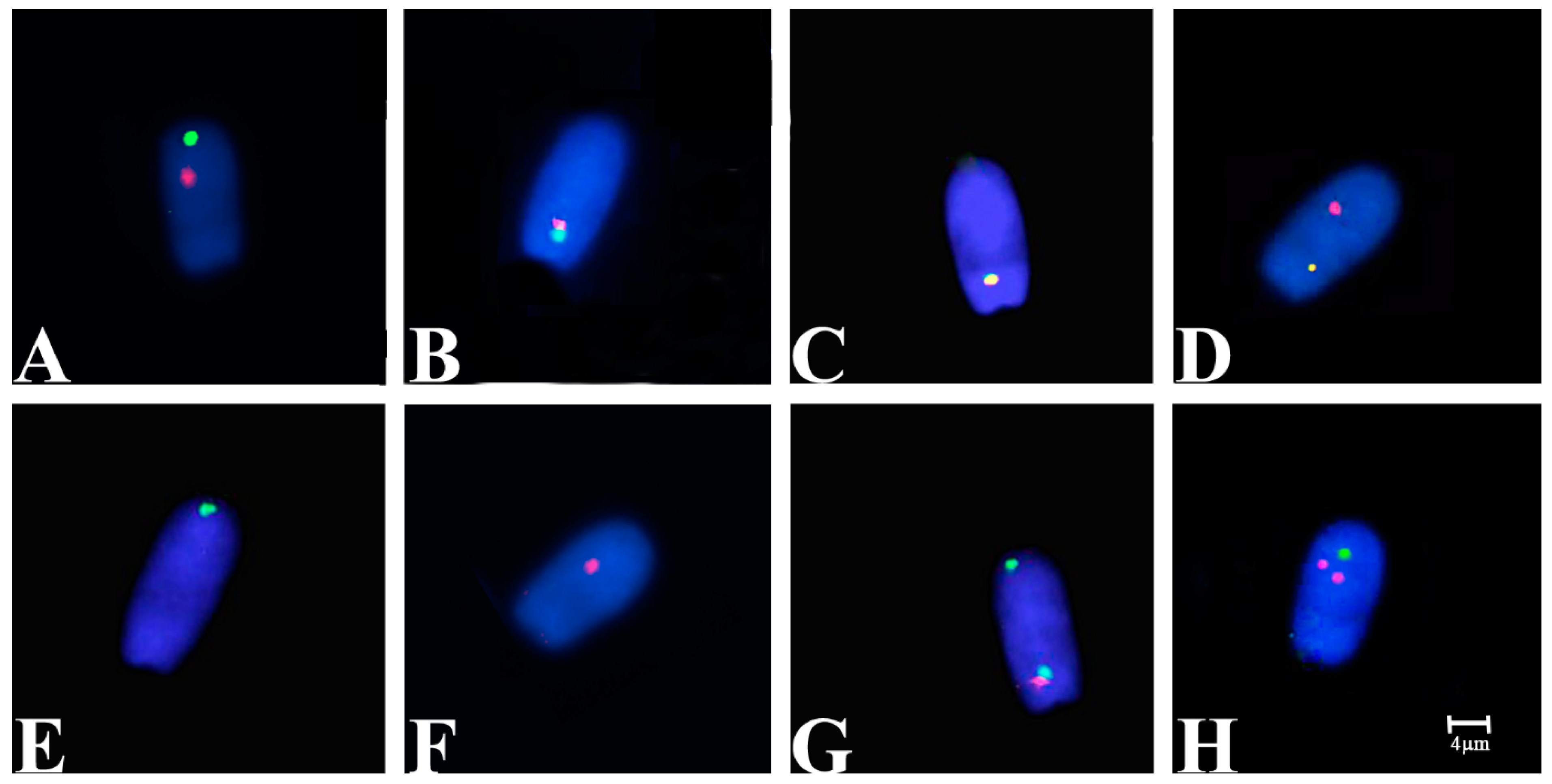{kind=link}
{kind=link}
| Probe | Ssc CH-242 Genome Localization (bp) | Chromosomal Localization | Label |
|---|---|---|---|
| 1st Pool | |||
| bI 0038D20 | 3,378,177–3,735,904 | ||
| bI 00379A02 | 4,104,594–4,360,253 | Ssc 15q12 | Biotin |
| bI 0508M05 | 4,664,773–4,976,019 | ||
| 2nd Pool | |||
| bI 0501J06 | 4,740,641–5,086,289 | ||
| bI 0069P06 | 5,160,276–5,425,104 | Ssc 17q12 | Digoxigenin |
| bI 0465A06 | 5,711,917–5,976,153 |
| Variables | Control | Hybrid | ||
|---|---|---|---|---|
| TSF | MSF | TSF | MSF | |
| %DFI (%) | 2.03 ± 0.3437 aA | 1.58 ± 0.3437 aA | 2.51 ± 0.3437 aA | 0.87 ± 0.3437 bA |
| %HG (%) | 0.11 ± 0.0268 aA | 0.20 ± 0.0268 bA | 0.34 ± 0.0268 aB | 0.90 ± 0.0268 bB |
| Fluorescent Signals a | Segregation | Chromosomal Constitution | Associated Genotype | % Of Combinations (Number of Gametes Investigated) | |||
|---|---|---|---|---|---|---|---|
| Hybrid TSF | Control TSF | Hybrid MSF | Control MSF | ||||
| G-R | Alternate | Ssc 15-17 | n = 19 | 26.73 (2673) | 96.05 (1921) | 66.63 (6663) | 96.70 (1934) |
| 49.61 b | 97.96 b | 74.16 b | 98.17 b | ||||
| GR | rob(15;17) | n = 18 | 23.97 (2397) | 2 (40) | 20.21 (2021) | 1.25 (25) | |
| 44.48 b | 2 b | 22.49 b | 1.26 b | ||||
| Y | rob(15;17) | n = 18 | 3.18 (318) | 0 (0) | 3.00 (301) | 0.55 (11) | |
| 5.90 b | 0 b | 3.35 b | 0.55 b | ||||
| tot | 53.88 (5388) | 98.05 (1961) | 89.85 (8987) | 98.50 (1970) | |||
| R-GR (Y) | Adjacent I | Ssc 17; rob(15;17) | n = 18 | 21.98 (2198) | 0.90 (18) | 2.040 (204) | 0.85 (17) |
| G | Ssc 15 | n = 19 | 10.10 (1010) | 0 (0) | 4.10 (401) | 0 (0) | |
| tot | 32.08 (3208) | 0.90 (18) | 6.04 (605) | 0.85 (17) | |||
| R | Adjacent II | Ssc 17 | n = 19 | 9.97 (997) | 0 (0) | 4.10 (410) | 0.65 (13) |
| G-GR | Ssc 15; rob(15;17) | n = 18 | 1.03 (103) | 1.05 (21) | 0 (0) | 0 (0) | |
| tot | 11.0 (1100) | 1.05 (21) | 4.09 (410) | 0.65 (13) | |||
| G-R-R | 3:1 | Ssc 15; Ssc 17; SSC 17 | n = 19 | 3.04 (304) | 0 (0) | 0 (0) | 0 (0) |
| Total | 10,000 | 2000 | 10,000 | 2000 | |||
| Alternate Segregations | TSF n (%) | MSF n (%) |
|---|---|---|
| Normal (G-R) | 2673 (50) A | 6663 (74) B |
| rob(15;17) (GR or Y) | 2715 (50) A | 2443 (26) B |
| Total | 5388 | 8985 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genualdo, V.; Turri, F.; Pizzi, F.; Castiglioni, B.; Marletta, D.; Iannuzzi, A. Sperm Nuclei Analysis and Nuclear Organization of a Fertile Boar–Pig Hybrid by 2D FISH on Both Total and Motile Sperm Fractions. Animals 2021, 11, 738. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030738
Genualdo V, Turri F, Pizzi F, Castiglioni B, Marletta D, Iannuzzi A. Sperm Nuclei Analysis and Nuclear Organization of a Fertile Boar–Pig Hybrid by 2D FISH on Both Total and Motile Sperm Fractions. Animals. 2021; 11(3):738. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030738
Chicago/Turabian StyleGenualdo, Viviana, Federica Turri, Flavia Pizzi, Bianca Castiglioni, Donata Marletta, and Alessandra Iannuzzi. 2021. "Sperm Nuclei Analysis and Nuclear Organization of a Fertile Boar–Pig Hybrid by 2D FISH on Both Total and Motile Sperm Fractions" Animals 11, no. 3: 738. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030738