
Batoid Abundances, Spatial Distribution, and Life History Traits in the Strait of Sicily (Central Mediterranean Sea): Bridging a Knowledge Gap through Three Decades of Survey

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
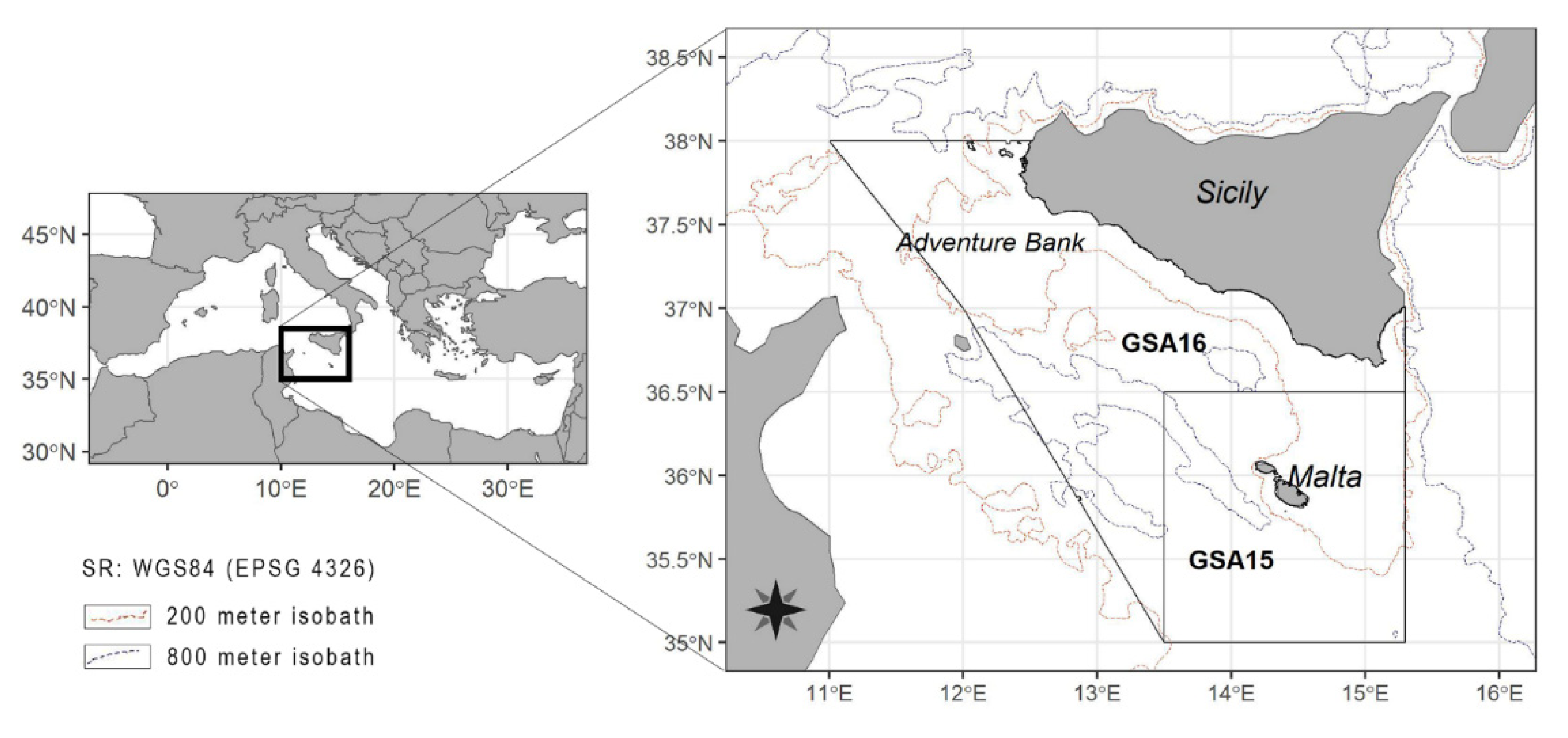
2.1. Study Area and Sampling Methodology
2.2. Abundance Data and Biological Parameter Analyses
- (a)
- frequency of occurrence as percentage of positive hauls (f%) and survey (s%);
- (b)
- two abundance indices: in weight, Biomass Index (BI; kg/km2), and number, Density Index (DI; N/km2), expressed as grand mean (the mean of the mean) with their relative standard error; f% and both abundance indices were estimated for the continental shelf (10–200 m), slope (200–800 m), and overall (10–800 m) depth stratum;
- (c)
- depth presence among the identified five microstrata;
- (d)
- correlation among survey abundance indexes and years were assessed by GSA, species, and macrostratum, by computing the nonparametric Spearman linear rank coefficient;
- (e)
- overall standing stock expressed in weight (tons) and number (thousand);
- (f)
- biological information in terms of overall sex ratio (sex ratio = F/M), median length, and length–weight relationship (LWR, power function; k will indicate the steepness; b will indicate the positive (b > 3) or negative (b < 3) allometric coefficient; isometric when b = 3) only from MEDITS16 because of the longer time series.
- (i)
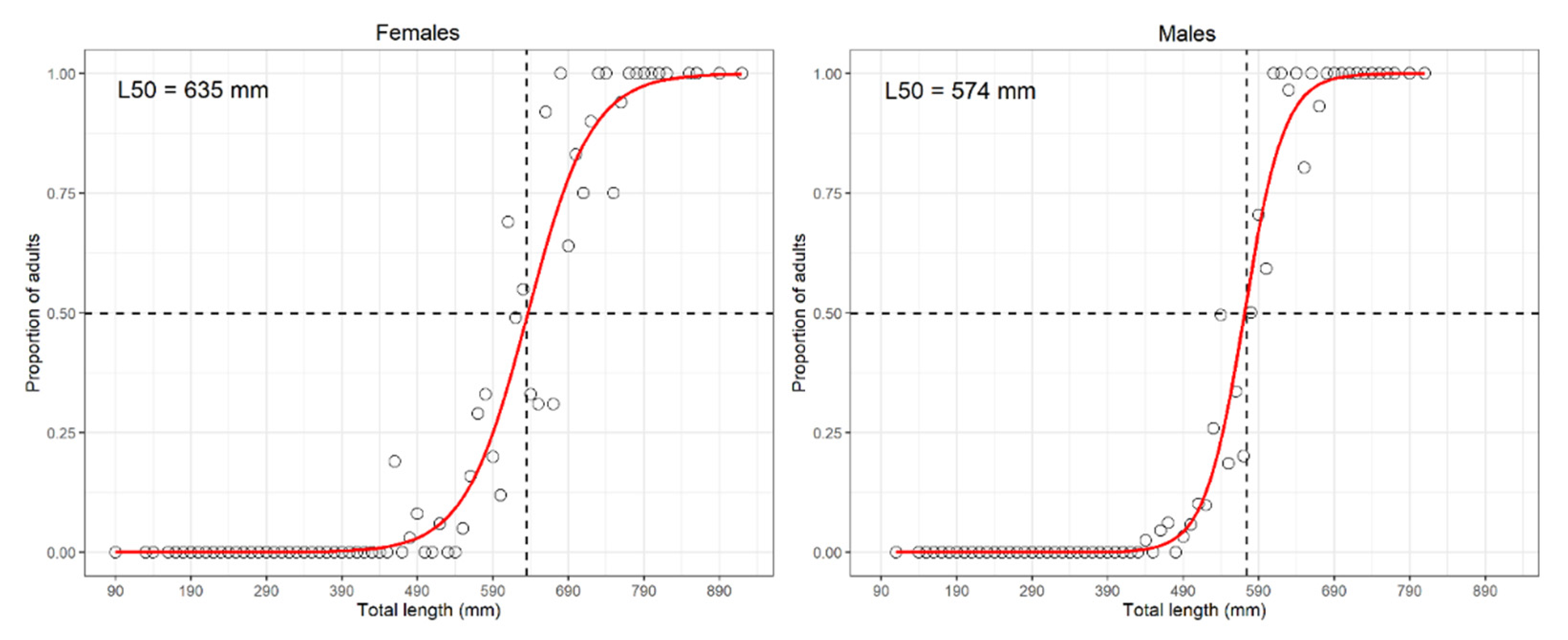
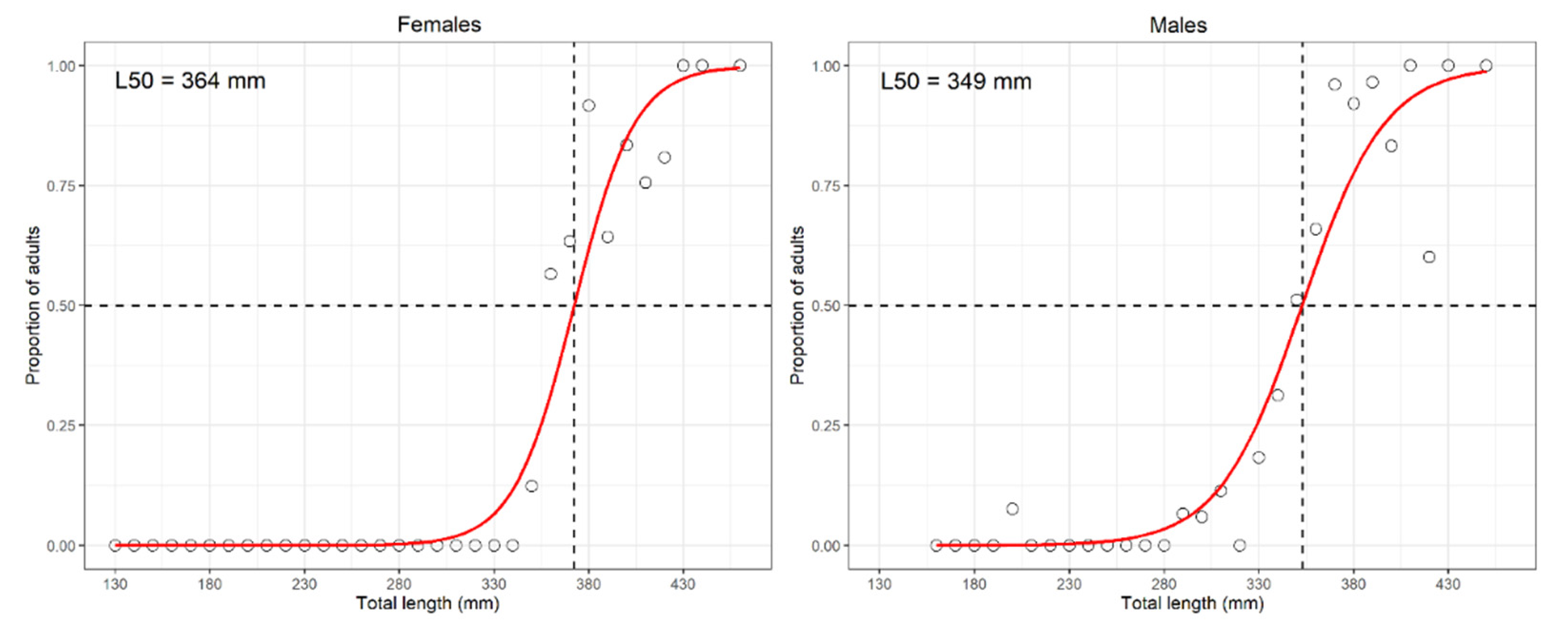
- length at first sexual maturity (L50, also known as at onset of sexual maturity), i.e., the length at which 50% of specimens resulted mature (stage 3a onward according to MEDITS gonadic scale [18]), or in case of lack of fit of the logistic using the median length of the 3a stage as a proxy of L50 [43];
- (ii)
- Length–Frequency Distribution (LFD) “stability” in the different years (performed via the Kolmogorov–Smirnov test);
- (iii)
- L∞ (the infinite/asymptotic length at which the average growth rate of the oldest cohort becomes close to zero) and K (Brody or curvature coefficient), both related to the von Bertalanffy growth function or;
- (iv)
- Z/K ratio by sex [44] at 95% confidence interval, where Z denotes the instantaneous rate of total mortality (as fishing, F, plus natural mortalities, M). The previous parameters were estimated using the ELEFAN (Electronic Length Frequencies Analysis) and Powell–Wetherall procedures in the TropFishR package [45]. All the size measurements were expressed as the Total Length (TL) in mm.
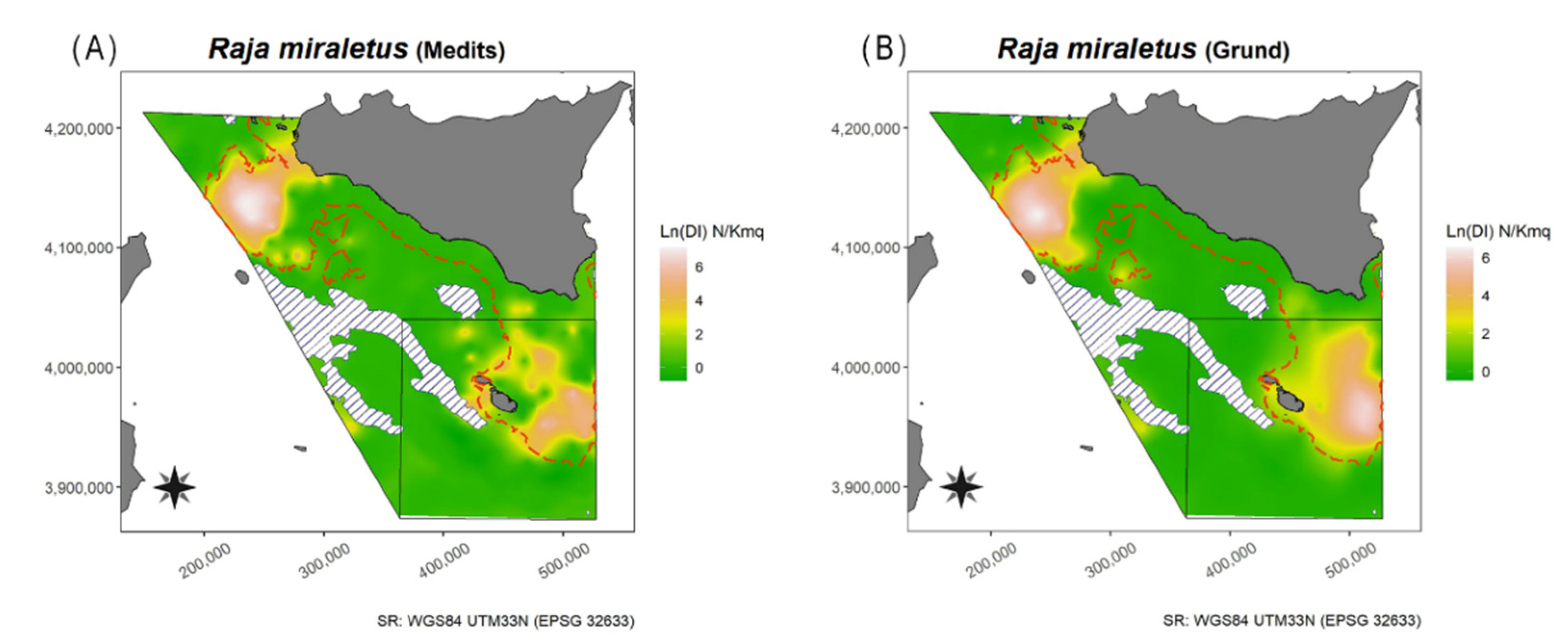
2.3. Spatial Analysis
3. Results
3.1. General
3.1.1. Great Torpedo Ray—Tetronarce nobiliana (Bonaparte, 1835)
3.1.2. Marbled Electric Ray—Torpedo marmorata Risso, 1810
3.1.3. Common Torpedo—Torpedo torpedo (Linnaeus, 1758)
3.1.4. Gray Skate—Dipturus batis (Linnaeus, 1758)
3.1.5. Norwegian Skate—Dipturus nidarosiensis (Storm, 1881)
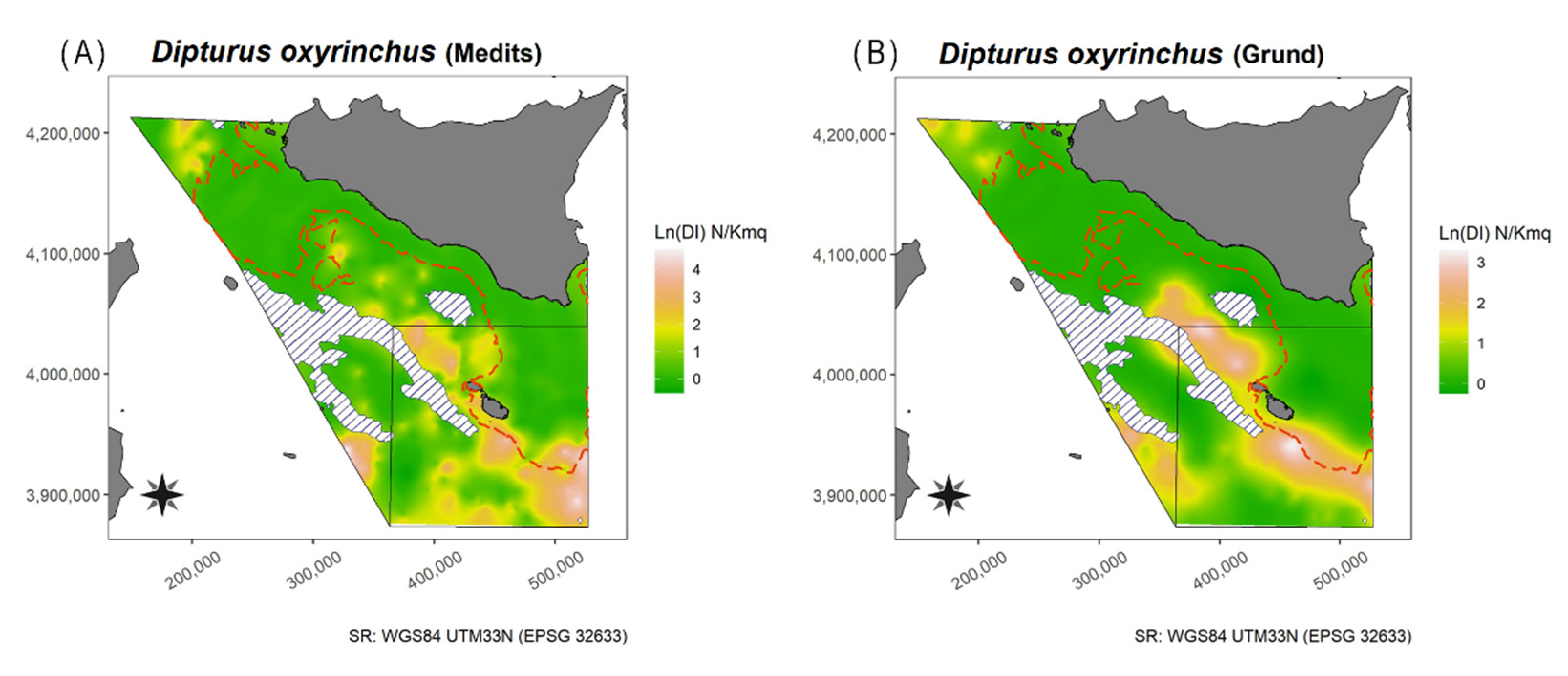
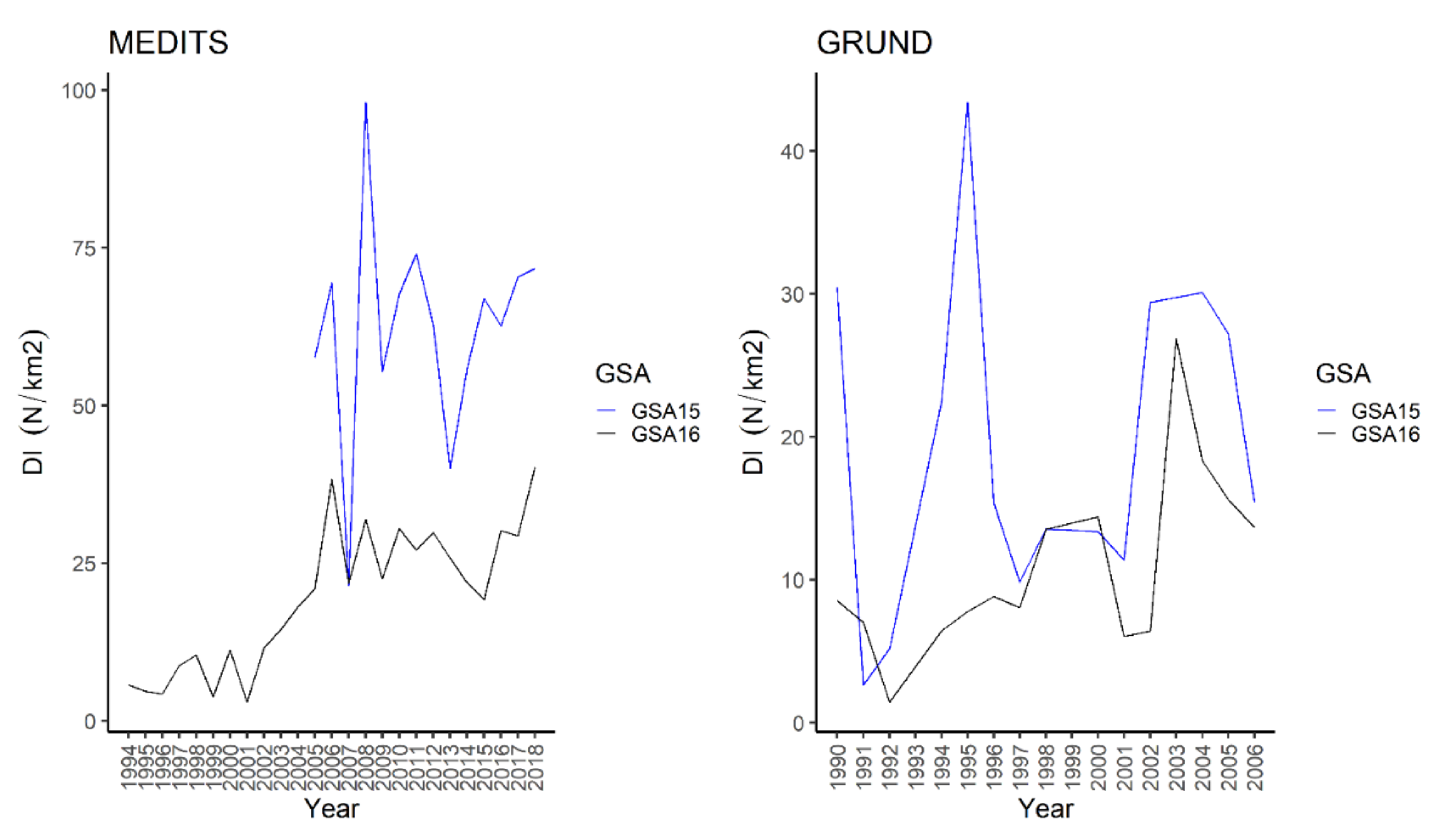
3.1.6. Longnosed Skate—Dipturus oxyrinchus (Linnaeus, 1758)
3.1.7. Sandy Ray—Leucoraja circularis (Couch, 1838)
3.1.8. Shagreen Ray—Leucoraja fullonica (Linnaeus, 1758)
3.1.9. Maltese Ray—Leucoraja melitensis (Clark, 1926)
3.1.10. Cuckoo Ray—Leucoraja naevus (Müller and Henle, 1841)
3.1.11. Starry Ray—Raja asterias Delaroche, 1809
3.1.12. Blonde Ray—Raja brachyura Lafont, 1873
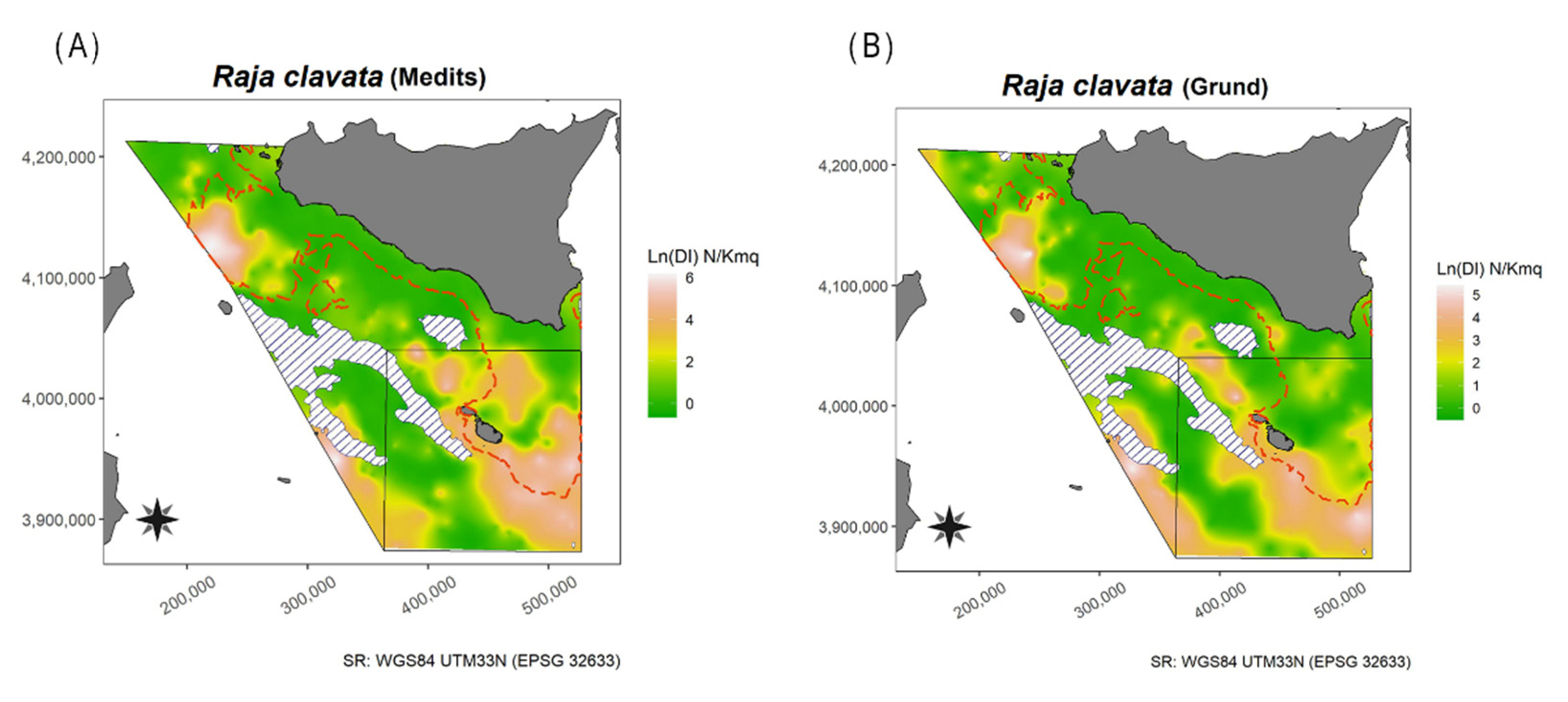
3.1.13. Thornback Ray—Raja clavata Linnaeus, 1758
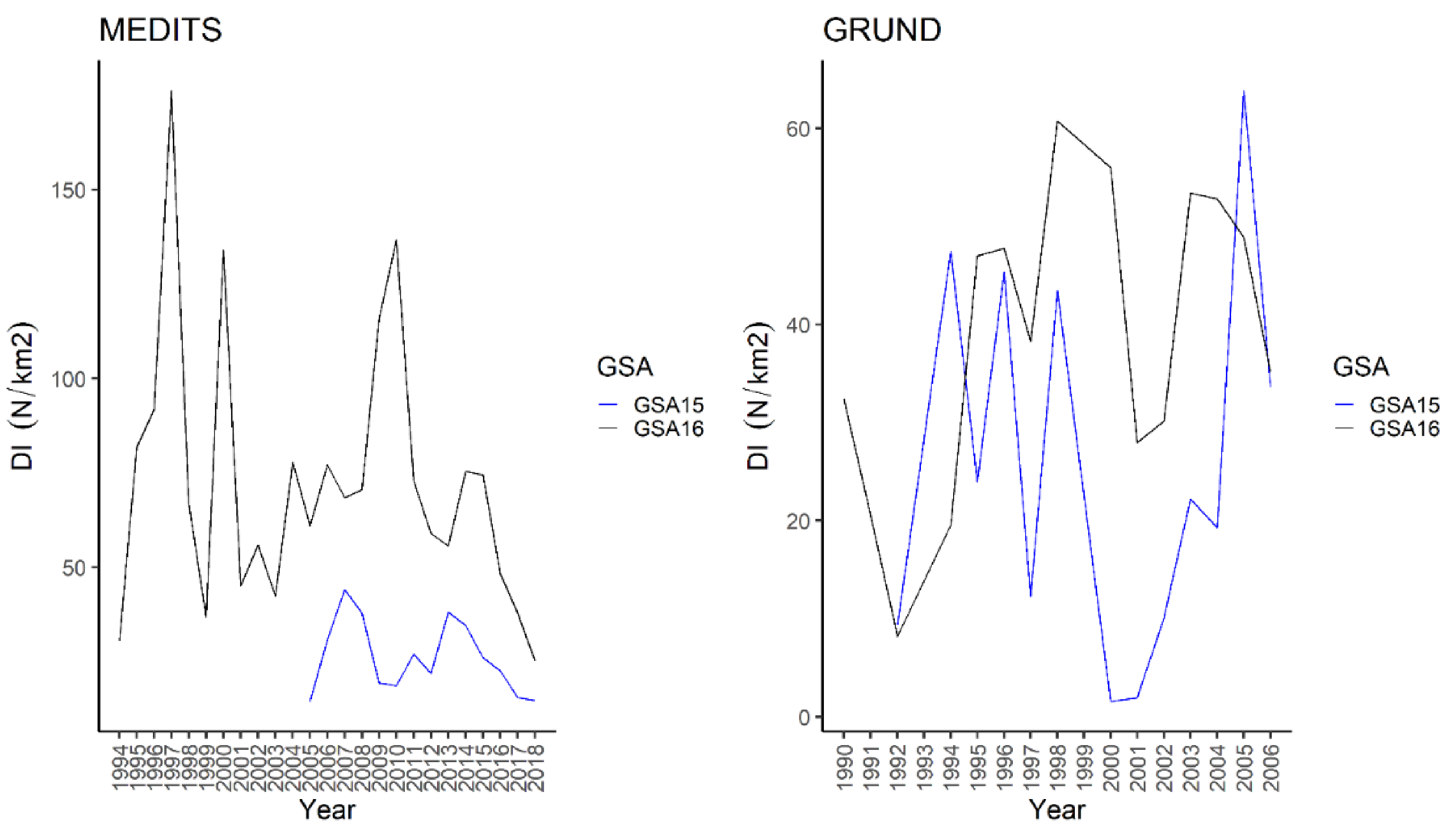
3.1.14. Brown Ray—Raja miraletus Linnaeus, 1758
3.1.15. Spotted Ray—Raja montagui Fowler, 1910
3.1.16. Speckled Ray—Raja polystigma Regan, 1923
3.1.17. Rough Ray—Raja radula Delaroche, 1809
3.1.18. Undulate Ray—Raja undulata Lacepède, 1802
3.1.19. White Skate—Rostroraja alba (Lacepède, 1803)
3.1.20. Common Stingray—Dasyatis pastinaca (Linnaeus, 1758)
3.1.21. Blue Stingray—Pteroplatytrygon violacea (Bonaparte, 1832)
3.1.22. Bull Ray—Aetomylaeus bovinus (Geoffroy Saint-Hilaire, 1817)
3.1.23. Common Eagle Ray—Myliobatis aquila (Linnaeus, 1758)
4. Discussion
4.1. General Consideration
4.1.1. Great Torpedo Ray—Tetronarce nobiliana (Bonaparte, 1835)
4.1.2. Marbled Electric Ray—Torpedo marmorata Risso, 1810
4.1.3. Common Torpedo—Torpedo torpedo (Linnaeus, 1758)
4.1.4. Gray Skate—Dipturus batis (Linnaeus, 1758)
4.1.5. Norwegian Skate—Dipturus nidarosiensis (Storm, 1881)
4.1.6. Longnosed Skate—Dipturus oxyrinchus (Linnaeus, 1758)
4.1.7. Sandy Ray—Leucoraja circularis (Couch, 1838)
4.1.8. Shagreen Ray—Leucoraja fullonica (Linnaeus, 1758)
4.1.9. Maltese Ray—Leucoraja melitensis (Clark, 1926)
4.1.10. Cuckoo Ray—Leucoraja naevus (Müller and Henle, 1841)
4.1.11. Starry Ray—Raja asterias Delaroche, 1809
4.1.12. Blonde Ray—Raja brachyura Lafont, 1873
4.1.13. Thornback Ray—Raja clavata Linnaeus, 1758
4.1.14. Brown Ray—Raja miraletus Linnaeus, 1758
4.1.15. Spotted Ray—Raja montagui Fowler, 1910
4.1.16. Speckled Ray—Raja polystigma Regan, 1923
4.1.17. Rough Ray—Raja radula Delaroche, 1809
4.1.18. Undulate Ray—Raja undulata Lacepède, 1802
4.1.19. White Skate—Rostroraja alba (Lacepède, 1803)
4.1.20. Common Stingray—Dasyatis pastinaca (Linnaeus, 1758)
4.1.21. Blue Stingray—Pteroplatytrygon violacea (Bonaparte, 1832)
4.1.22. Bull Ray—Aetomylaeus bovinus (Geoffroy Saint-Hilaire, 1817)
4.1.23. Common Eagle Ray—Myliobatis aquila (Linnaeus, 1758)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McEachran, J.D.; de Carvalho, M.R.; Carpenter, K.E. (Eds.) Batoid fishes. In The Living Marine Resources of the Western Central Atlantic; Introduction, Molluscs, Crustaceans, Hagfishes, Sharks, Batoid Fishes, and Chimaeras; FAO Species Identification Guide for Fishery Purposes; American Society of Ichthyologist and Herpetologists Special Publication No. 5; FAO: Rome, Italy, 2002; Volume 1, pp. 507–589. [Google Scholar]
- Iglesias, S.P. Handbook of the Marine Fishes of Europe and Adjacent Waters (A Natural Classification Based on Collection Specimens, with DNA Barcodes and Standardized Photographs). Volume I (Chondrichthyans and Cyclostomata). Provisional Version 08. 2014, p. 105. Available online: http://www.mnhn.fr/iccanam (accessed on 7 March 2019).
- Serena, F.; Abella, A.J.; Bargnesi, F.; Barone, M.; Colloca, F.; Ferretti, F.; Fiorentino, F.; Jenrette, J.; Moro, S. Species diversity, taxonomy and distribution of Chondrichthyes in the Mediterranean and Black Sea. Eur. Zool. J. 2020, 87, 497–536. [Google Scholar] [CrossRef]
- Guijarro, B.; Quetglas, A.; Moranta, J.; Ordines, F.; Valls, M.; González, N.; Massutí, E. Inter- and intra-annual trends and status indicators of nektobenthic elasmobranchs off the Balearic Islands (northwestern Mediterranean). Sci. Mar. 2012, 76, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Ligas, A.; Osio, G.C.; Sartor, P.; Sbrana, M.; De Ranieri, S. Long-term trajectory of some elasmobranch species off the Tuscany coasts (NW Mediterranean) from 50 years of catch data. Sci. Mar. 2013, 77, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, F.; Myers, R.A.; Serena, F.; Lotze, H.K. Loss of Large Predatory Sharks from the Mediterranean Sea. Conserv. Biol. 2008, 22, 952–964. [Google Scholar] [CrossRef]
- Walls, R.H.L.; Dulvy, N.K. Predicting the conservation status of Europe’s Data Deficient sharks and rays. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, M.; Bartolino, V.; Llope, M.; Maiorano, L.; Sköld, M.; Hagberg, J. Historical spatial baselines in conservation and management of marine resources. Fish Fish. 2010, 12, 289–298. [Google Scholar] [CrossRef]
- Abella, A. General review on the available methods for stock assessment of Elasmobranchs, especially in data shortage situations. In Sub-Committee on Stock Assessment (SCSA) Report of the Workshop on Stock Assessment of Selected Species of Elasmobranchs in the GFCM Area DG-MARE; GFCM-FAO: Brussels, Belgium, 2011; pp. 12–16. [Google Scholar]
- Booth, H.; Squires, D.; Milner-Gulland, E.J. The neglected complexities of shark fisheries, and priorities for holistic risk-based management. Ocean Coast. Manag. 2019, 182, 104994. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Allen, D.J.; Ralph, G.M.; Walls, R.H. The Conservation Status of Sharks, Rays and Chimaeras in the Mediterranean Sea [Brochure]; IUCN Centre for Mediterranean Cooperation: Malaga, Spain, 2016; p. 14. [Google Scholar]
- Milazzo, M.; Cattano, C.; Al Mabruk, S.A.A.; Giovos, I. Mediterranean sharks and rays need action. Science 2021, 371, 355–356. [Google Scholar] [CrossRef]
- Flowers, K.I.; Heithaus, M.R.; Papastamatiou, Y.P. Buried in the sand: Uncovering the ecological roles and importance of rays. Fish Fish. 2021, 22, 105–127. [Google Scholar] [CrossRef]
- Pratt, H.L., Jr.; Gruber, S.H.; Taniuchi, T. Elasmobranchs as living resources: Advances in the biology, ecology, systematics and the status of the fisheries. In NOAA Technical Report NMFS; National Marine Fisheries Service: Silver Spring, MD, USA, 1990; 518p. [Google Scholar]
- Dayton, P.K.; Thrush, S.F.; Agardy, M.T.; Hofman, R.J. Environmental effects of marine fishing. Aquat Conserv. 1995, 5, 205–232. [Google Scholar] [CrossRef]
- Stevens, J. The effects of fishing on sharks, rays, and chimaeras (chondrichthyans), and the implications for marine ecosystems. ICES J. Mar. Sci. 2000, 57, 476–494. [Google Scholar] [CrossRef]
- Nieto, A.; Ralph, G.M.; Comeros-Raynal, M.T.; Heessen, H.J.; Rijnsdorp, A.D. European Red List of Marine Fishes; Publications Office of the European Union: Luxembourg, 2015; p. 61. [Google Scholar]
- MEDITS Working Group. International Bottom Trawl Survey in the Mediterranean; Instruction manual; Version 9. [MEDITS–handbook. Version n. 9.]; MEDITS Working Group, 2017; p. 106. [Google Scholar]
- Spedicato, M.T.; Massutí, E.; Mérigot, B.; Tserpes, G.; Jadaud, A.; Relini, G. The MEDITS trawl survey specifications in an ecosystem approach to fishery management. Sci. Mar. 2019, 83, 9–20. [Google Scholar] [CrossRef]
- Jereb, P.; Cannizzaro, L.; Norrito, G.; Ragonese, S. Strait of Sicily’: Sensu stricto vs sensu lato. Setting a baseline definition for an important Mediterranean fisheries management area. Biol. Mar. Mediterr. 2016, 69, 49–56. (In Italian) [Google Scholar]
- Cannizzaro, L.; Garofalo, G.; Levi, D.; Rizzo, P.; Gancitano, S. Raja clavata (Linneo, 1758) nel Canale di Sicilia: Crescita, distribuzione e abbondanza. Biol. Mar. Mediterr. 1995, 2, 257–262. (In Italian) [Google Scholar]
- Ragonese, S.; Cigala Fulgosi, F.; Bianchini, M.L.; Norrito, G.; Sinacori, G. Annotated check list of the skates (Chondrichthyes, Rajidae) in the Strait of Sicily (Central Mediterranean). Biol. Mar. Mediterr. 2003, 10, 874–881. (In Italian) [Google Scholar]
- Lauria, V.; Gristina, M.; Attrill, M.J.; Fiorentino, F.; Garofalo, G. Predictive habitat suitability models to aid conservation of elasmobranch diversity in the central Mediterranean Sea. Sci. Rep. 2015, 5, 13245. [Google Scholar] [CrossRef] [Green Version]
- Abella, A.; Mancusi, C.; Serena, F. Raja asterias. In Sintesi delle Conoscenze di Biologia, Ecologia e pesca delle Specie Ittiche dei mari Italiani; Sartor, P., Mannini, A., Carlucci, R., Massaro, E., Queirolo, S., Sabatini, A., Scarcella, G., Simoni, R., Eds.; Società Italiana di Biologia Marina (S.I.B.M.): Genoa, Italy, 2017; pp. 144–149. [Google Scholar]
- Abella, A.; Mancusi, C.; Serena, F. Raja clavata. In Sintesi delle Conoscenze di Biologia, Ecologia e pesca delle Specie Ittiche dei mari Italiani; Sartor, P., Mannini, A., Carlucci, R., Massaro, E., Queirolo, S., Sabatini, A., Scarcella, G., Simoni, R., Eds.; Società Italiana di Biologia Marina (S.I.B.M.): Genoa, Italy, 2017; pp. 150–156. [Google Scholar]
- Geraci, M.L.; Ragonese, S.; Norrito, G.; Scannella, D.; Falsone, F.; Vitale, S. A Tale on the Demersal and Bottom Dwelling Chondrichthyes in the South of SicilySouth Sicily through 20 Years of Scientific Survey. In Chondrichthyes—Multidisciplinary Approach; Rodrigues-Filho, L.F., de Luna Sales, J.B., Eds.; IntechOpen: London, UK, 2017; pp. 13–37. [Google Scholar] [CrossRef] [Green Version]
- Follesa, M.C.; Marongiu, M.F.; Zupa, W.; Bellodi, A.; Cau, A.; Cannas, R.; Colloca, F.; Djurovic, M.; Isajlovic, I.; Jadaud, A.; et al. Spatial variability of Chondrichthyes in the northern Mediterranean. Sci. Mar. 2019, 83, 81–100. [Google Scholar] [CrossRef] [Green Version]
- Schembri, T.; Schembri, P.J.; Fergusson, I.K. Revision of the records of shark and ray species from the Maltese Islands (Chordata: Chondrichthyes). Cent. Medit. Nat. 2003, 4, 71–104. [Google Scholar]
- Bradai, M.N.; Saidi, B.; Enajjar, S. Elasmobranchs of the Mediterranean and Black Sea: Status, Ecology and Biology; Bibliographic analysis; Studies and Reviews-General Fisheries Commission for the Mediterranean; No. 91; FAO: Rome, Italy, 2012. [Google Scholar]
- Kadri, H.; Marouani, S.; Sadi, B.; Bradai, M.N.; Ghorbel, M.; Bouan, A.; Morize, E. Age, growth and reproduction of Raja miraletus (Linnaeus, 1758) (Chondrichthyes: Rajidae) of the Gulf of Gabès (Tunisia, Central Mediterranean Sea). Mar. Biol. Res. 2012, 8, 388–396. [Google Scholar] [CrossRef]
- Kadri, H.; Marouani, S.; Bradai, M.N.; Bouaïn, A.; Morize, E. Age, growth, longevity, mortality and reproductive biology of Dipturus oxyrinchus, (Chondrichthyes: Rajidae) off the Gulf of Gabès (Southern Tunisia, central Mediterranean). J. Mar. Biol. Assoc. UK 2014, 95, 569–577. [Google Scholar] [CrossRef]
- Kadri, H.; Marouani, S.; Saïdi, B.; Bradai, M.N.; Bouaïn, A.; Morize, E. Age, growth, sexual maturity and reproduction of the thornback ray, Raja clavata (L.), of the Gulf of Gabès (south-central Mediterranean Sea). Mar. Biol. Res. 2014, 10, 416–425. [Google Scholar] [CrossRef]
- Kadri, H.; Marouani, S.; Bradai, M.N.; Bouaïn, A.; Morize, E. Distribution and Morphometric Characters of the Mediterranean Brown Ray, Raja miraletus (Chondrichthyes: Rajidae) in the Gulf of Gabes (Tunisia, Central Mediterranean). Am. J. Agric. For. 2014, 2, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Kadri, H.; Marouani, S.; Bradai, M.N.; Bouaïn, A.; Morize, E. Age, growth and length-weight relationship of the rough skate, Raja radula (Linnaeus, 1758) (Chondrichthyans: Rajidae), from the Gulf of Gabes (Tunisia, Central Mediterranean). J. Coast. Life Med. 2014, 2, 344–349. [Google Scholar]
- Kadri, H.; Marouani, S.; Bradai, M.N.; Bouaïn, A.; Morize, E. Age, growth and length-weight relationship of the white skate, Rostroraja alba (Linnaeus, 1758) (Chondrichthyans: Rajidae), from the Gulf of Gabes (Tunisia, Central Mediterranean). J. Coast. Life Med. 2014, 14, 193–204. [Google Scholar] [CrossRef]
- Bradai, M.N.; Saidi, B.; Enajjar, S. Overview on Mediterranean shark’s fisheries: Impact on the biodiversity. In Marine Ecology–Biotic and Abiotic Interactions; Turkoglu, M., Onal, U., Ismen, A., Eds.; IntechOpen: London, UK, 2018; pp. 211–230. [Google Scholar] [CrossRef] [Green Version]
- GFCM. Resolution 31/2007/2: On the Establishment of GeographicalSub-Areas in the GFCM Area. 2007. Available online: http://www.gfcm.org/figis/pdf/gfcm/topic/16162/en?title=GFCM%20-%20Geographical%20Sub-Areas (accessed on 15 December 2013).
- Levi, D.; Ragonese, S.; Andreoli, M.G.; Norrito, G.; Rizzo, P.; Giusto, G.B.; Gancitano, S.; Sinacori, G.; Bono, G.; Garofalo, G.; et al. Sintesi delle ricerche sulle risorse demersali dello Stretto di Sicilia (Mediterraneo centrale) negli anni 1985–1997 svolte nell’ambito della legge 41/82. Biol. Mar. Mediterr. 1998, 5, 130–139. (In Italian) [Google Scholar]
- Relini, G. Demersal trawl surveys in Italian Seas: A short review. Actes Colloq. Ifremer 2000, 26, 76–93. [Google Scholar]
- Papaconstantinou, C.; Relini, G.; Souplet, A. The general specifications of the Medits surveys. Sci. Mar. 2002, 66, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Ragonese, S.; Vitale, S. Desirability of a standard notation for fisheries assessment. Agric. Sci. 2013, 04, 399–432. [Google Scholar] [CrossRef] [Green Version]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999; 662p. [Google Scholar]
- Lembo, G. SAMED Stock Assessment in the Mediterranean; EC project n° 99/047; COISPA: Bari, Italy, 2002. [Google Scholar]
- Wetherall, J.A.; Polovina, J.J.; Ralston, S. Estimating growth and mortality in steady-state fish stocks from length-frequency data. In ICLARM Conference Proceedings; WorldFish: Penang, Malaysia, 1987; Volume 13, pp. 53–74. [Google Scholar]
- Mildenberger, T.K.; Taylor, M.H.; Wolff, M. TropFishR: An R package for fisheries analysis with length-frequency data. Methods Ecol. Evol. 2017, 8, 1520–1527. [Google Scholar] [CrossRef] [Green Version]
- Petitgas, P. Geostatistics and their applications to fisheries survey data. In Computers in Fisheries Research; Megrey, B.A., Moksness, E., Eds.; Chapman &Hall: London, UK, 1996; pp. 113–142. [Google Scholar]
- Goovaerts, P. Geostatistics for Natural Resources Evaluation; Oxford University Press: New York, NY, USA, 1997. [Google Scholar]
- Matheron, G. La theorie des Variables Regionalisees et ses Applications. Cahiers du Centre de Morphologie Mathématique de Fontainebleau; École Nationale Supérieure des Mines: Paris, France, 1970; Volume 5, pp. 1–212. (In French) [Google Scholar]
- Isaaks, E.H.; Srivastava, R.M. Applied Geostatistics; Oxford University Press: New York, NY, USA, 1989. [Google Scholar]
- Rossi, R.E.; Mulla, D.J.; Journel, A.G.; Franz, E.H. Geostatistical tools for modeling and interpreting ecological spatial dependence. Ecol. Monogr. 1992, 62, 277–314. [Google Scholar] [CrossRef]
- Pebesma, E.J. Multivariable geostatistics in S: The gstat package. Comput. Geosci. 2004, 30, 683–691. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; Version 3.6.2; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 15 March 2020).
- Geraci, M.L.; Di Lorenzo, M.; Falsone, F.; Scannella, D.; Di Maio, F.; Colloca, F.; Vitale, S.; Serena, F. The occurrence of Norwegian skate, Dipturus nidarosiensis (Elasmobranchii: Rajiformes: Rajidae), in the Strait of Sicily, central Mediterranean. Acta Ichthyol. Piscat. 2019, 49, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Ferrá, C.; Fabi, G.; Polidori, P.; Tassetti, A.N.; Leoni, S.; Pellini, G.; Scarcella, G. Raja asterias population assessment in FAO GFCM GSA17 area. Mediterr. Mar. Sci. 2016, 17, 651–660. [Google Scholar] [CrossRef] [Green Version]
- Cashion, M.S.; Bailly, N.; Pauly, D. Official catch data underrepresent shark and ray taxa caught in Mediterranean and Black Sea fisheries. Mar. Policy. 2019, 105, 1–9. [Google Scholar] [CrossRef]
- Abella, A.J.; Serena, F. Comparison of Elasmobranch Catches from Research Trawl Surveys and Commercial Landings at Port of Viareggio, Italy, in the Last Decade. J. Northwest Atl. Fish. Sci. 2005, 35, 345–356. [Google Scholar] [CrossRef]
- Giudicelli, M. Malta: Simulated Commercial Trawling and Scouting Operations in the Central Mediterranean (January 1976–June 1977); FI: MAT/75/001/1; FAO: Rome, Italy, 1978; 93p. [Google Scholar]
- Garofalo, G.; Gristina, M.; Fiorentino, F.; Cigala Fulgosi, F.; Norrito, G.; Sinacori, G. Distributional pattern of rays (Pisces, Rajidae) in the Strait of Sicily in relation to fishing pressure. Hydrobiologia 2003, 503, 245–250. [Google Scholar] [CrossRef]
- Ragonese, S.; Vitale, S.; Dimech, M.; Mazzola, S. Abundances of demersal sharks and chimaera from 1994-2009 scientific surveys in the central Mediterranean Sea. PLoS ONE 2013, 8, e74865. [Google Scholar] [CrossRef] [Green Version]
- Lanfranco, G. The Fish around Malta (Central Mediterranean); Progress Press: Valletta, Malta, 1993. [Google Scholar]
- Farrugia Randon, S.; Sammut, R. Fishes of Maltese Waters; The Authors: Malta, 1999. [Google Scholar]
- Sammut, R. Mediterranean Sea Fishes Central Region; The Authors: Malta, 2001. [Google Scholar]
- Arena, P.; Li Greci, F. Indagine sulle condizioni faunistiche e sui rendimenti di pesca dei fondali batiali della Sicilia occidentale e della bordura settentrionale dei banchi della soglia Siculo-Tunisina. Quad. Lab. Tecnol. Pesca 1973, 1, 157–201. (In Italian) [Google Scholar]
- Maurin, C. Etude des fonds chalutables de la Mediterranee occidentale (Ecologie et Peche). Rev. des Trav. de l’Institut des Pêches Marit. 1968, 26, 163–218. (In French) [Google Scholar]
- Quignard, J.P.; Capapé, C. Liste commentée des Sélaciens de Tunisie 2. Bull. Inst. Océanogr. Pêche Salammbô 1971, 2, 131–142. (In French) [Google Scholar]
- Başusta, A.; Başusta, N.; Sulikowski, J.A.; Driggers, W.B.; Demirhan, S.A.; Çiçek, E. Length-weight relationships for nine species of batoids from the Iskenderun Bay, Turkey. J. Appl. Ichthyol. 2012, 28, 850–851. [Google Scholar] [CrossRef]
- Kaya, G.; Başusta, N. A study on age and growth of juvenile and semi adult Torpedo nobiliana Bonaparte, 1835 inhabiting Iskenderun Bay, northeastern Mediterranean Sea. Acta Biol. Turc. 2016, 29, 143–149. [Google Scholar]
- Bini, G. Atlante dei Pesci delle Coste Italiane. Milano: Mondo Sommerso; Vol. 1 Leptocardi, Ciclostomi, Selaci, Mondo Sommerso: Roma, Italy, 1967; 206p. (In Italian) [Google Scholar]
- Baino, R.; Serena, F.; Ragonese, S.; Rey, J.; Rinelli, P. Catch composition and abundance of elasmobranchs based on the MEDITS program. Rapp. Comm. Int. Mer. Médit. 2001, 36, 234. [Google Scholar]
- Bradai, M.N. Diversité du Peuplement Ichtyque et Contribution à la Connaisance des Sparidés du Golfe de Gabès. Ph.D. Thesis, University of Sfax, Sfax, Tunisia, 2000. (In French). [Google Scholar]
- Duman, Ö.V.; Başusta, N. Age and growth characteristics of marbled electric ray Torpedo marmorata (Risso, 1810) inhabiting Iskenderun Bay, North-eastern Mediterranean Sea. Turk. J. Fish Aquat. Sci. 2013, 13, 541–549. [Google Scholar] [CrossRef]
- Yeldan, H.; Gundogdu, S. Morphometric relationships and growth of common stingray, Dasyatis pastinaca (Linnaeus, 1758) and marbled stingray, Dasyatis marmorata (Steindachner, 1892) in the northeastern Levantine Basin. J. Black Sea/Mediterr. Environ. 2018, 24, 10–27. [Google Scholar]
- Filiz, H.; Bilge, G. Length-weight relationships of 24 fish species from the North Aegean Sea, Turkey. J. Appl. Ichthyol. 2004, 20, 431–432. [Google Scholar] [CrossRef]
- Bellodi, A.; Mulas, A.; Carbonara, P.; Cau, A.; Cuccu, D.; Marongiu, M.F.; Mura, V.; Pesci, P.; Zupa, W.; Porcu, C.; et al. New insights into life–history traits of Mediterranean Electric rays (Torpediniformes: Torpedinidae) as a contribution to their conservation. Zoology 2021, 146, 125922. [Google Scholar] [CrossRef]
- Consalvo, I.; Scacco, U.; Romanelli, M.; Vacchi, M. Comparative study on the reproductive biology of Torpedo torpedo (Linnaeus, 1758) and T. marmorata (Risso, 1810) in the central Mediterranean Sea. Sci. Mar. 2007, 71, 213–222. [Google Scholar] [CrossRef]
- Lteif, M.; Mouawad, R.; Jemaa, S.; Khalaf, G.; Lenfant, P.; Verdoit-Jarraya, M. The length-weight relationships of three sharks and five batoids in the Lebanese marine waters, eastern Mediterranean. Egypt. J. Aquat. Res. 2016, 42, 475–477. [Google Scholar] [CrossRef]
- Abdel-Aziz, S. Observations on the biology of the common torpedo (Torpedo torpedo, Linnaeus, 1758) and marbled electric ray (Torpedo marmorata, Risso, 1810) from Egyptian Mediterranean waters. Mar. Freshw. Res. 1994, 45, 693–704. [Google Scholar] [CrossRef]
- El Kamel-Moutalibi, O.; Mnasri, N.; Boumaïza, M.; Ben Amor, M.M.; Reynaud, C.; Capapé, C. Maturity, reproductive cycle and fecundity of common torpedo, Torpedo torpedo (Chondrichthyes, Torpedinidae) from the Lagoon of Bizerte (Northeastern Tunisia, central Mediterranean). J. Ichthyol. 2013, 53, 758–774. [Google Scholar] [CrossRef]
- Serena, F.; Mancusi, C.; Barone, M. (Eds.) Field guide to the identification of the skates (Rajidae) of the Mediterranean Sea. Guidelines for data collection and analysis. Biol. Mar. Mediterr. 2010, 17, 1–204. [Google Scholar]
- Serena, F.; Mancusi, C.; Barone, M. MEDiterranean Large Elasmobranchs Monitoring. In Protocollo di Acquisizione Dati; SharkLife Program: Roma, Italy, 2014; 130p. (In Italian) [Google Scholar]
- Cannas, R.; Follesa, M.C.; Cabiddu, S.; Porcu, C.; Salvadori, S.; Iglésias, S.P.; Deiana, A.M.; Cau, A. Molecular and morphological evidence of the occurrence of the Norwegian skate Dipturus nidarosiensis (Storm, 1881) in the Mediterranean Sea. Mar. Biol. Res. 2010, 6, 341–350. [Google Scholar] [CrossRef]
- Cariani, A.; Messinetti, S.; Ferrari, A.; Arculeo, M.; Bonello, J.J.; Bonnici, L.; Cannas, R.; Carbonara, P.; Cau, A.; Charilaou, C.; et al. Improving the Conservation of Mediterranean Chondrichthyans: The ELASMOMED DNA Barcode Reference Library. PLoS ONE 2017, 12, e0170244. [Google Scholar] [CrossRef]
- Fang, K.; Chen, L.; Zhou, J.; Yang, Z.P.; Dong, X.F.; Zhang, H.B. On the presence of Dipturus nidarosiensis (Storm, 1881) in the Central Mediterranean area. PeerJ 2019, 7, e7009. [Google Scholar] [CrossRef] [Green Version]
- Isajlović, I.; Dragičević, B.; Manfredi, C.; Vrgoč, N.; Piccinetti, C.; Dulčić, J. Additional records of Norwegian skate Dipturus nidarosiensis (Storm, 1881) (Pisces: Rajidae) in the Adriatic Sea. Acta Adriat. 2020, 61, 217–222. [Google Scholar] [CrossRef]
- Ramírez-Amaro, S.; Ordines, F.; Puerto, M.Á.; García, C.; Ramon, C.; Terrasa, B.; Massutí, E. New morphological and molecular evidence confirm the presence of the Norwegian skate Dipturus nidarosiensis (Storm, 1881) in the Mediterranean Sea and extend its distribution to the western basin. Mediterr. Mar. Sci. 2017, 18, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Carugati, L.; Melis, R.; Cariani, A.; Cau, A.; Crobe, V.; Ferrari, A.; Follesa, M.C.; Geraci, M.L.; Iglésias, S.P.; Pesci, P.; et al. Combined COI barcode-based methods to avoid mislabelling of threatened species of deep-sea skates. Anim. Conserv. 2021. [Google Scholar] [CrossRef]
- Bellodi, A.; Porcu, C.; Cannas, R.; Cau, A.; Marongiu, M.F.; Mulas, A.; Vittori, S.; Follesa, M.C. Life-history traits of the long-nosed skate Dipturus oxyrinchus. J. Fish Biol. 2016, 90, 867–888. [Google Scholar] [CrossRef]
- Yığın, C.Ç.; Ismen, A. Age, growth, reproduction and feed of longnosed skate, Dipturus oxyrinchus (Linnaeus, 1758) in Saros Bay, the north Aegean Sea. J. Appl. Ichthyol. 2010, 26, 913–919. [Google Scholar] [CrossRef]
- Alkusairy, H.; Saad, A. Some morphological and biological aspects of longnosed skate, Dipturus oxyrinchus (Elasmobranchii: Rajiformes: Rajidae), in Syrian marine waters (eastern Mediterranean). Acta Ichthyol. Piscat. 2017, 47, 371–383. [Google Scholar] [CrossRef] [Green Version]
- Powell, D.G. Estimation of mortality and growth parameters from the length frequency of a catch [model]. Rapp. Proc. Verb. Reun. 1979, 175, 167–169. [Google Scholar]
- Ebert, D.A.; Stehmann, M.F. Sharks, Batoids and Chimaeras of the North Atlantic; FAO: Roma, Italy, 2013. [Google Scholar]
- Deval, M.C.; Güven, O.; Saygu, İ.; Kabapçioğlu, T. Length-weight relationships of 10 fish species found off Antalya Bay, eastern Mediterranean. J. Appl. Ichthyol. 2013, 30, 567–568. [Google Scholar] [CrossRef]
- Mnasri, N.; Boumaïza, M.; Capapé, C. Morphological data, biological observations and occurrence of a rare skate, Leucoraja circularis (Chondrichthyes: Rajidae), off the northern coast of Tunisia (central Mediterranean). Pan-Am. J. Aquat. Sci. 2009, 4, 70–78. [Google Scholar]
- Stehmann, M.; Bürkel, D.L. Rajidae. In Fishes of the Northeastern Atlantic and Mediterranean; Whitehead, P.J.P., Bauchot, M.L., Hureau, J.C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1984; pp. 163–196. [Google Scholar]
- Froese, R.; Thorson, J.; Reyes, R.B., Jr. A Bayesian approach for estimating length-weight relationships in fishes. J. Appl. Ichthyol. 2014, 30, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Bono, L.; De Ranieri, S.; Fabiani, O.; Mancusi, C.; Serena, F. Studio sull’accrescimento di Raja asterias (Delaroche, 1809) (Chondrichthyes, Raidae) attraverso l’analisi delle vertebre. Biol. Mar. Mediterr. 2005, 12, 470–474. (In Italian) [Google Scholar]
- Serena, F.; Abella, A. Sintesi delle conoscenze sulle risorse da pesca dei fondi del Mediterraneo centrale (Italia e Corsica). Biol Mar. Medit. 1999, 6, 82–86. [Google Scholar]
- Cau, A.; Follesa, M.C.; Pesci, P.; Cuccu, D.; Sabatini, A. Campionamento Biologico Delle Catture. Sezioni C ed E. Dipartimento di Scienze della Vita e dell’Ambiente. In Programma Nazionale Italiano per la Raccolta Dati Alieutici 2012; Università di Cagliari: Cagliari, Italy, 2013. (In Italian) [Google Scholar]
- Carbonara, P.; Spedicato, M.T.; Casciaro, L.; Bitetto, I.; Facchini, M.T.; Zupa, W.; Gaudio, P.; Surico, A.; Gallo, P.; Fazi, S.; et al. Campionamento Biologico delle catture. Sezioni C ed E. Rapporto Finale. GSA19 Mar Ionio. In Programma Nazionale Italiano per la Raccolta Dati Alieutici 2012; COISPA Tecnologia Ricerca: Italy, 2012. (In Italian) [Google Scholar]
- De Ranieri, S. Campionamento Biologico delle catture. Sezioni C ed E. Rapporto Finale. In Programma Nazionale Italiano per la Raccolta Dati Alieutici 2011; Consorzio per il Centro Interuniversitario di Biologia Marina ed Ecologia Applicata “G. Bacci”: Italy, 2012. (In Italian) [Google Scholar]
- Minervini, R.; Giannotta, M.; Bianchini, M.L. Observations on the fishery of Rajiformes in Central Tyrrhenian Sea. Oebalia 1985, 11, 583–591. [Google Scholar]
- Abella, A.J.; Auteri, R.; Baino, R.; Lazzaretti, A.; Righini, P.; Serena, F.; Silvestri, R.; Volani, A.; Zucchi, A. Reclutamento di forme giovanili nella fascia costiera toscana (Juveniles recruitment along the Tuscan coastal area). Biol. Mar. Mediterr. 1997, 4, 172–181. (In Italian) [Google Scholar]
- Follesa, M.C.; Addis, P.; Murenu, M.; Saba, R.; Sabatini, A. Annotated check list of the skates (Chondrichthyes, Rajidae) in the Sardinian seas. Biol. Mar. Mediterr. 2003, 10, 828–833. [Google Scholar]
- Catalano, B.; Dalù, M.; Scacco, U.; Vacchi, M. New biological data on Raja brachyura (Chondrichthyes, Rajidae) from around Asinara Island (NW Sardinia, Western Mediterranean). Ital. J. Zool. 2007, 74, 55–61. [Google Scholar] [CrossRef]
- Porcu, C.; Bellodi, A.; Cannas, R.; Marongiu, M.F.; Mulas, A.; Follesa, M.C. Life-history traits of a commercial ray, Raja brachyura from the central western Mediterranean Sea. Med. Mar. Sci. 2014, 16, 90–102. [Google Scholar] [CrossRef] [Green Version]
- Bottari, T.; Rinelli, P.; Bianchini, M.L.; Ragonese, S. Stock identification of Raja clavata L. (Chondrichthyes, Rajidae) in two contiguous areas of the Mediterranean. Hydrobiologia 2013, 703, 215–224. [Google Scholar] [CrossRef]
- Ramírez-Amaro, S.; Ordines, F.; Terrasa, B.; Esteban, A.; García, C.; Guijarro, B.; Massutí, E. Demersal chondrichthyans in the western Mediterranean: Assemblages and biological parameters of their main species. Mar. Freshw. Res. 2016, 67, 636. [Google Scholar] [CrossRef] [Green Version]
- Barone, M. Life History of Raja clavata (Chondrichthyes, Rajiformes). Ph.D. Thesis, Faculty of Mathemathics, Physics and Natural Science of the University of Pisa, Pisa, Italy, 2009. [Google Scholar]
- Saglam, H.; Ak, O. Reproductive biology of Raja clavata (Elasmobranchii: Rajidae) from Southern Black Sea coast around Turkey. Helgol. Mar. Res. 2011, 66, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Bilgin, S.; Onay, H. Spawning Period and Size at Maturity of the Thornback ray, Raja clavata (Linnaeus, 1758), (Elasmobranchii: Rajidae) in the Black Sea. Acta Aquat. Turc. 2020, 16, 525–534. [Google Scholar] [CrossRef]
- Krstulović Šifner, S.; Vrgoč, N.; Dadić, V.; Isajlović, I.; Peharda, M.; Piccinetti, C. Long-term changes in distribution and demographic composition of thornback ray, Raja clavata, in the northern and central Adriatic Sea. J. Appl. Ichthyol. 2009, 25, 40–46. [Google Scholar] [CrossRef]
- Carbonara, P.; Bellodi, A.; Palmisano, M.; Mulas, A.; Porcu, C.; Zupa, W.; Donnaloia, M.; Carlucci, R.; Sion, L.; Follesa, M.C. Growth and Age Validation of the Thornback Ray (Raja clavata Linnaeus, 1758) in the South Adriatic Sea (Central Mediterranean). Front. Mar. Sci. 2020, 7, 586094. [Google Scholar] [CrossRef]
- Ungaro, N. Biological parameters of the brown ray, Raja miraletus, in the Southern Adriatic basin. Cybium 2004, 28, 174–176. [Google Scholar]
- Abdel-Aziz, S.H. The use of vertebral rings of the brown ray Raja miraletus (Linnaeus, 1758) off Egyptian Mediterranean coast for estimation of age and growth. Cybium 1992, 16, 121–132. [Google Scholar]
- Bellodi, A.; Cau, A.; Marongiu, M.F.; Mulas, A.; Porcu, C.; Vittori, S.; Follesa, M.C. Life history parameters of the small Mediterranean-endemic skate, Raja polystigma Regan 1923, from Sardinian seas. In Proceedings of the 5th International Otolith Symposium ICES-CIEM, Mallorca, Spain, 20–24 October 2014; pp. 230–231. [Google Scholar]
- Sin, T.M.; Ang, H.P.; Buurman, J.; Lee, A.C.; Leong, Y.L.; Ooi, S.K.; Steinberg, P.; Teo, S.L. Uncommon biological patterns of a little known endemic Mediterranean skate, Raja polystigma (Risso, 1810). Reg. Stud. Mar. Sci. 2020, 34, 101065. [Google Scholar] [CrossRef] [Green Version]
- Yığın, C.Ç.; Işmen, A. Age, growth and reproduction of the rough ray, Raja radula (Delaroche, 1809) in Saros Bay (North Aegean Sea). J. Black Sea/Mediterr. Environ. 2014, 20, 213–227. [Google Scholar]
- Tiralongo, F.; Messina, G.; Gatti, R.C.; Tibullo, D.; Lombardo, B.M. Some biological aspects of juveniles of the rough ray, Raja radula Delaroche, 1809 in Eastern Sicily (central Mediterranean Sea). J. Sea Res. 2018, 142, 174–179. [Google Scholar] [CrossRef]
- Bombace, G.; Sara, R. La pesca a strascico sui fondali da-500 a-700 metri nel settore a sud-est di Pantelleria. Mem MIN Mar Merc. 1972, 33, 63–77. (In Italian) [Google Scholar]
- Yığın, C.Ç.; Işmen, A. Age, growth, reproduction and feed of bottlenosed skate, Rostroraja alba (Lacepède, 1803) in Saros Bay, the north Aegean Sea. In Proceedings of the ICES Annual Science Conference, Gdańsk, Poland, 20–24 September 2010; pp. 20–24. [Google Scholar]
- Özbek, E.Ö.; Çardak, M.; Kebapçioğlu, T. Spatio-temporal patterns of abundance, biomass and length-weight relationships of Dasyatis species (Pisces: Dasyatidae) in the Gulf of Antalya, Turkey (Levantine Sea). J. Black Sea/Mediterr. Environ. 2015, 21, 169–190. [Google Scholar]
- Girgin, H.; Başusta, N. Testing staining techniques to determine age and growth of Dasyatis pastinaca (Linnaeus, 1758) captured in Iskenderun Bay, northeastern Mediterranean. J. Appl. Ichthyol. 2016, 32, 595–601. [Google Scholar] [CrossRef]
- Serena, F. Field identification guide to the sharks and rays of the Mediterranean and Black Sea. In FAO Species Identification Guide for Fishery Purposes; FAO: Rome, Italy, 2005; 97p. [Google Scholar]
- Yeldan, H.; Avsar, D.; Manaşırlı, M. Age, growth and feeding of the common stingray (Dasyatis pastinaca, L., 1758) in the Cilician coastal basin, northeastern Mediterranean Sea. J. Appl. Ichthyol. 2009, 25, 98–102. [Google Scholar] [CrossRef]
- Yigin, C.C.; Ismen, A. Age, growth and reproduction of the common stingray, Dasyatis pastinaca from the North Aegean Sea. Mar. Biol. Res. 2012, 8, 644–653. [Google Scholar] [CrossRef]
- Ismen, A. Age, growth, reproduction and food of common stingray (Dasyatis pastinaca L., 1758) in İskenderun Bay, the eastern Mediterranean. Fish Res. 2003, 60, 169–176. [Google Scholar] [CrossRef]
- Saadaoui, A.; Saidi, B.; Enajjar, S.; Bradai, M.N. Reproductive biology of the common stingray Dasyatis pastinaca (Linnaeus, 1758) off the Gulf of Gabès (Central Mediterranean Sea). Cah. Biol. Mar. 2015, 56, 389–396. [Google Scholar]
- Başusta, N.; Aslan, E. Age and growth of bull ray Aetomylaeus bovinus (Chondrichthyes: Myliobatidae) from the northeastern Mediterranean coast of Turkey. Cah. Biol. Mar. 2018, 59, 107–114. [Google Scholar] [CrossRef]
- Schwamborn, R. How reliable are the Powell–Wetherall plot method and the maximum-length approach? Implications for length-based studies of growth and mortality. Rev. Fish Biol Fish. 2018, 28, 587–605. [Google Scholar] [CrossRef] [Green Version]
- Russo, T.; Carpentieri, P.; D’Andrea, L.; De Angelis, P.; Fiorentino, F.; Franceschini, S.; Garofalo, G.; Labanchi, L.; Parisi, A.; Scardi, M.; et al. Trends in Effort and Yield of Trawl Fisheries: A Case Study From the Mediterranean Sea. Front. Mar. Sci. 2019, 6, 153. [Google Scholar] [CrossRef]
- Kynoch, R.J.; Fryer, R.J.; Neat, F.C. A simple technical measure to reduce bycatch and discard of skates and sharks in mixed-species bottom-trawl fisheries. ICES J. Mar. Sci. 2015, 72, 1861–1868. [Google Scholar] [CrossRef] [Green Version]
- Chosid, D.M.; Pol, M.; Szymanski, M.; Mirarchi, F.; Mirarchi, A. Development and observations of a spiny dogfish Squalus acanthias reduction device in a raised footrope silver hake Merluccius bilinearis trawl. Fish Res. 2012, 114, 66–75. [Google Scholar] [CrossRef]
- Scannella, D.; Geraci, M.L.; Falsone, F.; Colloca, F.; Zava, B.; Serena, F.; Vitale, S. A new record of a great white shark, Carcharodon carcharias (Chondrichthyes: Lamnidae) in the Strait of Sicily, Central Mediterranean Sea. Acta Adriat. 2020, 61, 181–186. [Google Scholar] [CrossRef]
- ICES. ICES Workshop on Innovative Fishing Gear (WKING). ICES Sci. Rep. 2020, 2, 96. [Google Scholar] [CrossRef]
- Sardo, G.; Okpala, C.O.; Geraci, M.L.; Fiorentino, F.; Vitale, S. The effects of different artificial light wavelengths on some behavioural features of juvenile pelagic atlantic horse mackerel, Trachurus trachurus (actinopterygii: Perciformes: Carangidae). Acta Ichthyol. Piscat. 2020, 50, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Sardo, G.; Geraci, M.L.; Scannella, D.; Falsone, F.; Vitale, S. New records of two uncommon species, Calappa tuerkayana Pastore, 1995 (Decapoda, Calappidae) and Parasquilla ferrussaci (Roux, 1828) (Stomatopoda, Parasquillidae), from the Strait of Sicily (central Mediterranean Sea). Arx. Miscel·Lània Zoològica 2020, 18, 113–121. [Google Scholar] [CrossRef]
- Last, P.R.; White, W.; De Carvalho, M.; Séret, B.; Stehmann, M.; Naylor, G.J.P. Rays of the World; CSIRO Publishing: Clayton, Australia, 2016. [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species, References [Internet]. Electronic Version. 2020. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 10 September 2020).
- Capapé, C.; Guélorget, O.; Vergne, Y.; Quignard, J.P.; Amor, M.M.B.; Bradaï, M.N. Biological observations on the black torpedo, Torpedo nobiliana Bonaparte 1835 (Chondrichthyes: Torpedinidae), from two Mediterranean areas. Ann. Ser. Hist. Nat. 2006, 16, 19. [Google Scholar]
- Barone, M.; De Ranieri, S.; Fabiani, O.; Pirone, A.; Serena, F. Gametogenesis and maturity stages scale of Raja asterias Delaroche, 1809 (Chondrichthyes, Raijdae) from the South Ligurian Sea. Hydrobiologia 2007, 580, 245–254. [Google Scholar] [CrossRef]
- Bakiu, R.; Kolitari, J.; Lleshaj, A. Biological Characteristics and Length-Weight Relationships of landed Thornback Ray (Raja clavata, Linnaeus 1758) in the Fishing Port of Durres, Albania. Albanian J. Agric. Sci. 2021, 20, 14–23. [Google Scholar]
- Capapé, C.; Quignard, J.P. Contribution ò lo biologie des Rajidae. Raja polystigma. Cah. Biol. Mar. 1978, 19, 233–244. [Google Scholar]
- Deval, M.C.; Saygu, İ.; Güven, O.; Özgen, G. Elasmobranch species caught by demersal trawl fisheries in Gulf of Antalya, eastern Mediterranean. In Proceedings of the GFCM First Transversal Expert Meeting on the status of Elasmobranches in the Mediterranean and the Black Sea, Sfax, Tunisia, 20–22 September 2010. [Google Scholar]
- Hemida, F.; Seridji, R.; Ennajar, S.; Bradai, M.N.; Collier, E.; Guelorget, O.; Capape, C. New observations on the reproductive biology of the pelagic stingray, Dasyatis violacea Bonaparte, 1832 (Chondrichthyes: Dasyatidae) from the Mediterranean Sea. Acta Adriat. 2003, 44, 193–204. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Frequency of Occurrence | SSw (t) and SSn (Mean ± SE) | Stratum Distribution (Depth Interval in m) | BI (kg/km2) Mean ± SE | Spearman Coeff. BI | DI (N/km2) Mean ± SE | Spearman Coeff. DI | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| %N. Survey | % Positive Hauls | Tons | Number (Thousand) | 10–50 | 50–100 | 100–200 | 200–500 | 500–800 | Shelf | Slope | Overall | Shelf | Slope | Shelf | Slope | Overall | Shelf | Slope | |
| Tetronarce nobiliana | 76 | 2.1 | 12.5 ± 6.7 | 9 ± 2 |  | | | | <0.1 | 0.7 ± 0.4 | 0.4 ± 0.2 | 0.05 | 0.20 | 0.2 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | −0.03 | −0.10 | |
| Torpedo marmorata | 100 | 8.7 | 20.8 ± 2.5 | 69 ± 7 | | | | | | 1.2 ± 0.1 | 0.2 ± 0.0 | 0.7 ± 0.1 | −0.22 | −0.17 | 3.6 ± 0.4 | 1.0 ± 0.2 | 2.2 ± 0.2 | −0.29 | 0.08 |
| Torpedo torpedo | 40 | 0.8 | 5.8 ± 3.2 | 9 ± 3 | | | | 0.1 ± 0.0 | 0.3 ± 0.2 | 0.2 ± 0.1 | 0.49 | −0.09 | 0.6 ± 0.2 | <0.1 | 0.3 ± 0.1 | 0.49 | −0.09 | ||
| Dipturus batis | 4 | 0.1 | 2.0 ± 2.0 | 0.2 ± 0.2 | | // | 0.1 ± 0.1 | <0.1 | // | NC | // | <0.1 | <0.1 | // | NC | ||||
| Dipturus oxyrinchus | 100 | 10.6 | 92.3 ± 9.4 | 73 ± 7 | | | | <0.1 | 5.5 ± 0.6 | 2.9 ± 0.3 | NC | 0.54 * | <0.1 | 4.3 ± 0.4 | 2.3 ± 0.2 | NC | 0.36 | ||
| Leucoraja circularis | 40 | 0.6 | 2.6 ± 1.0 | 2 ± 1 | | | | <0.1 | 0.2 ± 0.1 | 0.1 ± 0.0 | NC | 0.37 | <0.1 | 0.1 ± 0.0 | 0.1 ± 0.0 | NC | 0.38 | ||
| Leucoraja fullonica | 4 | 0.1 | 0.5 ± 0.5 | 0.1 ± 0.1 | | // | <0.1 | <0.1 | // | NC | // | <0.1 | <0.1 | // | NC | ||||
| Leucoraja melitensis | 96 | 6.9 | 26.7 ± 3.0 | 132 ± 15 | | | | | | 0.4 ± 0.1 | 1.2 ± 0.2 | 0.8 ± 0.1 | 0.54 * | 0.04 | 1.6 ± 0.5 | 6.5 ± 0.8 | 4.2 ± 0.5 | 0.54 * | 0.03 |
| Leucoraja naevus | 4 | 0.2 | 0.1 ± 0.1 | 0.4 ± 0.4 | | // | <0.1 | <0.1 | // | NC | // | <0.1 | <0.1 | // | NC | ||||
| Raja asterias | 96 | 4.3 | 32.7 ± 4.7 | 71 ± 12 | | | | | | 2.1 ± 0.3 | 0.1 ± 0.1 | 1.0 ± 0.1 | −0.37 | 0.24 | 4.4 ± 0.6 | 0.4 ± 0.3 | 2.3 ± 0.4 | −0.29 | 0.24 |
| Raja brachyura | 28 | 0.9 | 9.1 ± 5.6 | 10 ± 6 | | | | 0.3 ± 0.2 | // | 0.1 ± 0.1 | 0.41 | // | 0.3 ± 0.2 | // | 0.1 ± 0.1 | 0.42 | // | ||
| Raja clavata | 100 | 19.8 | 603.6 ± 66.4 | 609 ± 70 | | | | | | 33.3 ± 3.8 | 7.2 ± 1.0 | 19.2 ± 2.1 | 0.86 * | 0.59 * | 31.8 ± 3.6 | 8.8 ± 1.4 | 19.4 ± 2.2 | 0.85 * | 0.66 * |
| Raja miraletus | 100 | 18.9 | 371.9 ± 34.2 | 2282 ± 224 | | | | | | 25.4 ± 2.4 | 0.2 ± 0.1 | 11.8 ± 1.1 | −0.00 | 0.46 | 156.2 ± 15.5 | 1.9 ± 0.4 | 35.2 ± 3.7 | −0.21 | 0.42 |
| Raja montagui | 96 | 6.0 | 19.8 ± 2.6 | 73 ± 13 | | | | | 1.3 ± 0.2 | 0.1 ± 0.0 | 0.6 ± 0.1 | −0.09 | 0.07 | 4.7 ± 0.9 | 0.3 ± 0.1 | 2.3 ± 0.4 | −0.16 | 0.08 | |
| Raja polystigma | 28 | 0.5 | 1.2 ± 0.5 | 4 ± 1 | | | | 0.1 ± 0.0 | <0.1 | <0.1 | 0.45 | 0.49 | 0.2 ± 0.1 | <0.1 | 0.1 ± 0.0 | 0.38 | 0.49 | ||
| Raja radula | 12 | 0.4 | 1.2 ± 0.9 | 1 ± 1 | | 0.1 ± 0.1 | // | <0.1 | 0.07 | // | 0.1 ± 0.1 | // | <0.1 | 0.06 | // | ||||
| Rostroraja alba | 64 | 1.6 | 57.1 ± 15.3 | 11 ± 3 | | | | 3.0 ± 1.1 | 0.8 ± 0.3 | 1.8 ± 0.5 | 0.73 * | 0.13 | 0.6 ± 0.2 | 0.1 ± 0.0 | 0.3 ± 0.1 | 0.69 * | 0.06 | ||
| Dasyatis pastinaca | 64 | 1.2 | 51.7 ± 13.0 | 11 ± 2 | | | | 3.6 ± 0.8 | <0.1 | 1.6 ± 0.4 | 0.73 * | NC | 0.8 ± 0.2 | <0.1 | 0.4 ± 0.1 | 0.67 * | NC | ||
| Pteroplatytrygon violacea | 4 | 0.1 | 1.9 ± 1.9 | 0.2 ± 0.2 | | 0.1 ± 0.1 | // | <0.1 | NC | // | <0.1 | // | <0.1 | NC | // | ||||
| Aetomylaeus bovinus | 12 | 0.4 | 41.7 ± 23.0 | 3 ± 2 | | 2.2 ± 1.6 | // | 1.0 ± 0.7 | 0.01 | // | 0.2 ± 0.1 | // | 0.1 ± 0.1 | −0.02 | // | ||||
| Myliobatis aquila | 44 | 0.7 | 31.0 ± 9.7 | 8 ± 3 | | | 1.0 ± 0.3 | // | 0.5 ± 0.1 | 0.71 * | // | 0.6 ± 0.2 | // | 0.3 ± 0.1 | 0.73 * | // | |||
| Species | Frequency of Occurrence | SSw (t) and SSn (Mean ± SE) | Stratum Distribution (Depth Interval in m) | BI (kg/km2) Mean ± SE | Spearman Coeff. BI | DI (N/km2) Mean ± SE | Spearman Coeff. DI | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % N. Survey | % Positive Hauls | Tons | Number (Thousand) | 50–100 | 100–200 | 200–500 | 500–800 | Shelf | Slope | Overall | Shelf | Slope | Shelf | Slope | Overall | Shelf | Slope | |
| Tetronarce nobiliana | 29 | 1.4 | 1.5 ± 1.1 | 3 ± 1 | | | <0.1 | 0.2 ± 0.2 | 0.1 ± 0.1 | −0.38 | −0.48 | 0.1 ± 0.1 | 0.4 ± 0.2 | 0.3 ± 0.1 | −0.38 | −0.49 | ||
| Torpedo marmorata | 100 | 11.1 | 9.2 ± 2.2 | 35 ± 7 | | | | 1.8 ± 0.5 | 0.1 ± 0.0 | 0.9 ± 0.2 | −0.33 | 0.11 | 6.5 ± 1.3 | 0.7 ± 0.3 | 3.3 ± 0.6 | −0.16 | 0.05 | |
| Dipturus oxyrincus | 100 | 30.94 | 262.4 ± 26.5 | 202 ± 36.8 | | | | 5.1 ± 1.1 | 40.5 ± 5.4 | 24.8 ± 2.5 | 0.15 | −0.15 | 2.5 ± 0.5 | 32.3 ± 4.8 | 19.0 ± 2.9 | 0.53 * | −0.34 | |
| Leucoraja circularis | 79 | 5.7 | 247.5 ± 233.8 | 20 ± 7 | | | | 0.3 ± 0.3 | 41.8 ± 39.8 | 23.4 ± 22.1 | −0.49 | −0.27 | 0.2 ± 0.2 | 3.3 ± 1.2 | 1.9 ± 0.7 | −0.49 | −0.09 | |
| Leucoraja fullonica | 43 | 1.8 | 2.1 ± 1.5 | 3 ± 1 | | | // | 0.4 ± 0.2 | 0.2 ± 0.1 | // | −0.19 | // | 0.5 ± 0.2 | 0.3 ± 0.1 | // | −0.12 | ||
| Leucoraja melitensis | 93 | 10.5 | 20.9 ± 3.8 | 105 ± 19 | | | | | 0.3 ± 0.1 | 3.3 ± 0.6 | 2.0 ± 0.4 | 0.05 | 0.34 | 1.9 ± 1.0 | 16.3 ± 2.9 | 9.9 ± 1.8 | 0.04 | 0.24 |
| Raja asterias | 7 | 0.2 | <0.1 | 1 ± 1 | | // | <0.1 | <0.1 | // | NC | // | <0.1 | <0.1 | // | NC | |||
| Raja brachyura | 14 | 0.3 | 2.2 ± 1.8 | 1 ± 1 | | | 0.4 ± 0.4 | <0.1 | 0.2 ± 0.2 | NC | NC | 0.2 ± 0.2 | <0.1 | 0.1 ± 0.1 | NC | NC | ||
| Raja clavata | 100 | 52.8 | 706.8 ± 61.6 | 660 ± 49 | | | | | 90.7 ± 15.0 | 47.7 ± 5.2 | 66.8 ± 5.8 | 0.80 * | −0.44 | 59.0 ± 7.6 | 65.1 ± 9.1 | 62.4 ± 4.7 | 0.74 * | −0.41 |
| Raja miraletus | 100 | 24.3 | 70.3 ± 8.0 | 277 ± 27 | | | | | 14.1 ± 1.7 | 0.7 ± 0.3 | 6.6 ± 0.8 | 0.26 | −0.29 | 53.8 ± 5.4 | 4.0 ± 2.0 | 26.2 ± 2.6 | −0.13 | −0.23 |
| Raja montagui | 93 | 9.4 | 28.6 ± 8.4 | 103 ± 38 | | | | | 1.5 ± 0.6 | 3.6 ± 1.1 | 2.7 ± 0.8 | −0.02 | 0.83 * | 3.3 ± 1.4 | 14.9 ± 5.8 | 9.7 ± 3.6 | 0.03 | 0.83 * |
| Raja polystigma | 29 | 1.3 | 4.3 ± 2.6 | 5 ± 3 | | | 0.1 ± 0.1 | 0.6 ± 0.4 | 0.4 ± 0.2 | NC | −0.41 | <0.1 | 0.8 ± 0.5 | 0.5 ± 0.3 | NC | −0.44 | ||
| Raja radula | 50 | 1.7 | 22.4 ± 13.6 | 24 ± 13 | | 4.8 ± 2.9 | // | 2.1 ± 1.3 | −0.42 | // | 5.2 ± 2.7 | // | 2.3 ± 1.2 | −0.42 | // | |||
| Rostroraja alba | 7 | 0.3 | 0.4 ± 0.4 | 4 ± 4 | | // | <0.1 | <0.1 | // | NC | <0.1 | 0.7 ± 0.7 | 0.4 ± 0.4 | // | NC | |||
| Dasyatis pastinaca | 71 | 4.3 | 60.2 ± 17.6 | 24 ± 6 | | | | 12.8 ± 3.7 | <0.1 | 5.7 ± 1.7 | −0.97 * | NC | 5.1 ± 1.4 | <0.1 | 2.3 ± 0.6 | −0.78 * | NC | |
| Myliobatis aquila | 64 | 4.6 | 56.5 ± 16.8 | 23 ± 7 | | | 12.0 ± 3.6 | // | 5.3 ± 1.6 | −0.29 | // | 4.8 ± 1.5 | // | 2.1 ± 0.7 | −0.40 | // | ||
| Species | Frequency of Occurrence | SSw (t) and SSn (Mean ± SE) | Stratum Distribution (Depth Interval in m) | BI (kg/km2) Mean ± SE | Spearman Coeff. BI | DI (N/km2) Mean ± SE | Spearman Coeff. DI | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % N. Survey | % Positive Hauls | Tons | Number (Thousand) | 10–50 | 50–100 | 100–200 | 200–500 | 500–800 | Shelf | Slope | Overall | Shelf | Slope | Shelf | Slope | Overall | Shelf | Slope | |
| Tetronarce nobiliana | 40 | 1.2 | 18.7 ± 15.7 | 4 ± 2 | | | | | | <0.1 | 1.1 ± 0.9 | 0.6 ± 0.5 | NC | −0.23 | 0.1 ± 0.1 | 0.2 ± 0.1 | 0.1 ± 0.1 | NC | −0.26 |
| Torpedo marmorata | 80 | 8.2 | 9.2 ± 2.5 | 29 ± 5 | | | | | | 0.6 ± 0.2 | 0.1 ± 0.0 | 0.3 ± 0.1 | −0.39 | 0.44 | 1.6 ± 0.3 | 0.3 ± 0.1 | 0.9 ± 0.2 | −0.26 | 0.49 |
| Torpedo torpedo | 40 | 0.7 | 1.1 ± 0.8 | 3 ± 1 | | | | <0.1 | <0.1 | <0.1 | 0.27 | NC | 0.2 ± 0.1 | <0.1 | 0.1 ± 0.0 | 0.19 | NC | ||
| Dipturus batis | 13 | 0.2 | 1.4 ± 0.9 | 1 ± 1 | | | // | 0.1 ± 0.1 | <0.1 | // | NC | // | <0.1 | <0.1 | // | NC | |||
| Dipturus oxyrinchus | 73 | 6.3 | 47.7 ± 11.0 | 47 ± 19 | | | | <0.1 | 2.8 ± 0.7 | 1.5 ± 0.4 | −0.20 | 0.05 | <0.1 | 2.8 ± 1.1 | 1.5 ± 0.6 | −0.20 | 0.00 | ||
| Leucoraja circularis | 27 | 0.7 | 3.6 ± 1.9 | 2 ± 1 | | // | 0.2 ± 0.1 | 0.1 ± 0.1 | // | 0.35 | // | 0.1 ± 0.1 | 0.1 ± 0.0 | // | 0.34 | ||||
| Leucoraja melitensis | 73 | 4.9 | 14.2 ± 4.0 | 77 ± 25 | | | | | 0.2 ± 0.1 | 0.7 ± 0.2 | 0.5 ± 0.1 | 0.24 | 0.11 | 0.8 ± 0.3 | 3.9 ± 1.4 | 2.5 ± 0.8 | 0.31 | 0.07 | |
| Leucoraja naevus | 7 | 0.1 | <0.1 | 0.1 ± 0.1 | | // | <0.1 | <0.1 | // | NC | // | <0.1 | <0.1 | // | NC | ||||
| Raja asterias | 73 | 2.5 | 4.9 ± 1.4 | 10 ± 2 | | | | 0.3 ± 0.1 | <0.1 | 0.2 ± 0.0 | 0.31 | 0.31 | 0.7 ± 0.2 | <0.1 | 0.3 ± 0.1 | 0.35 | 0.31 | ||
| Raja brachyura | 7 | 0.1 | 0.2 ± 0.2 | 0.2 ± 0.2 | | // | <0.1 | <0.1 | // | 0.36 | // | <0.1 | <0.1 | // | 0.36 | ||||
| Raja clavata | 100 | 18.6 | 304.4 ± 31.8 | 341 ± 51 | | | | | | 16.5 ± 2.3 | 3.8 ± 0.7 | 9.7 ± 1.0 | 0.81 * | −0.15 | 26.2 ± 5.8 | 4.7 ± 1.1 | 14.6 ± 3.0 | 0.86 * | −0.16 |
| Raja miraletus | 100 | 23.2 | 196.2 ± 18.7 | 1211 ± 125 | | | | | 13.3 ± 1.3 | 0.3 ± 0.1 | 6.3 ± 0.6 | 0.67 * | −0.20 | 81.7 ± 8.7 | 1.6 ± 0.8 | 38.6 ± 4.0 | 0.58 * | −0.33 | |
| Raja montagui | 73 | 5.2 | 8.2 ± 1.8 | 30 ± 6 | | | | 0.5 ± 0.1 | 0.1 ± 0.0 | 0.3 ± 0.1 | −0.07 | −0.12 | 1.8 ± 0.4 | 0.2 ± 0.1 | 1.0 ± 0.2 | 0.09 | 0.16 | ||
| Raja polystigma | 7 | 0.1 | <0.1 | 0.1 ± 0.1 | | <0.1 | // | <0.1 | NC | // | <0.1 | // | <0.1 | NC | // | ||||
| Raja radula | 13 | 0.2 | 0.1 ± 0.1 | 1 ± 0.4 | | | <0.1 | // | <0.1 | −0.08 | // | <0.1 | // | <0.1 | −0.08 | // | |||
| Rostroraja alba | 73 | 2.5 | 26.1 ± 9.5 | 9 ± 2 | | | | | 1.6 ± 0.6 | 0.2 ± 0.2 | 0.8 ± 0.3 | −0.11 | −0.41 | 0.6 ± 0.1 | 0.1 ± 0.0 | 0.3 ± 0.1 | 0.04 | −0.39 | |
| Dasyatis pastinaca | 80 | 2.3 | 36.9 ± 9.2 | 9 ± 2 | | | | 2.5 ± 0.6 | // | 1.2 ± 0.3 | 0.36 | // | 0.7 ± 0.2 | // | 0.3 ± 0.1 | 0.36 | // | ||
| Pteroplatytrygon violacea | 7 | 0.1 | 1.5 ± 1.5 | 0.3 ± 0.3 | | 0.1 ± 0.1 | // | <0.1 | NC | // | <0.1 | // | <0.1 | NC | // | ||||
| Aetomylaeus bovinus | 13 | 0.1 | 0.4 ± 0.3 | 0.5 ± 0.3 | | <0.1 | <0.1 | <0.1 | NC | NC | <0.1 | <0.1 | <0.1 | NC | NC | ||||
| Myliobatis aquila | 27 | 0.4 | 1.5 ± 0.9 | 1 ± 1 | | | 0.1 ± 0.1 | // | <0.1 | 0.21 | // | 0.1 ± 0.0 | // | <0.1 | 0.31 | // | |||
| Species | Frequency of Occurrence | SSw (t) and SSn (Mean ± SE) | Stratum Distribution (Depth Interval in m) | BI (kg/km2) Mean ± SE | Spearman Coeff. BI | DI (N/km2) Mean ± SE | Spearman Coeff. DI | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % N. Survey | % Positive hauls | Tons | Number (thousand) | 10–50 | 51–100 | 101–200 | 201–500 | 501–800 | Shelf | Slope | Overall | Shelf | Slope | Shelf | Slope | Overall | Shelf | Slope | |
| Tetronarce nobiliana | 20 | 0.8 | 1.4 ± 0.8 | 2 ± 1 | | | | <0.1 | 0.1 ± 0.0 | <0.1 | NC | NC | <0.1 | 0.1 ± 0.1 | 0.1 ± 0.0 | NC | NC | ||
| Torpedo marmorata | 67 | 10.8 | 9.0 ± 4.0 | 24 ± 7 | | | | | 0.8 ± 0.4 | <0.1 | 0.3 ± 0.1 | 0.1 | 0.71 * | 1.9 ± 0.6 | 0.1 ± 0.1 | 0.8 ± 0.2 | 0.01 | 0.74 * | |
| Torpedo torpedo | 7 | 0.4 | 1.2 ± 1.2 | 1 ± 1 | | | 0.1 ± 0.1 | // | <0.1 | NC | // | 0.1 ± 0.1 | // | <0.1 | NC | // | |||
| Dipturus batis | 7 | 0.6 | 13.8 ± 13.8 | 4 ± 4 | | // | 0.7 ± 0.7 | 0.4 ± 0.4 | // | NC | // | 0.2 ± 0.2 | 0.1 ± 0.1 | // | NC | ||||
| Dipturus oxyrichus | 73 | 19.4 | 283.2 ± 71.5 | 199 ± 43 | | | | 1.6 ± 0.8 | 13.2 ± 3.3 | 9.0 ± 2.3 | 0.75 * | 0.38 * | 0.5 ± 0.2 | 9.7 ± 2.1 | 6.4 ± 1.4 | 0.75 * | 0.22 | ||
| Leucoraja circularis | 40 | 3.1 | 9.7 ± 5.5 | 7 ± 3 | | | // | 0.5 ± 0.3 | 0.3 ± 0.2 | // | 0.41 | // | 0.4 ± 0.1 | 0.2 ± 0.1 | // | 0.25 | |||
| Leucoraja melitensis | 60 | 4.7 | 9.7 ± 4.7 | 51 ± 22 | | | | <0.1 | 0.5 ± 0.2 | 0.3 ± 0.1 | NC | 0.29 | 0.1 ± 0.1 | 2.5 ± 1.1 | 1.6 ± 0.7 | NC | 0.21 | ||
| Raja asterias | 13 | 0.4 | 0.7 ± 0.7 | 1 ± 1 | | | // | <0.1 | <0.1 | // | NC | // | 0.1 ± 0.1 | <0.1 | // | NC | |||
| Raja brachyura | 7 | 0.2 | 0.8 ± 0.8 | 1 ± 1 | | // | <0.1 | <0.1 | // | NC | // | <0.1 | <0.1 | // | NC | ||||
| Raja clavata | 100 | 49 | 521.3 ± 67.3 | 627 ± 93 | | | | | 13.0 ± 3.1 | 18.6 ± 3.2 | 16.6 ± 2.1 | 0.78 * | 0.12 | 8.1 ± 2.2 | 26.6 ± 4.6 | 20.0 ± 2.9 | 0.87 * | 0.09 | |
| Raja miraletus | 93 | 27.6 | 135.8 ± 24.1 | 741 ± 156 | | | | | 11.6 ± 2.1 | 0.2 ± 0.2 | 4.3 ± 0.8 | 0.16 | 0.23 | 63.4 ± 13.9 | 1.3 ± 1.0 | 23.6 ± 5.0 | 0.18 | 0.34 | |
| Raja montagui | 47 | 3.2 | 4.0 ± 1.6 | 13 ± 5 | | | 0.2 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.50 * | −0.21 | 0.3 ± 0.3 | 0.5 ± 0.2 | 0.4 ± 0.2 | 0.50 * | −0.21 | |||
| Raja radula | 16 | 2.4 | 20.5 ± 11.7 | 55 ± 36 | | | | 1.8 ± 1.0 | // | 0.7 ± 0.4 | 0.75 * | // | 4.8 ± 3.2 | // | 1.7 ± 1.1 | 0.75 * | // | ||
| Raja undulata | 7 | 0.1 | 0.2 ± 0.2 | 1 ± 1 | | <0.1 | // | <0.1 | NC | // | 0.1 ± 0.1 | // | <0.1 | NC | // | ||||
| Rostroraja alba | 33 | 1.8 | 7.1 ± 5.4 | 6 ± 3 | | | // | 0.4 ± 0.3 | 0.2 ± 0.2 | // | 0.18 | // | 0.3 ± 0.1 | 0.2 ± 0.1 | // | 0.22 | |||
| Dasyatis pastinaca | 47 | 10.1 | 108.4 ± 59.5 | 43 ± 27 | | | | 9.6 ± 5.3 | // | 3.5 ± 1.9 | 0.56 * | // | 3.8 ± 2.4 | // | 1.4 ± 1.0 | 0.55 * | // | ||
| Myliobatis aquila | 33 | 1.4 | 43.1 ± 23.6 | 12 ± 6 | | | 3.8 ± 2.1 | // | 1.4 ± 0.8 | 0.64 * | // | 1.1 ± 0.6 | // | 0.4 ± 0.2 | 0.67 * | // | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geraci, M.L.; Ragonese, S.; Scannella, D.; Falsone, F.; Gancitano, V.; Mifsud, J.; Gambin, M.; Said, A.; Vitale, S. Batoid Abundances, Spatial Distribution, and Life History Traits in the Strait of Sicily (Central Mediterranean Sea): Bridging a Knowledge Gap through Three Decades of Survey. Animals 2021, 11, 2189. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11082189
Geraci ML, Ragonese S, Scannella D, Falsone F, Gancitano V, Mifsud J, Gambin M, Said A, Vitale S. Batoid Abundances, Spatial Distribution, and Life History Traits in the Strait of Sicily (Central Mediterranean Sea): Bridging a Knowledge Gap through Three Decades of Survey. Animals. 2021; 11(8):2189. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11082189
Chicago/Turabian StyleGeraci, Michele Luca, Sergio Ragonese, Danilo Scannella, Fabio Falsone, Vita Gancitano, Jurgen Mifsud, Miriam Gambin, Alicia Said, and Sergio Vitale. 2021. "Batoid Abundances, Spatial Distribution, and Life History Traits in the Strait of Sicily (Central Mediterranean Sea): Bridging a Knowledge Gap through Three Decades of Survey" Animals 11, no. 8: 2189. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11082189