
Sex Hormones in Hemolymph of Red King Crabs from the Barents Sea
by
 , , , and
, , , and
Alexander G. Dvoretsky
1,* ,
,
Elena V. Tipisova
2,
Aleksandra E. Elfimova
2 ,
,
Viktoria A. Alikina
2 and
Vladimir G. Dvoretsky
1 1
Murmansk Marine Biological Institute (MMBI), 183010 Murmansk, Russia
2
N. Laverov Federal Center for Integrated Arctic Research of the Ural Branch of the Russian Academy of Sciences (FECIAR UrB RAS), 163000 Arkhangelsk, Russia
*
Author to whom correspondence should be addressed.
Animals 2021, 11(7), 2149; https://0-doi-org.brum.beds.ac.uk/10.3390/ani11072149
Submission received: 25 June 2021
/
Revised: 17 July 2021
/
Accepted: 18 July 2021
/
Published: 20 July 2021
(This article belongs to the Special Issue Aquatic Animals: New Horizons for Blue Growth)
Abstract
:Simple Summary
Well-known sex hormones, testosterone and 17β-estradiol, play a crucial role in the reproduction of vertebrates. Biochemical assays have detected these substances in a few crustaceans, and it has been hypothesized that these hormones are involved in the regulation of crustacean reproduction. Red king crab is a large commercially important species harvested both in their native areas (North Pacific) and in the area of its introduction (Barents Sea). The presence of 17β-estradiol and testosterone and fluctuations of their concentrations in relation to different factors have not yet been investigated. For this reason, we provided a pilot study to reveal the levels of sex hormones in hemolymph of red king crabs captured in the coastal Barents Sea. These hormones were detected in the crabs and we compared our data with previously published data involving a wide range of crustaceans. We found seasonal variations in the level of testosterone with the maximum in the spawning period. Our data expand the current knowledge about the red king crab physiology and may be used for the development of its aquaculture.
Abstract
The presence of vertebrate-related steroid sex hormones has been reported in both freshwater and marine crustaceans. However, despite the commercial importance of king crabs, many aspects of their endocrinology are still unknown. For this reason, we examined hemolymph samples of the red king crab Paralithodes camtschaticus from the Barents Sea population for the presence of testosterone and 17β-estradiol using radioimmunoassay. The mean testosterone concentration was 0.46 ± 0.04 (range 0.08–1.39) ng mL–1, whereas the mean 17β-estradiol concentration was 1248.9 ± 91.4 (range 217.7–4100.1) pg mL–1. In general, the levels of 17β-estradiol and testosterone in red king crabs were higher than reported for the hemolymph of amphipods, crabs, and shrimps from warm and temperate waters, probably because the king crabs analyzed were larger and heavier than the other crustaceans. The concentrations of sex steroids did not differ significantly between males and females and between immature and mature red king crabs. Seasonal variations in the level of testosterone with the maximum value in the spawning period (May) indicate a potential role of the sex hormones in the maturation and reproduction processes of red king crab. Taking into account the slow growth rate in P. camtschaticus, our data could be useful not only for further physiological studies but also for the development of reliable techniques for red king crab aquaculture.
1. Introduction
The red king crab, Paralithodes camtschaticus (Tilesius, 1815), is a large commercially important species native to the North Pacific. A new self-sustaining population of this crab was reported to be established in the Barents Sea in the mid-1990s after its successful introduction by Russian specialists in the 1960s. Exponential population growth and range expansion of the crab has led to an increase in P. camtschaticus stock indices. Commercial fisheries for the red king crab were opened in 2002 in Norway and 2004 in Russia [1,2]. In Russian waters of the Barents Sea, annual landings of red king crab accounted for 9187 t in 2018, 9836 t in 2019 and 10,820 t in 2020 [3,4].
In the Barents Sea, different aspects of the red king crab biology and ecology have been intensively studied by marine scientists due to the invasive status of P. camtschaticus and its economic importance. Russian researchers have focused on distribution patterns, migration activity, molting and growth, reproduction, symbiotic relationships and effects on local benthic communities [2,4,5,6,7,8,9,10]. However, our knowledge of the physiology of this introduced species in its new environment is scarce, and information on biochemical profiles of red king crab is needed. Recently, we have reported concentrations of circulation ecdysteroids in hemolymph of P. camtschaticus [11]. Concentrations of a respiratory pigment, hemocyanin, were studied as a function of the meat content of commercial red king crabs from the Barents Sea [12].
The use of sex hormones and other hormones to regulate biological processes is a common strategy of vertebrate animals [13,14,15,16,17]. Usually, alterations in steroid hormone metabolism relate with effects on steroid hormone-dependent processes such as reproduction and growth [18]. Such alterations may be used as a biomarker for the effects of endocrine disruptors in invertebrates. Most stages of the steroidogenic pathways discovered for vertebrates have been demonstrated to occur in invertebrates. Although it was proposed that crustaceans do not actively synthesize vertebrate-type steroids in the same ways that echinoderms and mollusks do [19], the metabolism of these steroids was only investigated in a few crustaceans [18,20]. In crabs and lobsters, the endogenous precursor androstenedione is reduced to testosterone [21], while estrone is converted to estradiol [20]. Sex steroid-like substances can also be synthesized from cholesterol [22].
A high similarity was observed among the endocrine system of different crustaceans [23,24] and, therefore, sex hormones similar to those in vertebrates have been identified in several crustacean species including amphipods, shrimps, crayfish, lobsters, and crabs [19,25,26,27,28]. The presence of these hormones and their concentrations in red king crabs both in their native and non-native areas have not yet been reported. For this reason, the aim of our study was to measure concentrations of vertebrate-type sex hormones in hemolymph of the king crabs P. camtschaticus from the Barents Sea in relation to their sex and age.
2. Materials and Methods
Red king crabs were collected in August 2008 and May, July, and October 2009 in the coastal zone of the Barents Sea (Figure 1). The crabs were collected by divers at 5–40 m depths and using bottom traps at 50–60 m depths. Sex of each crab was determined visually. The crabs were weighed and their sizes were measured across the length of the carapace (CL, the greatest straight-line distance from the posterior margin of the right eye orbit to the medial-posterior margin of the carapace) [29]. All the crabs used for biochemical analyzes had new or old shells (2–24 months post ecdysis) [29]. Male crabs with CL < 90 mm were considered “immature,” whereas males with the CL > 90 mm were classified as “mature” according to Dvoretsky and Dvoretsky [8]. This separation is supported by the results of histological studies: in our samples the crabs smaller than 90 mm CL had no ripe spermatozoa in their testes in contrast to larger specimens. The same female groups were separated according to both histological data (immature females had oocyte diameter < 280 μm and white ovaries [5]) and size-at-maturity data (absence or presence of external eggs on their pleopods) [9]. In total, 23 immature (7 males and 16 females) and 33 mature (26 males and 7 females) crabs were analyzed.
After measuring the crabs, we sampled their hemolymph (0.6 mL) from the articulating membrane between the dactyl and propodus of the third walking leg using a sterile syringe with a needle, both washed with an anticoagulant containing EDTA and sodium citrate to prevent coagulation of the samples [29]. Each hemolymph sample was transferred to a sterile 2-mL plastic tube containing 1.4 mL of 96% ethanol. The samples were frozen and then transferred to the laboratory of the Institute of Physiology of Natural Adaptations (Arkhangelsk, Russia) for biochemical analyses.
Radioimmunoassays were carried out according to the protocols suggested by the manufacturer (A Beckman Coulter Company, Villepinte, France) using Immunotech RIA Testosterone or Immunotech RIA Estradiol test kits. In the case of testosterone, a 50-μL hemolymph sample and 500 μL of 125I-labelled testosterone were added serially to an antibody-coated tube. In the case of 17β-estradiol, a 100-μL hemolymph sample was used in the assays. A tube containing 500 μL of 125I-labelled testosterone (or estradiol) was included in every assay to determine the total 125I-activity (the sample “T”). The “testosterone” tubes were incubated at 37 °C in a waterbath for 3 h while the “estradiol” tubes were incubated with constant shaking (350 rpm) at ambient temperature for 3 h. At the end of incubation, the radioactivity of samples was determined using a scintillation counter (ARIAN, producer OOO VITAKO, Moscow, Russia). The standard graphs were plotted and the concentrations of testosterone and 17β-estradiol were obtained from the standard plot. The results were multiplied by a dilution factor of 3.33. The analytical sensitivity of the testosterone kit was 0.025 ng mL–1, the working RIA range was 0.025−20 ng mL–1, the coefficient of variation between samples was 14.8%. For the estradiol kit, these levels were <6 pg mL–1, 6−5000 pg mL–1 and 12.1%, respectively.
Carapace lengths and weight data in male and female red king crabs were compared using a one-way analysis of variance (ANOVA) or non-parametric Kruskal–Wallis tests in the case of non-normal data distribution. Since the sex hormone data were not normally distributed and failed testing for homogeneity of variances (modified Levene’s test, p < 0.05), testosterone and 17β-estradiol concentrations in relation to crab sex, crab size, and sampling season were compared using the Kruskal–Wallis test. When the test was significant, medians for different groups were compared using Bonferroni tests and differences were considered significant if p < 0.05. Statistical analyzes were carried out using STATISTICA (data analysis software system; http://www.statsoft.com/, accessed on 10 May 2020), version 6. Data are presented as means ± SE (standard error).
The taxonomic nomenclature follows the nomenclature according to WoRMS [30].
3. Results
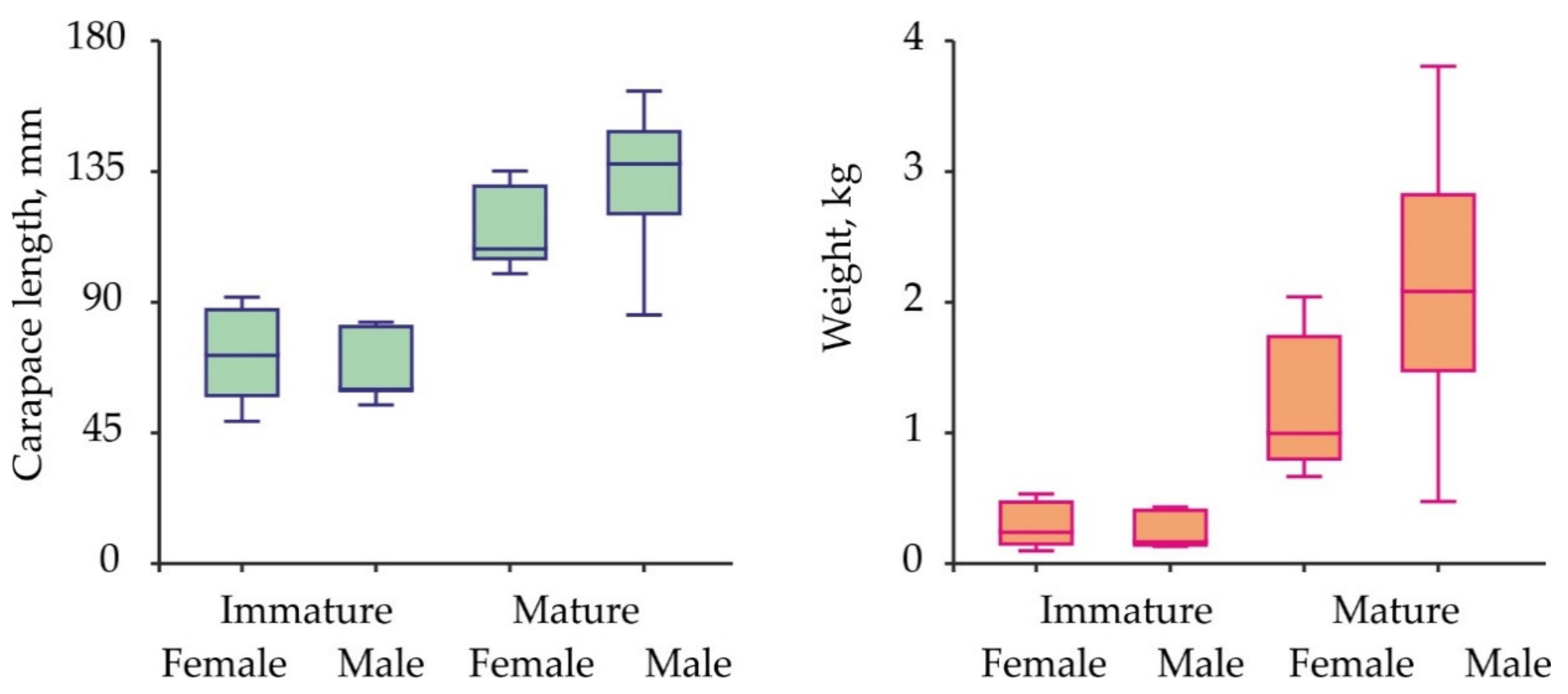
Size and weight variations in red king crabs collected for the hemolymph sampling are presented in Figure 2.
Carapace length ranged from 49.0 to 162.7 mm, body weight—from 97 to 3805 g. Mean carapace length in immature male and female crabs was similar (Kruskal-Wallis test, df = 1, H = 0.090, p = 0.764). The same result was obtained for mature specimens (Kruskal-Wallis test, df = 1, H = 3.421, p = 0.064). Mean weight did not differ significantly between immature males and females (ANOVA, df = 1, F = 0.600, p = 0.447). Mature males were heavier than mature females (ANOVA, df = 1, F = 4.645, p = 0.039).
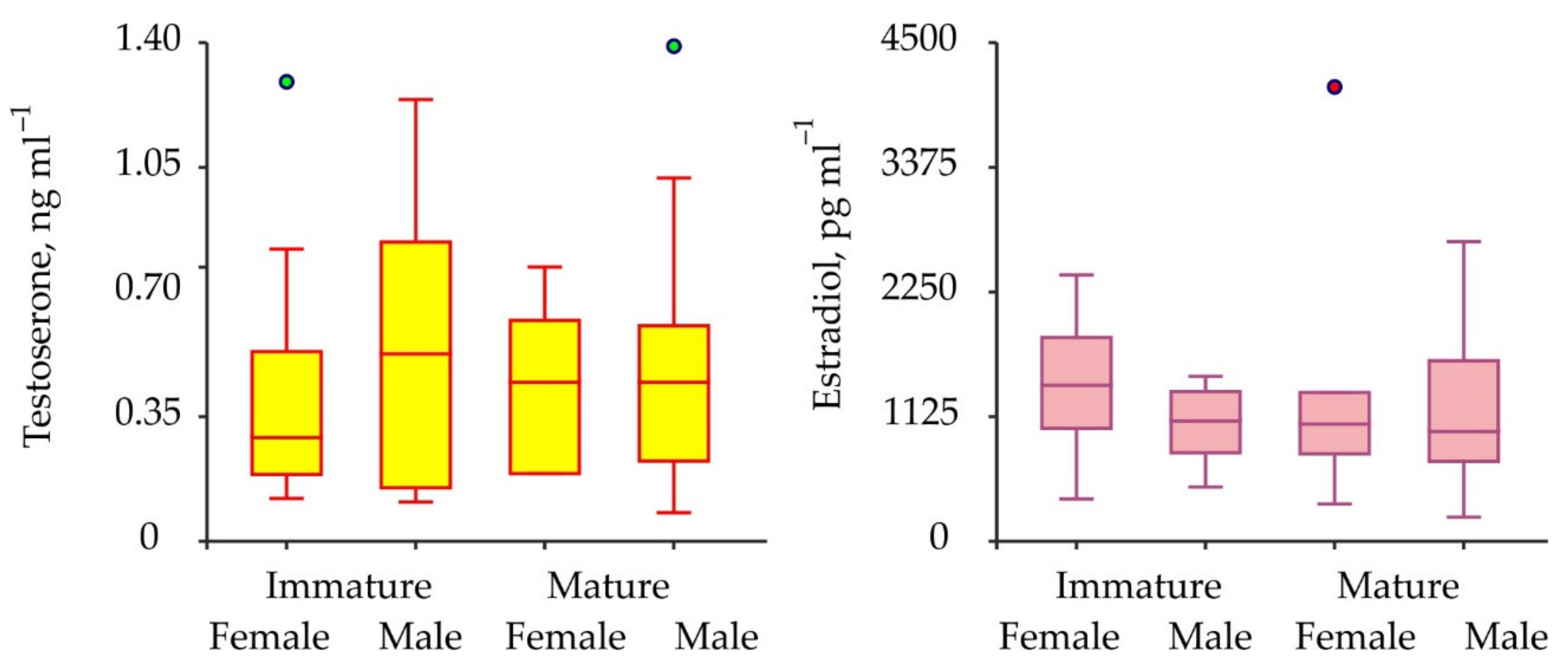
Biochemical assays have shown the presence of both testosterone and 17β-estradiol in the hemolymph of the crabs analyzed. Concentrations of testosterone varied from 0.08 to 1.39 ng mL–1 (mean for combined male and female data 0.46 ± 0.04 ng mL–1), while concentrations of 17β-estradiol ranged from 217.7–4100.1 pg mL–1 (1248.9 ± 91.4 pg mL–1) (Figure 3).
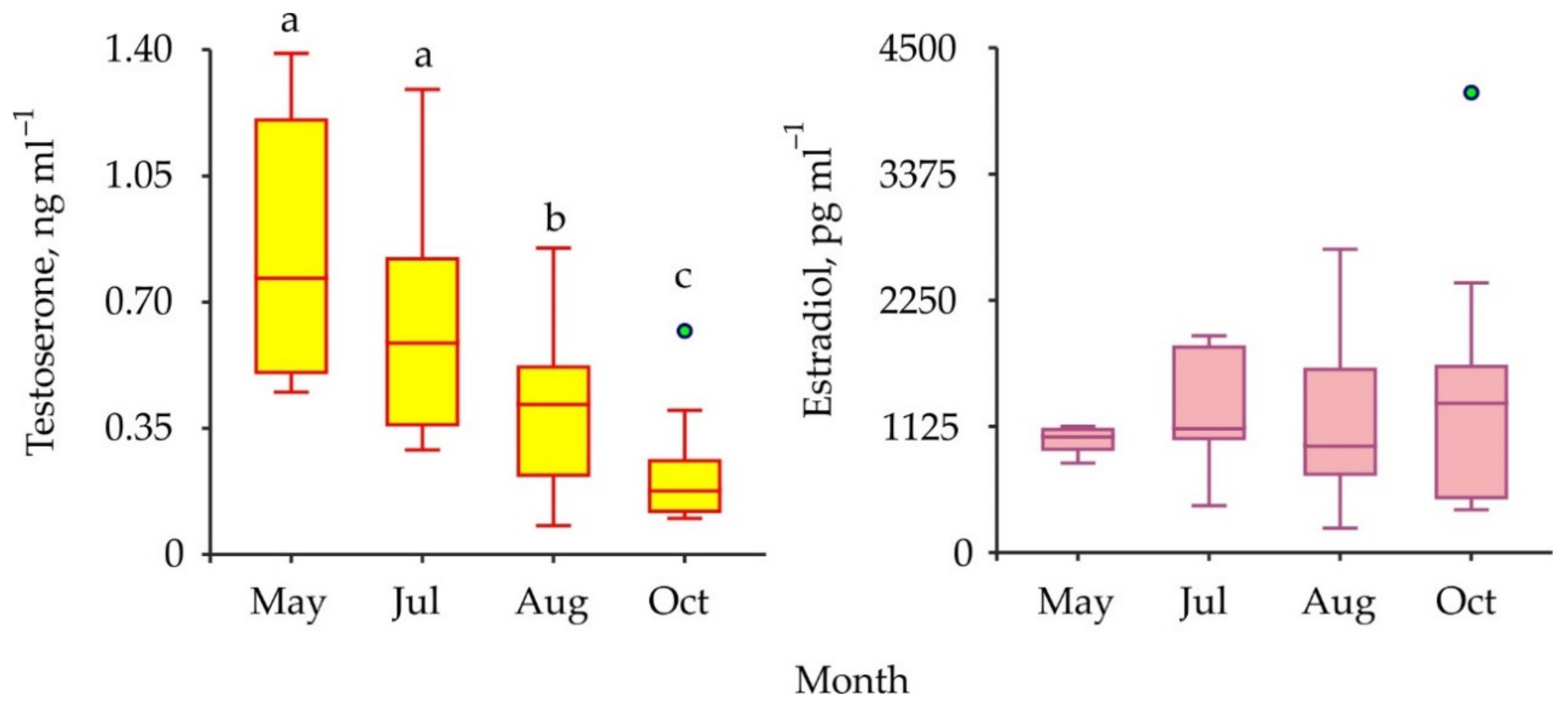
The comparisons of sex steroids in male and female red king crabs as well as in immature and mature specimens have shown that concentrations of both testosterone and 17β-estradiol were similar between all the combinations of the groups compared (Table 1). To increase the power of further analysis we used pooled data from males and females at different maturational stages in one complex. Seasonal variations in testosterone and 17β-estradiol concentrations are presented in Figure 4.
The mean level of testosterone decreased significantly from May to October whereas there was no significant difference detected among the seasonal concentrations of 17β-estradiol in the hemolymph samples (Table 1).
4. Discussion
According to our published [8] and unpublished size-at-age data, the age of the red king crabs used in this study varied from 3 to 11 years (females) and 3 to 13 years (males). The mean weight of large males was higher than that calculated for females because mature male red king crabs invest more energy in somatic growth in comparison to females, which invest the major proportion of their resources into reproduction processes [7,10].
Our study indicated that red king crabs have detectable concentrations of 17β-estradiol and testosterone in both male and female hemolymph. In general, the level of 17β-estradiol found for the female red king crabs was higher (Table 2) in comparison to females of some other decapod species such as Norwegian lobster Nephrops norvegicus in western Scotland [31], mud crabs Scylla serrata in India [32] and Scylla paramamosain in China [27], Chinese mitten crab Eriocheir sinensis in China [33], edible crab Spiralothelphusa senex (cited as Oziothelphusa senex senex) in India [34], soldier crab Mictyris brevidactylus in northern Taiwan [35] and grass shrimp Pandalus kessleri in Japan [25].
The estradiol concentration registered in the red king crab male hemolymph was higher than in Eriocheir sinensis males [33], while the male testosterone level was comparable to the concentration reported for the hemolymph of S. paramamosain [27]. In the red king crabs, the levels of both hormones were much higher than in gammarid amphipods Gammarus duebeni celticus and G. pulex in Northern Ireland and G. pseudolimnaeus in Waukesha County, WI, USA [28]. It is difficult to evaluate causes responsible for this result, but most likely the higher concentrations of sex hormones in the red king crab hemolymph were associated with their larger size and weight. For example, the mud crabs S. paramamosain used in the study described by Huiyang et al. [27] had a body length of 36–87 mm, while the average weight of mature Spiralothelphusa senex females used for biochemical assays in the study by Swetha et al. [34] was 32–37 g. Additionally, the largest CL reported for adult female Nephrops norvegicus is about 50 mm CL [41]; females of Scylla serrata attain sexual maturity after reaching 80 mm carapace width (CW) [42], and females of Eriocheir sinensis become mature at 34.1 ± 3.9 mm CL with a weight of 26.6 ± 8.1 g [43]. The largest adult females of Mictyris brevidactylus belong to a size class of 110–120 mm CW [44]. The majority of the mentioned crustaceans occur in temperate and warm waters whereas the red king crabs live in a cold-water environment. Different habitats may also contribute to differences between the levels of sex hormones in P. camtschaticus and those crustacean species.
In vertebrates, both males and females produce testosterone and 17β-estradiol, although the concentrations of these hormones may vary significantly depending on sex and the reproductive cycle [28].
Taking into account the differences in invertebrate and vertebrate endocrine systems, there was no reason to assume that typical sex-specific differences in sex hormones among vertebrates would also be found in red king crabs. Our data support to some extent this assumption: we found no significant differences in testosterone and 17β-estradiol concentrations in relation to age and sex of the red king crabs. Similar results were reported by other authors. For example, males of Gammarus duebeni celticus, G. pulex, and G. pseudolimnaeus had comparable concentrations of 17β-estradiol, and females of these species had comparable concentrations of testosterone [28].
On the other hand, the ovaries of some decapods may synthesize 17β-estradiol and release this hormone into the hemolymph, and then it may reach the hepatopancreas to stimulate vitellogenin synthesis [26]. For this reason, concentrations of sex steroids in gonads, eggs, and hepatopancreas of crustaceans are higher than in their hemolymph (Table 2). Thus, an association between sex hormone profiles and stages of vitellogenesis was detected in some decapod species [45]. For example, the levels of 17β-estradiol increased significantly in the tissues of crabs Scylla serrata, S. paramamosain, and Spiralothelphusa senex during late vitellogenesis [27,32,34]. Similar trends were reported for testosterone as well [27]. In addition, concentrations of sex hormones were found to positively correlate with the reproductive status of decapod crustaceans, as they were high during the pre-reproductive season and low during the non-reproductive season [35,37,46]. We also found seasonal changes in the testosterone profile assayed in the Barents Sea red king crabs. The maximum (0.8 ng mL–1) was registered in May and the minimum (0.2 ng mL–1) in October. Spawning season in P. camtschaticus lasts from February to May with a peak in April [5]. Thus, we expected to find the highest level of testosterone in the May samples of adult crabs. We found no seasonal fluctuations in the 17β-estradiol concentration. This result could be explained by the fact that we studied hemolymph, whereas estrogens are synthesized in the ovaries of red king crabs. These organs are much heavier than the male testes. Monthly variations of testosterone in immature crabs more likely reflected seasonal patterns of P. camtschaticus maturation in the Barents Sea.
The effects of 17β-estradiol on vitellogenesis (vitellogenin synthesis, oocyte development, an increase of ovarian indices, and growth rates) have been shown for the kuruma prawn Marsupenaeus japonicus [47], giant freshwater prawn Macrobrachium rosenbergii [48], freshwater crayfish Cherax albidus [49], burrowing crab Chasmagnathus grdnulata [50], and lobsters Homarus americanus and Panulirus homarus [51]. In addition, Nagabhushanam and Kulkarni [52] showed that exogenous testosterone can stimulate testicular development in the marine penaeid prawn, Mierspenaeopsis hardwickii (cited as Parapenaeopsis hardwickii). This hormone also induced sperm development and subsequent mating success in male tiger shrimps Penaeus monodon [53]. In contrast, the results by Okumura and Sakiyama [38] suggested that 17β-estradiol and testosterone did not play an important role in ovarian development in Penaeus japonicus (cited as Marsupenaeus japonicus), and Koskela et al. [54] reported no effect 17β-estradiol on development and reproduction of the tiger prawn Penaeus esculentus.
In the last several decades, aquaculture has grown rapidly in complexity and in volume around the world. The majority of aquaculture businesses require substantial amounts of both operating and investment capital. For this reason, profitable aquaculture cannot be organized without excellent knowledge on the biology and ecology of target species [55,56,57]. The development of specific methods to increase growth rates of slow-growing species such as red king crab is a great challenge for modern science [58,59], and our results may have important implications for further studies focused on aquaculture of P. camtschaticus.
5. Conclusions
Because molting and reproduction in crustaceans are controlled both by ecdysteroids (molting hormones) and vertebrate-like steroids, these hormones are considered as potential inductors of growth and maturation processes in cultured crustaceans. Understanding the controlling mechanisms of reproductive processes and molting in red king crabs is an important issue because of the slow growth rate of this species (the commercial size of 150 mm CW is reached at 10 years). Thus, the determination of the sex steroid levels in the red king crab Paralithodes camtschaticus is a first step toward developing new aquaculture techniques. The level of testosterone/17β-estradiol varied from 0.08–1.39 ng mL–1/217.7–2703.2 pg mL–1 in males and from 0.12–1.29 ng mL–1/335.6–4100.1 pg mL–1 in females. Further studies should reveal the possibility of the use of artificially synthesized hormonal preparations for accelerating the maturation process in red king crabs and, consequently, increasing their growth rates under laboratory conditions. Such studies will provide a baseline for the effective planning of regulatory measures and aquaculture programs in the Barents Sea region.
Author Contributions
Conceptualization, A.G.D. and E.V.T.; data curation, A.G.D., A.E.E., V.A.A., and V.G.D.; validation, formal analysis, A.G.D., E.V.T., V.G.D., A.E.E. and V.A.A., funding acquisition, A.G.D. and E.V.T.; methodology, E.V.T.; software, visualization, V.G.D., project administration, A.G.D.; writing—original draft, A.G.D. and V.G.D.; writing—review and editing, A.G.D., V.G.D. and E.V.T. All authors have read and agreed to the published version of the manuscript.
Funding
This article is based upon work that was supported by the Ministry of Science and Higher Education of the Russian Federation under projects AAAA-A19-119011690138-0 for MMBI RAS and AAAA-A19-119120990060-0 for FECIAR UrB RAS.
Institutional Review Board Statement
This study was carried out in accordance with Russian legislation guidelines (Federal Act 52-F3, 24/04/1995) and approved by the Institutional Review of Murmansk Marine Biological Institute (6732/253-ok, 01/12/2016).
Data Availability Statement
Data will be made available upon any reasonable request to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dvoretsky, A.G.; Dvoretsky, V.G. Commercial fish and shellfish in the Barents Sea: Have introduced crab species affected the population trajectories of commercial fish? Rev. Fish Biol. Fish. 2015, 25, 297–322. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Red king crab (Paralithodes camtschaticus) fisheries in Russian waters: Historical review and present status. Rev. Fish Biol. Fish. 2018, 28, 331–353. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acid composition of the Barents Sea red king crab (Paralithodes camtschaticus) leg meat. J. Food Compos. Anal. 2021, 98, 103826. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. New echinoderm-crab epibiotic associations from the coastal Barents Sea. Animals 2021, 11, 917. [Google Scholar] [CrossRef]
- Kuzmin, S.A.; Gudimova, E.N. Introduction of the Kamchatka (Red King) Crab in the Barents Sea. Pecularities of Biology, Perspectives of Fishery; Publishing Kola Science Centre Russian Academy of Sciences: Apatity, Russia, 2002. (In Russian) [Google Scholar]
- Pavlova, L.V. Red king crab trophic relations and its influence on bottom biocenoses. In Biology and Physiology of the Red King Crab from the Coastal Zone of the Barents Sea; Matishov, G.G., Ed.; Publishing Kola Scientific Centre Russian Academy of Sciences: Apatity, Russia, 2008; pp. 77–104. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Population dynamics of the invasive lithodid crab, Paralithodes camtschaticus, in a typical bay of the Barents Sea. ICES J. Mar. Sci. 2013, 70, 1255–1262. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size-at-age of juvenile red king crab (Paralithodes camtschaticus) in the coastal Barents Sea. Cah. Biol. Mar. 2014, 55, 43–48. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size at maturity of female red king crab, Paralithodes camtschaticus, from the costal zone of Kola Peninsula (southern Barents Sea). Cah. Biol. Mar. 2015, 56, 49–54. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Effects of environmental factors on the abundance, biomass, and individual weight of juvenile red king crabs in the Barents Sea. Front. Mar. Sci. 2020, 7, 726. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Hemolymph molting hormone concentrations in red king crabs from the Barents Sea. Polar Biol. 2010, 33, 1293–1298. [Google Scholar] [CrossRef]
- Moiseeva, S.A.; Moiseev, S.I. Relation between muscular tissue condition in limbs and hemocyanin concentration in the hemolymph of red king crab (Paralithodes camtschaticus) in the Barents Sea. Probl. Fish. 2008, 9, 200–217, (In Russian with English Abstract). [Google Scholar]
- Kutzler, M.A. Possible relationship between long-term adverse health effects of gonad-removing surgical sterilization and luteinizing hormone in dogs. Animals 2020, 10, 599. [Google Scholar] [CrossRef] [Green Version]
- Am-in, N.; Suwimonteerabutr, J.; Kirkwood, R.N. Serum anti-mullerian hormone and estradiol concentrations in gilts and their age at puberty. Animals 2020, 10, 2189. [Google Scholar] [CrossRef] [PubMed]
- Hyndman, T.H.; Algar, K.L.; Woodward, A.P.; Coiacetto, F.; Hampton, J.O.; Nickels, D.; Hamilton, N.; Barnes, A.; Algar, D. Estradiol-17β pharmacokinetics and histological assessment of the ovaries and uterine horns following intramuscular administration of estradiol cypionate in feral cats. Animals 2020, 10, 1708. [Google Scholar] [CrossRef]
- Massoud, D.; Lao-Pérez, M.; Ortega, E.; Burgos, M.; Jiménez, R.; Barrionuevo, F.J. Divergent seasonal reproductive patterns in syntopic populations of two murine species in southern Spain, Mus spretus and Apodemus sylvaticus. Animals 2021, 11, 243. [Google Scholar] [CrossRef]
- García, M.-L.; Muelas, R.; Argente, M.-J.; Peiró, R. Relationship between prenatal characteristics and body condition and endocrine profile in rabbits. Animals 2021, 11, 95. [Google Scholar] [CrossRef]
- Verslycke, T.; De Wasch, K.; De Brabander, H.F.; Janssen, C.R. Testosterone metabolism in the estuarine mysid Neomysis integer (Crustacea; Mysidacea): Identification of testosterone metabolites and endogenous vertebrate-type steroids. Gen. Comp. Endocrinol. 2002, 126, 190–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janer, G.; Porte, C. Sex steroids and potential mechanisms of non-genomic endocrine disruption in invertebrates. Ecotoxicology 2007, 16, 145–160. [Google Scholar] [CrossRef]
- Swevers, L.; Lambert, J.G.; De Loof, A. Metabolism of vertebrate-type steroids by tissues of three crustacean species. Comp. Biochem. Physiol. 1991, 99B, 35–41. [Google Scholar] [CrossRef]
- Blanchet, M.-F.; Ozon, R.; Meusy, J.J. Metabolism of steroids, in vitro, in the male crab Carcinus maenas Linne. Comp. Biochem. Physiol. 1972, 41B, 251–261. [Google Scholar] [CrossRef]
- Shih, J.T.; Liao, C.F. Conversion of cholesterol to sex steroid-like substances by tissues of Mictyris brevidactylus in vitro. Zool. Stud. 1998, 37, 102–110. [Google Scholar]
- Charmantier, G.; Charmantier-Daures, M.; Van Herp, F. Hormonal regulation of growth and reproduction in crustaceans. In Recent Advances in Marine Biotechnology; Fingerman, M., Nagabhushanam, R., Eds.; Oxford & IBH: New Delhi, India, 1997; Volume 1, pp. 109–161. [Google Scholar]
- Swetha, C.H.; Sainath, S.B.; Reddy, P.R.; Reddy, P.S. Reproductive Endocrinology of Female Crustaceans: Perspective and Prospective. J. Mar. Sci. Res. Dev. 2011, 3, 1–13. [Google Scholar]
- Quinitio, E.T.; Yamauchi, K.; Hara, A.; Fuji, A. Profiles of progesterone-and estradiol-like substances in the hemolymph of female Pandalus kessleri during an annual reproductive cycle. Gen. Comp. Endocrinol. 1991, 81, 343–348. [Google Scholar] [CrossRef]
- Gunamalai, V.; Kirubagaran, R.; Subramoniam, T. Vertebrate steroids and the control of female reproduction in two decapods, Emertia asiatica and Macrobrachium rosenbergii. Curr. Sci. 2006, 90, 119–123. [Google Scholar]
- Huiyang, H.; Haihui, Y.; Shizhao, H.; Guizhong, W. Profiles of gonadotropins and steroid hormone-like substances in the hemolymph of mud crab Scylla paramamosain during the reproduction cycle. Mar. Freshw. Behav. Physiol. 2009, 42, 297–305. [Google Scholar] [CrossRef]
- Lewis, S.E.; Freund, J.G.; Riddell, G.E.; Wankowski, J.L.; Dick, J.T.A.; Baldridge, M.G. Interspecific comparison of estrogen and testosterone concentrations in three species of amphipods (Gammarus duebeni celticus, G. pseudolimnaeus, and G. pulex). J. Crustacean Biol. 2015, 35, 789–792. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, W.E.; Byersdorfer, S.E. Biological Field Techniques for Lithodid Crabs. Fairbanks, Alaska Sea Grant College Program; University of Alaska: Fairbanks, AK, USA, 2005. [Google Scholar]
- WoRMS. World Register of Marine Species. 2021. Available online: http://www.marinespecies.org/ (accessed on 22 February 2021).
- Fairs, N.J.; Evershed, R.P.; Quinlan, P.T.; Goad, L.J. Detection of unconjugated and conjugated steroids in the ovary, eggs, and haemolymph of the decapod crustacean Nephrops norvegicus. Gen. Comp. Endocrinol. 1989, 4, 199–208. [Google Scholar] [CrossRef]
- Warrier, S.R.; Tirumalai, R.; Subramoniam, T. Occurrence of vertebrate steroids, estradiol 17 β and progesterone in the reproducing females of the mud crab Scylla serrata. Comp. Biochem. Physiol. Part A 2001, 130, 283–294. [Google Scholar] [CrossRef]
- Wei, W.; Wei, H.; Liu, Q. Effect of estradiol in hemolymph and gonad on precociousness of Eriocheir sinensis. J. Fish. China 2005, 29, 826–865. Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-SCKX200506020.htm (accessed on 20 May 2020). (In Chinese with English Abstract).
- Swetha, C.H.; Girish, B.P.; Reddy, P.S. Elucidation of the role of estradiol and progesterone in regulating reproduction in the edible crab, Oziothelphusa senex senex. RSC Adv. 2016, 6, 24959. [Google Scholar] [CrossRef]
- Shih, J.-T. Sex steroid-like substances in the ovaries, hepatopancreases, and body fluid of female Mictyris brevidactylus. Zool. Stud. 1997, 36, 136–145. [Google Scholar]
- Amin-Safwan, A.; Muhd-Farouk, H.; Mardhiyyah, M.P.; Nadirah, M.; Ikhwanuddin, M. Does water salinity affect the level of 17β-estradiol and ovarian physiology of orange mud crab, Scylla olivacea (Herbst, 1796) in captivity? J. King Saud. Univ. Sci. 2019, 31, 827–835. [Google Scholar] [CrossRef]
- Quinitio, E.T.; Hara, A.; Yamauchi, K.; Nakao, S. Changes in the steroid hormone and vitellogenin levels during the gametogenic cycle of the giant tiger shrimp, Penaeus monodon. Comp. Biochem. Physiol. 1994, 109C, 21–26. [Google Scholar] [CrossRef]
- Okumura, T.; Sakiyama, K. Hemolymph levels of vertebrate-type steroid hormones in female kuruma prawn Marsupenaeus japonicus (Crustacea: Decapoda: Penaeidae) during natural reproductive cycle and induced ovarian development by eyestalk ablation. Fish. Sci. 2004, 70, 372–380. [Google Scholar] [CrossRef]
- Burns, B.G.; Sangalang, G.B.; Freeman, H.C.; McMenemy, M. Isolation and identification of testosterone from the serum and testes of the American lobster (Homarus americanus). Gen. Comp. Endocrinol. 1984, 54, 429–432. [Google Scholar] [CrossRef]
- Huang, D.-J.; Chen, H.-C. Effects of chlordane and lindane on testosterone and vitellogenin levels in green neon shrimp (Neocaridina denticulata). Int. J. Toxicol. 2004, 23, 91–95. [Google Scholar] [CrossRef]
- Tuck, I.D.; Atkinson, R.J.A.; Chapman, C.J. Population biology of the Norway lobster, Nephrops norvegicus (L.) in the Firth of Clyde, Scotland II: Fecundity and size at onset of sexual maturity. ICES J. Mar. Sci. 2000, 57, 1227–1239. [Google Scholar] [CrossRef]
- Prasad, P.N.; Neelakantan, B. Maturity and breeding of the mud crab, Scylla serrata (Forskal) (Decapoda: Brachyura: Portunidae). Proc. Indian Acad. Sci. Anim. Sci. 1989, 98, 341–349. [Google Scholar] [CrossRef]
- Tanglin, Z.; Zhongjie, L.; Yibo, C. Survival, growth, sex ratio, and maturity of the Chinese mitten crab (Eriocheir sinensis) reared in a Chinese pond. J. Freshw. Ecol. 2001, 16, 633–640. [Google Scholar]
- Takeda, S. Sexual differences in behaviour during the breeding seasonin the soldier crab (Mictyris brevidactylus). J. Zool. Lond. 2005, 266, 197–204. [Google Scholar] [CrossRef]
- Subramoniam, T. Steroidal control of vitellogenesis in Crustacea: A new understanding for improving shrimp hatchery production. Proc. Indian Acad. Sci. Anim. Sci. 2017, 83, 595–610. [Google Scholar]
- Pan, J.; Liu, M.; Chen, T.; Yongxu, C.; Xugan, W. Immunolocalization and changes of 17beta-estradiol during ovarian development of Chinese mitten crab Eriocheir Sinensis. Cell Tissue Res. 2018, 373, 509–520. [Google Scholar]
- Yano, I.; Hoshino, R. Effects of 17β-estradiol on the vitellogenin synthesis and oocyte development in the ovary of kuruma prawn (Marsupenaeus japonicus). Comp. Biochem. Physiol. Part A 2006, 144, 18–23. [Google Scholar] [CrossRef]
- Pakdeenarong, N. Effect of estradiol-17β on embryonic tolerance, growth, and muscular compactness of giant freshwater prawn, Macrobrachium rosenbergii. J. Appl. Biol. Biotechnol. 2019, 7, 16–20. [Google Scholar]
- Coccia, E.; De Lisa, E.; Di Cristo, C.; Di Cosmo, A.; Paolucci, M. Effects of estradiol and progesterone on the reproduction of the freshwater crayfish Cherax albidus. Biol. Bull. 2010, 218, 36–47. [Google Scholar] [CrossRef]
- Zapata, V.; Lopez Greco, L.S.; Medesani, D.; Rodriguez, E.M. Ovarian growth in the crab, Chasmagnathus granulata induced by hormones and neuroregulators throughout the year. In vivo and in vitro studies. Aquaculture 2003, 224, 1–4. [Google Scholar] [CrossRef]
- Subramoniam, T.; Kirubagaran, R. Endocrine regulation of vitellogenesis in lobsters. J. Mar. Biol. Assoc. India 2010, 52, 229–236. [Google Scholar]
- Nagabhushanam, R.; Kulkarni, G.K. Effect of exogenous testosterone on the androgenic gland and testis of a marine penaeid prawn, Parapenaeopsis hardwickii (Miers) (Crustacea, Decapoda, Penaeidae). Aquaculture 1981, 23, 19–27. [Google Scholar] [CrossRef]
- Maheswarudu, G.; Rajkumar, U.; Sreeram, M.P.; Chakravarty, M.S.; Sajeev, C.K. Effect of testosterone hormone on performance of male broodstock of black tiger shrimp Penaeus monodon Fabricius, 1798. J. Vet. Sci. Photon 2015, 116, 446–456. [Google Scholar]
- Koskela, R.W.; Greenwood, J.G.; Rothlisberg, P.C. The influence of prostaglandin E2 and the steroid hormones, 17α-hydroxyprogesterone and 17β-estradiol on moulting and ovarian development in the tiger prawn, Penaeus esculentus Haswell, 1879 (Crustacea: Decapoda). Comp. Biochem. Physiol. Part A 1992, 101, 295–299. [Google Scholar] [CrossRef]
- Fiordelmondo, E.; Magi, G.E.; Mariotti, F.; Bakiu, R.; Roncarati, A. Improvement of the water quality in rainbow trout farming by means of the feeding type and management over 10 years (2009–2019). Animals 2020, 10, 1541. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Aquaculture of green sea urchin in the Barents Sea: A brief review of Russian studies. Rev. Aquac. 2020, 12, 1280–1290. [Google Scholar] [CrossRef]
- Samat, N.A.; Yusoff, F.M.; Rasdi, N.W.; Karim, M. Enhancement of live food nutritional status with essential nutrients for improving aquatic animal health: A review. Animals 2020, 10, 2457. [Google Scholar] [CrossRef] [PubMed]
- Dvoretsky, A.G.; Dvoretsky, V.G. Does spine removal affect molting process in the king red crab (Paralithodes camtschaticus) in the Barents Sea? Aquaculture 2012, 326–329, 173–177. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Inter-annual dynamics of the Barents Sea red king crab (Paralithodes camtschaticus) stock indices in relation to environmental factors. Polar Sci. 2016, 10, 541–552. [Google Scholar] [CrossRef]
Figure 1.
Study area in the southern Barents Sea.

Figure 2.
Carapace length and weight variations in the Barents Sea red king crabs analyzed for sex hormones.
Figure 2.
Carapace length and weight variations in the Barents Sea red king crabs analyzed for sex hormones.

Figure 3.
Variations of sex hormones in immature and mature red king crabs from the coastal zone of the Barents Sea. Green and red circles are outliers.
Figure 3.
Variations of sex hormones in immature and mature red king crabs from the coastal zone of the Barents Sea. Green and red circles are outliers.

Figure 4.
Seasonal variations in vertebrate-type sex hormones in hemolymph of red king crabs from the Barents Sea. Green circles are outliers. Bars with the same letter are not significantly different (p > 0.05).
Figure 4.
Seasonal variations in vertebrate-type sex hormones in hemolymph of red king crabs from the Barents Sea. Green circles are outliers. Bars with the same letter are not significantly different (p > 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of Kruskal–Wallis tests comparing the differences in sex hormone levels of red king crabs.
Table 1.
Results of Kruskal–Wallis tests comparing the differences in sex hormone levels of red king crabs.
| Hormone | Comparisons | d | H | p |
|---|---|---|---|---|
| Testosterone | M-im vs. M-mat | 1 | 0.436 | 0.509 |
| F-im vs. F-mat | 1 | 0.875 | 0.349 | |
| M-tot vs. F-tot | 1 | 0.750 | 0.386 | |
| May vs. July vs. August vs. October | 3 | 24.030 | 0.00002 | |
| 17β-estradiol | M-im vs. M-mat | 1 | 0.001 | 0.982 |
| F-im vs. F-mat | 1 | 0.698 | 0.404 | |
| M-tot vs. F-tot | 1 | 1.032 | 0.310 | |
| May vs. July vs. August vs. October | 3 | 0.950 | 0.813 |
Note. M—male, F- female, im—immature, mat—mature, tot—total, d—degree of freedom, H—chi-square level, p—probability level.
Table 2.
Concentrations of vertebrate-like sex steroids identified in various crustacean species.
| Steroid | Species | Sex | Matrix | Levels | Reference |
|---|---|---|---|---|---|
| E | Nephrops norvegicus | F | Eggs | 900 pg g–1 | [31] |
| E | Nephrops norvegicus | F | Hemolymph | 800 pg mL–1 | [31] |
| E | Scylla serrata | F | Hemolymph | 230–1040 pg g–1 | [32] |
| E | Scylla serrata | F | Ovary | 180–1250 pg g–1 | [32] |
| E | Scylla serrata | F | Hepatopancreas | 300–4100 pg mg–1 | [32] |
| E | Scylla olivacea | F | Hemolymph | 970–3520 pg mL–1 | [36] |
| E | Pandalus latirostris | F | Hemolymph | 18.1–54.3 pg mL–1 | [25] |
| E | Penaeus monodon | F | Hemolymph | 30.4 pg mL–1 | [37] |
| E | Penaeus monodon | F | Ovary | 30–77 pg g–1 | [37] |
| E | Penaeus monodon | F | Hepatopancreas | 20–752.6 pg mg–1 | [37] |
| E | Penaeus japonicus | F | Hemolymph | 2.8–8.7 pg mL–1 | [38] |
| E | Gammarus duebeni | F | Hemolymph | 86.6 pg mL–1 | [28] |
| E | Gammarus duebeni | M | Hemolymph | 41.6 pg mL–1 | [28] |
| E | Gammarus pulex | F | Hemolymph | 38.8 pg mL–1 | [28] |
| E | Gammarus pulex | M | Hemolymph | 47.4 pg mL–1 | [28] |
| E | Gammarus pseudolimnaeus | F | Hemolymph | 18.8 pg mL–1 | [28] |
| E | Gammarus pseudolimnaeus | M | Hemolymph | 29.3 pg mL–1 | [28] |
| E | Scylla paramamosain | F | Hemolymph | 18.5–75.4 pg mL–1 | [27] |
| E | Emertia emeritus | F | Ovary | 2000–8000 pg g–1 | [26] |
| E | Emertia emeritus | F | Hepatopancreas | 2900–9700 pg g–1 | [26] |
| E | Macrobrachium rosenbergii | F | Ovary | 67–341 pg g–1 | [26] |
| E | Macrobrachium rosenbergii | F | Hepatopancreas | 168–663 pg g–1 | [26] |
| E | Eriocheir sinensis | F | Hemolymph | 118.8–291.8 pg mL–1 | [33] |
| E | Eriocheir sinensis | F | Ovary | 232.2–257.8 pg g–1 | [33] |
| E | Eriocheir sinensis | M | Hemolymph | 122.7–237.6 pg mL–1 | [33] |
| E | Eriocheir sinensis | M | Testes | 128.5–150.0 pg g–1 | [33] |
| E | Spiralothelphusa senex | F | Hemolymph | 27.9–134.2 pg mL–1 | [34] |
| E | Mictyris brevidactylus | F | Hemolymph | 0.2–0.6 pg g–1 | [35] |
| E | Mictyris brevidactylus | F | Ovary | 8.8–70.0 pg g–1 | [35] |
| E | Mictyris brevidactylus | F | Hepatopancreas | 0.6–122.9 pg g–1 | [35] |
| T | Nephrops norvegicus | F | Ovary | 19000 pg g–1 | [31] |
| T | Homarus americanus | M | Testes | 14300 pg g–1 | [39] |
| T | Gammarus duebeni | F | Hemolymph | 25.3 pg mL–1 | [28] |
| T | Gammarus duebeni | M | Hemolymph | 15.6 pg mL–1 | [28] |
| T | Gammarus pulex | F | Hemolymph | 22.1 pg mL–1 | [28] |
| T | Gammarus pulex | M | Hemolymph | 22.7 pg mL–1 | [28] |
| T | Gammarus pseudolimnaeus | F | Hemolymph | 21.7 pg mL–1 | [28] |
| T | Gammarus pseudolimnaeus | M | Hemolymph | 31.4 pg mL–1 | [28] |
| T | Scylla paramamosain | M | Hemolymph | 394–607 pg mL–1 | [27] |
| T | Penaeus japonicus | F | Hemolymph | 2.2–4.4 pg mL–1 | [38] |
| T | Neocaridina denticulata | ND | Hemolymph | 2700 pg g–1 | [40] |
Note: M—male, F—female, ND—no data, E—17β-estradiol, T—testosterone.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dvoretsky, A.G.; Tipisova, E.V.; Elfimova, A.E.; Alikina, V.A.; Dvoretsky, V.G. Sex Hormones in Hemolymph of Red King Crabs from the Barents Sea. Animals 2021, 11, 2149. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11072149
AMA Style
Dvoretsky AG, Tipisova EV, Elfimova AE, Alikina VA, Dvoretsky VG. Sex Hormones in Hemolymph of Red King Crabs from the Barents Sea. Animals. 2021; 11(7):2149. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11072149
Chicago/Turabian StyleDvoretsky, Alexander G., Elena V. Tipisova, Aleksandra E. Elfimova, Viktoria A. Alikina, and Vladimir G. Dvoretsky. 2021. "Sex Hormones in Hemolymph of Red King Crabs from the Barents Sea" Animals 11, no. 7: 2149. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11072149
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.