
Effect of Different Kefir Source on Fermentation, Aerobic Stability, and Microbial Community of Alfalfa Silage




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Forage and Silage Preparation
2.2. Chemical Analysis
2.3. Microbial Populations
2.4. Aerobic Stability Analysis
2.5. Microbial Diversity Analysis
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, X.; Wen, A.; Dong, Z.; Desta, S.T.; Shao, T. Effects of formic acid and potassium diformate on the fermentation quality, chemical composition and aerobic stability of alfalfa silage. Grass Forage Sci. 2017, 72, 833–839. [Google Scholar] [CrossRef]
- Jiang, D.; Li, B.; Zheng, M.; Niu, D.; Zuo, S.; Xu, C. Effects of Pediococcus pentosaceus on fermentation, aerobic stability and microbial communities during ensiling and aerobic spoilage of total mixed ration silage containing alfalfa (Medicago sativa L.). Grassl. Sci. 2020, 66, 215–224. [Google Scholar] [CrossRef]
- Mc Garvey, J.A.; Franco, R.B.; Palumbo, J.D.; Hnasko, R.; Stanker, L.; Mitloehner, F.M. Bacterial population dynamics during the ensiling of Medicago sativa (alfalfa) and subsequent exposure to air. J. Appl. Microbiol. 2013, 114, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yu, Z.; Wang, X.; Tian, J. Effects of inoculants and environmental temperature on fermentation quality and bacterial diversity of alfalfa silage. Anim. Sci. J. 2018, 89, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.S.; Bai, J.; Li, F.H.; Xu, D.M.; Zhang, Y.X.; Bu, D.P.; Zhao, L.S. Effects of malate, citrate, succinate and fumarate on fermentation, chemical composition, aerobic stability and digestibility of alfalfa silage. Anim. Feed. Sci. Technol. 2020, 268, 114604. [Google Scholar] [CrossRef]
- Zhao, S.S.; Wang, Y.P.; Yang, F.Y.; Wang, Y.; Zhang, H. Screening a Lactobacillus plantarum strain for good adaption in alfalfa ensiling and demonstrating its improvement of alfalfa silage quality. J. Appl. Microbiol. 2020, 129, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Zhou, H.; Zhang, N.; Si, B.; Tu, Y.; Ma, T.; Diao, Q. Effects of different source additives and wilt conditions on the pH value, aerobic stability, and carbohydrate and protein fractions of alfalfa silage. Anim. Sci. J. 2017, 88, 99–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnürer, J.; Jonsson, A. Pichia anomala J121: A 30-year overnight near success biopreservation story. Antonie Van Leeuwenhoek 2011, 99, 5–12. [Google Scholar] [CrossRef]
- Chen, R.; Yang, S.; Zhang, L.; Zhou, Y.J. Advanced strategies for production of natural products in yeast. iScience 2020, 23, 100879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Liu, T.; Ye, T.; Yang, X.; Xue, Y.; Shen, Y.; Zhang, Q.; Zheng, X. Effect of lactic acid bacteria and yeasts on the structure and fermentation properties of Tibetan kefir grains. Int. Dairy J. 2021, 114, 104943. [Google Scholar] [CrossRef]
- da Silva, M.S.B.; Okura, M.H. Produtos à base de kefir desenvolvidos e estudados no Brasil. Res. Soc. Dev. 2021, 10, e19010716491. [Google Scholar] [CrossRef]
- John, S.M.; Deeseenthum, S. Properties and benefits of kefir—A review. Songklanakarin J. Sci. Technol. 2015, 37, 275–282. [Google Scholar]
- Kim, D.H.; Jeong, D.; Kim, H.; Kang, I.B.; Chon, J.W.; Song, K.Y.; Seo, K.H. Antimicrobial activity of kefir against various food pathogens and spoilage bacteria. Korean J. Food Sci. Anim. Resour. 2016, 36, 787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhao, X.; Jiang, Y.; Zhao, W.; Guo, T.; Cao, Y.; Teng, J.; Hao, X.; Zhao, J.; Yang, Z. Antioxidant status and gut microbiota change in an aging mouse model as influenced by exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibetan kefir. J. Dairy Sci. 2017, 100, 6025–6041. [Google Scholar] [CrossRef] [PubMed]
- Gonda, M.; Garmendia, G.; Rufo, C.; León Peláez, Á.; Wisniewski, M.; Droby, S.; Vero, S. Biocontrol of Aspergillus flavus in ensiled sorghum by water kefir microorganisms. Microorganisms 2019, 7, 253. [Google Scholar] [CrossRef] [Green Version]
- Okuyucu, B.; Esen, S. Is it possible to improve the fermentation and nutritional quality of wheat straw silage by replacing commercial inoculant with kefir? KSU J. Agric. Nat. 2021. [Google Scholar] [CrossRef]
- Anonymous. The Analysis of Agricultural Material; Reference Book; HMSO: London, UK, 1986; pp. 427–428. [Google Scholar]
- Playne, M.J.; Mc Donald, P. The buffering constituent of herbage and of silage. J. Sci. Food Agric. 1966, 17, 264–268. [Google Scholar] [CrossRef]
- Ulger, I.; Buyukkılıc Beyzi, S.; Kaliber, M.; Konca, Y. Chemical, nutritive, fermentation profile and gas production of citrus pulp silages, alone or combined with maize silage. S Afr. J. Anim. Sci. 2020, 50, 161–169. [Google Scholar] [CrossRef]
- Koc, F.; Coskuntuna, L. The comparison of the two different methods on the determination of organic acids in silage fodders. J. Anim. Prod. 2003, 44, 37–47. [Google Scholar]
- Seale, D.R.; Pahlow, G.; Spoelstra, S.F.; Lindgren, S.; Dellaglio, F.; Lowe, J.F. Methods for the Microbiological Analysis of Silage. In Proceedings of the Eurobac Conference, Uppsala, Sweden, 12–16 August 1986; pp. 147–164. [Google Scholar]
- Ashbell, G.; Weinberg, Z.G.; Azrieli, A.; Hen, Y.; Horev, B. A simple system to study the aerobic determination of silages. Can. Agric. Eng. 1991, 34, 171–175. [Google Scholar]
- Liu, B.; Huan, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef]
- Beasley, S.S.; Saris, P.E.J. Nisin-producing Lactococcus lactis strains from human milk. Appl. Environ. Microbiol. 2004, 70, 5051–5053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SAS. SAS/STAT® User’s Guide: Statistics (Version 9.1); SAS Institute Inc.: Cary, NC, USA, 2004. [Google Scholar]
- Li, P.; Ji, S.; Hou, C.; Tang, H.; Wang, Q.; Shen, Y. Effects of chemical additives on the fermentation quality and N distribution of alfalfa silage in south of China. Anim. Sci. J. 2016, 87, 1472–1479. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.H.; Yang, F.Y.; Zhang, J.G.; Shao, T. Characteristics of Lactobacillus parafarraginis ZH1 and its role in improving the aerobic stability of silages. J. Appl. Microbiol. 2014, 117, 405–416. [Google Scholar] [CrossRef]
- Lynch, J.P.; Jin, L.; Lara, E.C.; Baah, J.; Beauchemin, K.A. The effect of exogenous fibrolytic enzymes and a ferulic acid esterase-producing inoculant on the fibre degradability, chemical composition and conservation characteristics of alfalfa silage. Anim. Feed. Sci. Technol. 2014, 193, 21–31. [Google Scholar] [CrossRef]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L., Jr. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Wang, B.; Yu, Z. Effects of moisture content and additives on the ensiling quality and vitamins changes of alfalfa silage with or without rain damage. Anim. Sci. J. 2020, 91, e13379. [Google Scholar] [CrossRef]
- Wang, C.; He, L.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Fermentation quality and microbial community of alfalfa and stylo silage mixed with Moringa oleifera leaves. Bioresour. Technol. 2019, 284, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhao, M.; Wang, X.; Yu, Z.; Na, R. Ensiling alfalfa with whole crop corn improves the silage quality and in vitro digestibility of the silage mixtures. Grassl. Sci. 2017, 63, 211–217. [Google Scholar] [CrossRef]
- Ni, K.; Zhao, J.; Zhu, B.; Su, R.; Pan, Y.; Ma, J.; Zhou, G.; Tao, Y.; Liu, X.; Zhong, J. Assessing the fermentation quality and microbial community of the mixed silage of forage soybean with crop corn or sorghum. Bioresour. Technol. 2018, 265, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Ávila, C.L.S.; Carvalho, B.F.; Pinto, J.C.; Duarte, W.F.; Schwan, R.F. The use of Lactobacillus species as starter cultures for enhancing the quality of sugar cane silage. J. Dairy Sci. 2014, 97, 940–951. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Xu, D.; Xie, D.; Wang, M.; Li, Z.; Guo, X. Effects of antibacterial peptide-producing Bacillus subtilis and Lactobacillus buchneri on fermentation, aerobic stability, and microbial community of alfalfa silage. Bioresour. Technol. 2020, 315, 123881. [Google Scholar] [CrossRef] [PubMed]
- Kleinschmit, D.H.; Kung, L., Jr. The effects of Lactobacillus buchneri 40788 and Pediococcus pentosaceus R1094 on the fermentation of corn silage. J. Dairy Sci. 2006, 89, 3999–4004. [Google Scholar] [CrossRef] [Green Version]
- Borreani, G.; Tabacco, E.; Schmidt, R.J.; Holmes, B.J.; Muck, R.E. Silage review: Factors affecting dry matter and quality losses in silages. J. Dairy Sci. 2018, 101, 3952–3979. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Yao, D.; Li, D.; Lin, Y.; Bureenok, S.; Ni, K.; Yang, F. Effects of lactic acid bacteria isolated from rumen fluid and feces of dairy cows on fermentation quality, microbial community, and in vitro digestibility of alfalfa silage. Front. Microbiol. 2020, 10, 2998. [Google Scholar] [CrossRef] [PubMed]
- Auerbach, H.; Nadeau, E. Effects of additive type on fermentation and aerobic stability and its interaction with air exposure on silage nutritive value. Agronomy 2020, 10, 1229. [Google Scholar] [CrossRef]
- Schmidt, R.J.; Kung, L., Jr. The effects of Lactobacillus buchneri with or without a homolactic bacterium on the fermentation and aerobic stability of corn silages made at different locations. J. Dairy Sci. 2010, 93, 1616–1624. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.S.; Undersander, D.J.; Combs, D.K. Effect of Lactobacillus inoculants and forage dry matter on the fermentation and aerobic stability of ensiled mixed-crop tall fescue and meadow fescue. J. Dairy Sci. 2013, 96, 1735–1744. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Nishino, N. Identification and isolation of Lactobacillus fructivorans from wilted alfalfa silage with and without molasses. J. Appl. Microbiol. 2015, 120, 543–551. [Google Scholar] [CrossRef]
- Bai, J.; Ding, Z.; Ke, W.; Xu, D.; Wang, M.; Huang, W.; Zhang, Y.; Liu, F.; Guo, X. Different lactic acid bacteria and their combinations regulated the fermentation process of ensiled alfalfa: Ensiling characteristics, dynamics of bacterial community and their functional shifts. Microb. Biotechnol. 2021, 14, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.M.; Ke, W.C.; Zhang, P.; Li, F.H.; Guo, X.S. Characteristics of Pediococcus pentosaceus Q6 isolated from Elymus nutans growing on the Tibetan Plateau and its application for silage preparation at low temperature. J. Appl. Microbiol. 2018, 126, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yuan, X.; Desta, S.T.; Dong, Z.; Mugabe, W.; Shao, T. Characterization of Enterococcus faecalis JF85 and Enterococcus faecium Y83 isolated from Tibetan yak (Bos grunniens) for ensiling Pennisetum sinese. Bioresour. Technol. 2018, 257, 76–83. [Google Scholar] [CrossRef] [PubMed]






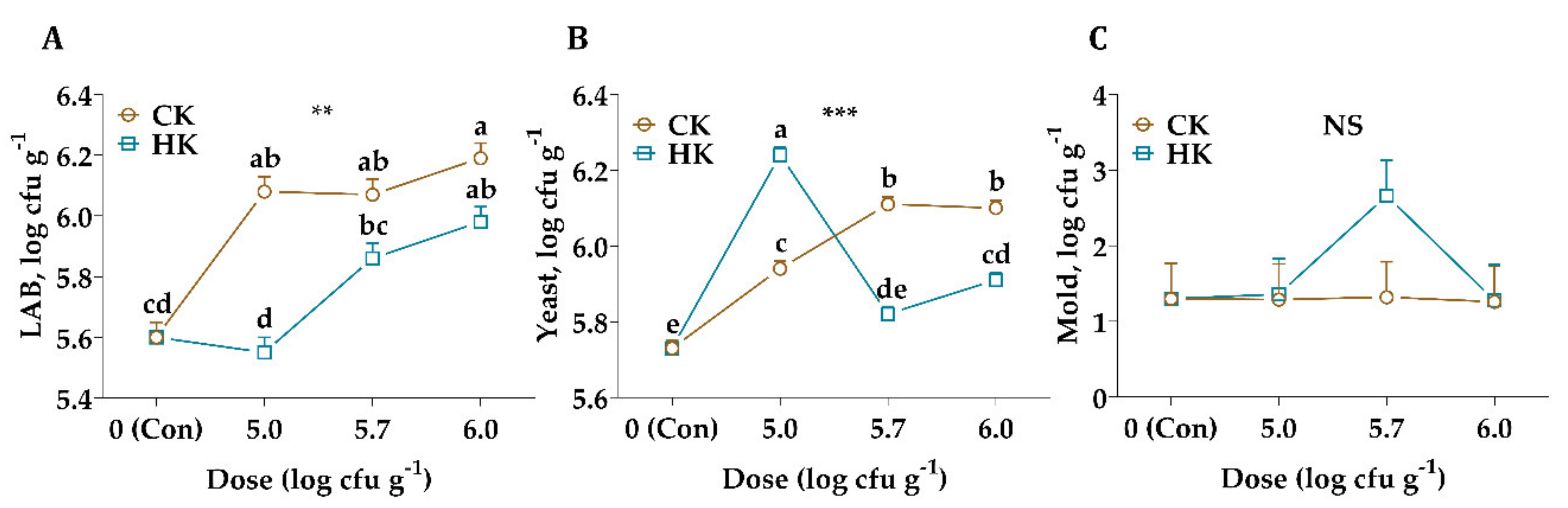{kind=link}
{kind=link}
{kind=link}
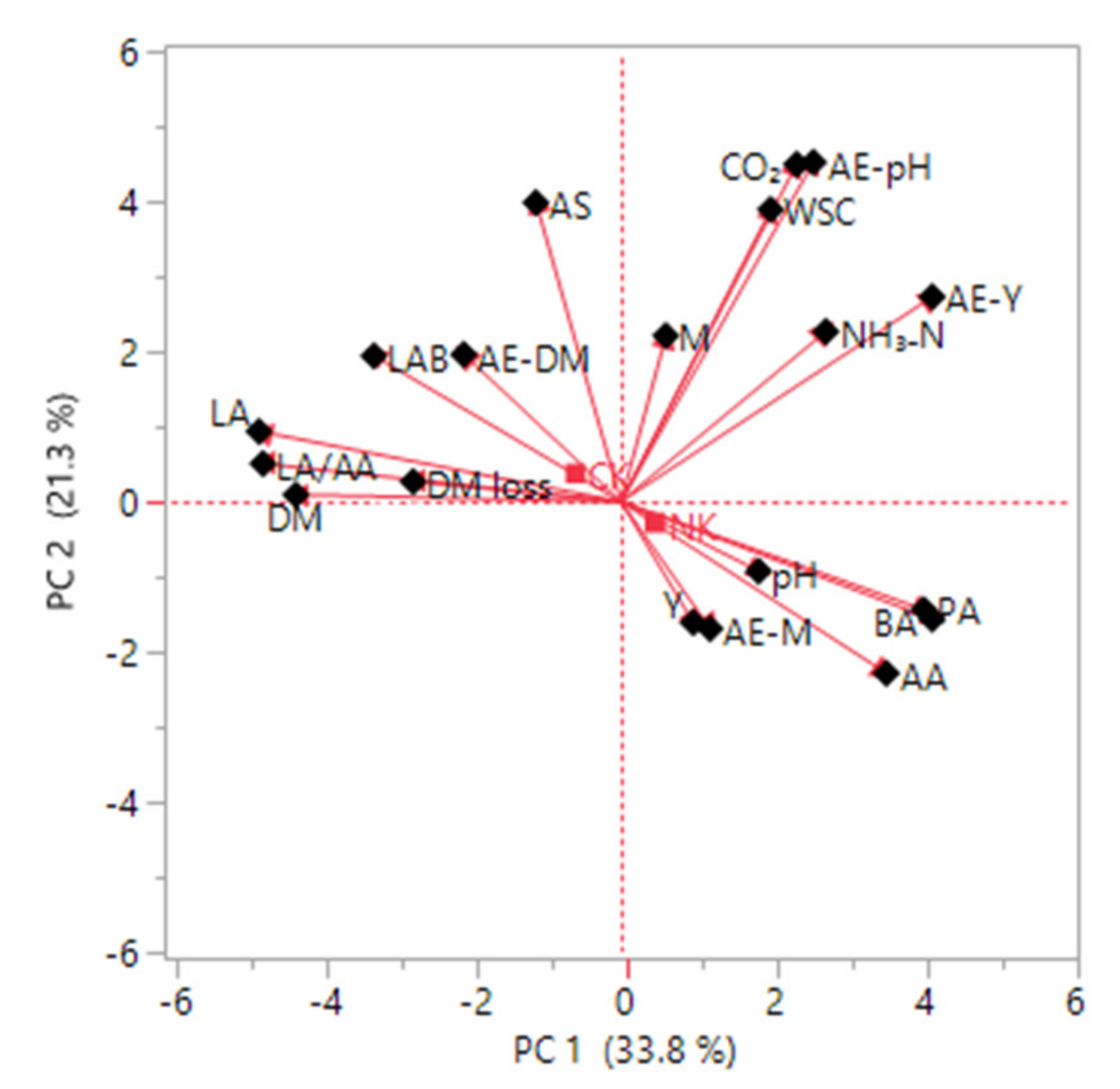{kind=link}
{kind=link}
| Item | CON (n = 10) | CK (Log cfu g−1) | HK (Log cfu g−1) | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|---|
| 5.0 (n = 10) | 5.7 (n = 10) | 6.0 (n = 10) | 5.0 (n = 10) | 5.7 (n = 10) | 6.0 (n = 10) | ||||
| DM | 284.5 | 305.2 | 292.3 | 299.4 | 272.5 | 279.5 | 315.4 | 9.54 | 0.151 |
| DM loss | 26.2 a | 26.2 a | 21.0c | 23.1 b | 20.9c | 25.6 a | 26.4 a | 0.21 | <0.001 |
| pH | 5.60 ab | 5.70 ab | 5.85 ab | 5.50 ab | 5.90 a | 5.45 b | 5.55 ab | 0.08 | 0.024 |
| WSC | 8.00 | 6.62 | 11.68 | 7.67 | 8.90 | 11.30 | 7.92 | 1.05 | 0.607 |
| Original Variable | PC 1 | PC 2 | PC 3 | PC 4 | PC 5 |
|---|---|---|---|---|---|
| Yeast (after aerobic exposure) | 0.780 * | 0.514 * | −0.050 | 0.231 | −0.030 |
| Butyric acid | 0.780 * | −0.294 | 0.023 | −0.180 | 0.276 |
| Propionic acid | 0.759 * | −0.271 | 0.120 | 0.003 | 0.382 |
| Acetic acid | 0.665 * | −0.430 | 0.429 | 0.272 | 0.077 |
| Ammonia nitrogen | 0.513 * | 0.427 | 0.446 | 0.049 | −0.547 * |
| pH | 0.345 | −0.174 | −0.626 * | 0.262 | −0.538 * |
| pH (after aerobic exposure) | 0.482 | 0.851 * | −0.016 | −0.053 | −0.007 |
| Carbon dioxide | 0.440 | 0.846 * | −0.156 | −0.091 | 0.051 |
| Aerobic stability | −0.213 | 0.750 * | 0.340 | 0.447 | −0.061 |
| Water soluble carbohydrate | 0.375 | 0.733 * | −0.288 | −0.067 | 0.240 |
| Mold | 0.113 | 0.416 | 0.010 | −0.630 * | 0.366 |
| Mold (after aerobic exposure) | 0.223 | −0.318 | 0.751 * | 0.410 | 0.212 |
| Dry matter loss | −0.521 * | 0.050 | 0.693 * | −0.423 | −0.145 |
| Lactic acid bacteria | −0.619 * | 0.366 | −0.088 | 0.507 * | 0.281 |
| Dry matter (after aerobic exposure) | −0.394 | 0.369 | 0.228 | 0.447 | 0.196 |
| Yeast | 0.181 | −0.303 | −0.826 * | 0.335 | 0.197 |
| Lactic acid/Acetic acid | −0.897 * | 0.095 | −0.151 | −0.218 | −0.037 |
| Lactic acid | −0.907 * | 0.176 | −0.206 | −0.068 | 0.077 |
| Dry matter | −0.815 * | 0.018 | 0.068 | 0.272 | 0.124 |
| Eigenvalue | 6.428 | 4.038 | 2.870 | 1.885 | 1.261 |
| Proportion (%) | 33.83 | 21.26 | 15.11 | 9.92 | 6.64 |
| Cumulative (%) | 33.83 | 55.09 | 70.19 | 80.11 | 86.75 |
| Treatments | Silo Opening | After 7 d of Aerobic Exposure |
|---|---|---|
| CON | Lactobacillus brevis | Enterococcus gallinarum, Enterococcus casseliflavus, Weissella paramesenteroides |
| CK1 | Pediococcus pentosaceus, Enterococcus faecium | Lactobacillus plantarum, Enterococcus faecalis |
| CK2 | Pediococcus pentosaceus, Enterococcus faecium | Bacillus sp., Enterococcus faecalis |
| CK3 | Enterococcus faecium | Lactobacillus plantarum, Lactobacillus brevis |
| HK1 | Lactobacillus brevis, Enterococcus faecium | Lactobacillus plantarum, Lactobacillus brevis |
| HK2 | Pediococcus pentosaceus, Enterococcus faecium | Weissella paramesenteroides, Bacillus sp. |
| HK3 | Lactobacillus brevis | Lactobacillus plantarum, Lactobacillus brevis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koç, F.; Özkan Ünal, E.; Okuyucu, B.; Esen, S.; Işık, R. Effect of Different Kefir Source on Fermentation, Aerobic Stability, and Microbial Community of Alfalfa Silage. Animals 2021, 11, 2096. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11072096
Koç F, Özkan Ünal E, Okuyucu B, Esen S, Işık R. Effect of Different Kefir Source on Fermentation, Aerobic Stability, and Microbial Community of Alfalfa Silage. Animals. 2021; 11(7):2096. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11072096
Chicago/Turabian StyleKoç, Fisun, Emel Özkan Ünal, Berrin Okuyucu, Selim Esen, and Raziye Işık. 2021. "Effect of Different Kefir Source on Fermentation, Aerobic Stability, and Microbial Community of Alfalfa Silage" Animals 11, no. 7: 2096. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11072096