
Hair Cortisol, Testosterone, Dehydroepiandrosterone Sulfate and Their Ratios in Stallions as a Retrospective Measure of Hypothalamic–Pituitary–Adrenal and Hypothalamic–Pituitary–Gonadal Axes Activity: Exploring the Influence of Seasonality



{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Horses, Housing, Diet, Handling, and Ethical Considerations
2.2. Hair Samples
2.3. Hair Steroid Extraction
2.4. Hormone Detection and Biochemical Validation Tests
2.5. Statistical Analysis
3. Results
3.1. Biochemical Validation of the Enzyme Immunoassay
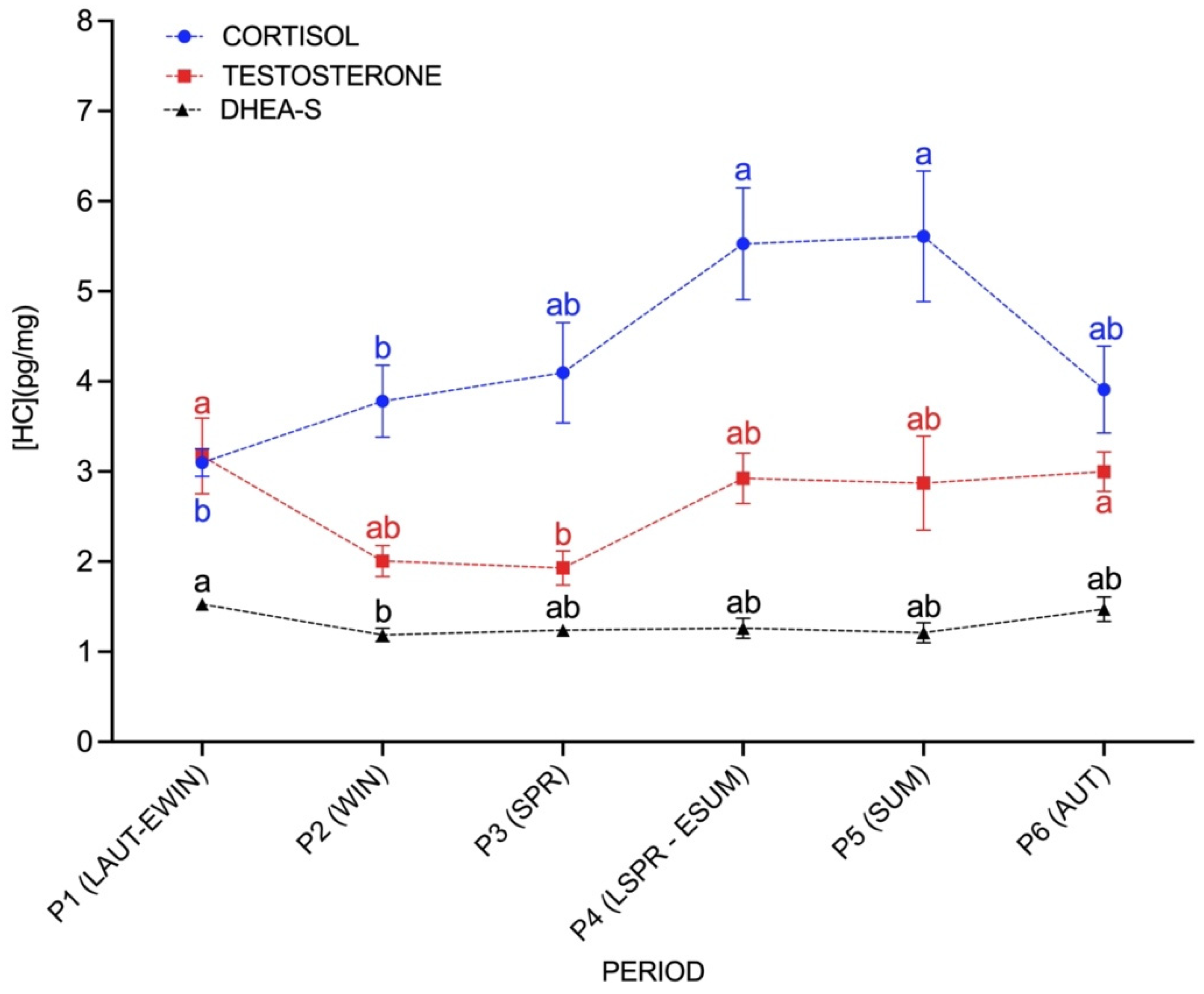
3.2. Seasonal Effect on C, T and DHEA-S Concentrations
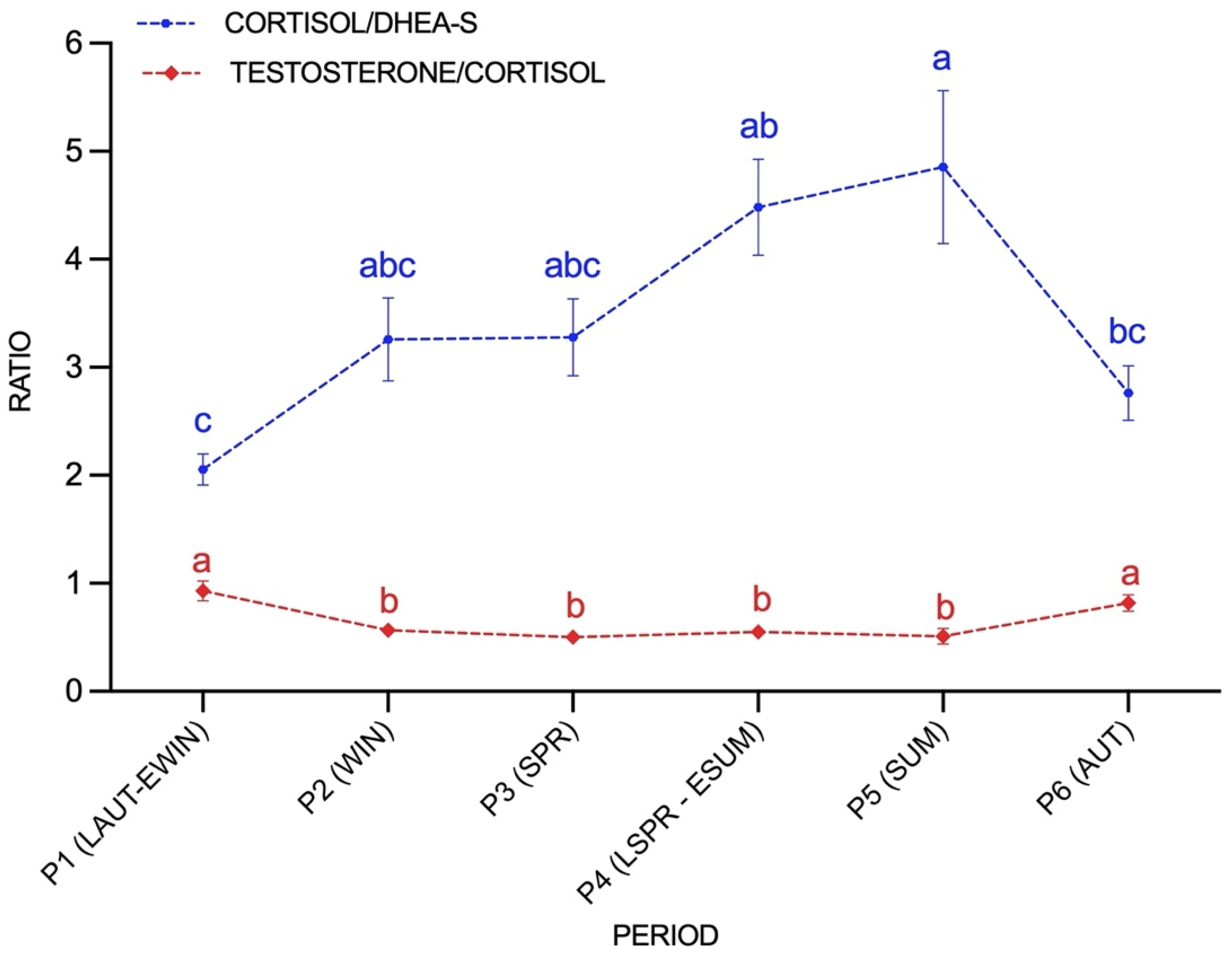
3.3. Seasonal Variations of C/DHEA and T/C Ratios
3.4. Hormone Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murphy, B.A. Circadian and Circannual Regulation in the Horse: Internal Timing in an Elite Athlete. J. Equine Vet. Sci. 2019, 76, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Sollberger, S.; Ehlert, U. How to use and interpret hormone ratios. Psychoneuroendocrinology 2016, 63, 385–397. [Google Scholar] [CrossRef]
- Whitham, J.C.; Bryant, J.L.; Miller, L.J. Beyond Glucocorticoids: Integrating Dehydroepiandrosterone (DHEA) into Animal Welfare Research. Animals 2020, 10, 1381. [Google Scholar] [CrossRef]
- Ralph, C.R.; Tilbrook, A.J. Invited Review: The usefulness of measuring glucocorticoids for assessing animal welfare. J. Anim. Sci. 2016, 94, 457–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheriff, M.J.; Dantzer, B.; Delehanty, B.; Palme, R.; Boonstra, R. Measuring stress in wildlife: Techniques for quantifying glucocorticoids. Oecologia 2011, 166, 869–887. [Google Scholar] [CrossRef] [PubMed]
- Möstl, E.; Palme, R. Hormones as indicators of stress. Domest. Anim. Endocrinol. 2002, 23, 67–74. [Google Scholar] [CrossRef]
- Bergamin, C.; Comin, A.; Corazzin, M.; Faustini, M.; Peric, T.; Scollo, A.; Gottardo, F.; Montillo, M.; Prandi, A. Cortisol, DHEA, and Sexual Steroid Concentrations in Fattening Pigs’ Hair. Animals 2019, 9, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Placci, M.; Marliani, G.; Sabioni, S.; Gabai, G.; Mondo, E.; Borghetti, P.; De Angelis, E.; Accorsi, P.A. Natural Horse Boarding Vs Traditional Stable: A Comparison of Hormonal, Hematological and Immunological Parameters. J. Appl. Anim. Welf. Sci. 2020, 23, 366–377. [Google Scholar] [CrossRef]
- Fels, M.; Rauterberg, S.; Schwennen, C.; Ligges, U.; Herbrandt, S.; Kemper, N.; Schmicke, M. Cortisol/dehydroepiandrosterone ratio in saliva: Endocrine biomarker for chronic stress in pigs? Livest. Sci. 2019, 222, 21–24. [Google Scholar] [CrossRef]
- Peric, T.; Corazzin, M.; Romanzin, A.; Bovolenta, S.; Prandi, A.; Montillo, M.; Comin, A. Cortisol and DHEA concentrations in the hair of dairy cows managed indoor or on pasture. Livest. Sci. 2017, 202, 39–43. [Google Scholar] [CrossRef]
- Khalil, A.M.; Nakahara, K.; Tokuriki, M.; Kaseda, Y.; Murakami, N. Variation in fecal testosterone hormone concentration with season and harem size in Misaki feral horses. J. Vet. Med. Sci. 2009, 71, 1075–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palme, R. Monitoring stress hormone metabolites as a useful, non-invasive tool for welfare assessment in farm animals. Anim. Welf. 2012, 21, 331–337. [Google Scholar] [CrossRef]
- Heimbürge, S.; Kanitz, E.; Otten, W. The use of hair cortisol for the assessment of stress in animals. Gen. Comp. Endocrinol. 2019, 270, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Stalder, T.; Steudte, S.; Miller, R.; Skoluda, N.; Dettenborn, L.; Kirschbaum, C. Intraindividual stability of hair cortisol concentrations. Psychoneuroendocrinology 2012, 37, 602–610. [Google Scholar] [CrossRef]
- Stalder, T.; Kirschbaum, C. Analysis of cortisol in hair - State of the art and future directions. Brain. Behav. Immun. 2012, 26, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Webb, E.; Thomson, S.; Nelson, A.; White, C.; Koren, G.; Rieder, M.; Van Uum, S. Assessing individual systemic stress through cortisol analysis of archaeological hair. J. Archaeol. Sci. 2010, 37, 807–812. [Google Scholar] [CrossRef]
- Duran, M.C.; Janz, D.M.; Waldner, C.L.; Campbell, J.R.; Marques, F.J. Hair Cortisol Concentration as a Stress Biomarker in Horses: Associations With Body Location and Surgical Castration. J. Equine Vet. Sci. 2017, 55, 27–33. [Google Scholar] [CrossRef]
- Heimbürge, S.; Kanitz, E.; Tuchscherer, A.; Otten, W. Within a hair’s breadth – Factors influencing hair cortisol levels in pigs and cattle. Gen. Comp. Endocrinol. 2020, 288, 113359. [Google Scholar] [CrossRef] [PubMed]
- del Rosario González-de-la-Vara, M.; Valdez, R.A.; Lemus-Ramirez, V.; Vázquez-Chagoyán, J.C.; Villa-Godoy, A.; Romano, M.C. Effects of adrenocorticotropic hormone challenge and age on hair cortisol concentrations in dairy cattle. Can. J. Vet. Res. 2011, 75, 216–221. [Google Scholar]
- Tallo-Parra, O.; Manteca, X.; Sabes-Alsina, M.; Carbajal, A.; Lopez-Bejar, M. Hair cortisol detection in dairy cattle by using EIA: Protocol validation and correlation with faecal cortisol metabolites. Animal 2015, 9, 1059–1064. [Google Scholar] [CrossRef] [Green Version]
- Comin, A.; Veronesi, M.C.; Montillo, M.; Faustini, M.; Valentini, S.; Cairoli, F.; Prandi, A. Hair cortisol level as a retrospective marker of hypothalamic-pituitary-adrenal axis activity in horse foals. Vet. J. 2012, 194, 131–132. [Google Scholar] [CrossRef]
- Fourie, N.H.; Brown, J.L.; Jolly, C.J.; Phillips-Conroy, J.E.; Rogers, J.; Bernstein, R.M. Sources of variation in hair cortisol in wild and captive non-human primates. Zoology 2016, 119, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Lafferty, D.J.R.; Laudenslager, M.L.; Mowat, G.; Heard, D.; Belant, J.L. Sex, diet, and the social environment: Factors influencing hair cortisol concentration in free-ranging black bears (Ursus americanus). PLoS ONE 2015, 10, 1–14. [Google Scholar] [CrossRef]
- Bacci, M.L.; Nannoni, E.; Govoni, N.; Scorrano, F.; Zannoni, A.; Forni, M.; Martelli, G.; Sardi, L. Hair cortisol determination in sows in two consecutive reproductive cycles. Reprod. Biol. 2014, 14, 218–223. [Google Scholar] [CrossRef]
- Ventrella, D.; Elmi, A.; Barone, F.; Carnevali, G.; Govoni, N.; Bacci, M.L. Hair testosterone and cortisol concentrations in pre-and post-rut roe deer bucks: Correlations with blood levels and testicular morphometric parameters. Animals 2018, 8, 113. [Google Scholar] [CrossRef] [Green Version]
- Banse, H.E.; Getachew, F.; Levy, M.; Smits, J. Influence of season and pituitary pars intermedia dysfunction on hair cortisol concentration in horses. Domest. Anim. Endocrinol. 2019. [Google Scholar] [CrossRef]
- Gardela, J.; Carbajal, A.; Tallo-Parra, O.; Olvera-Maneu, S.; Álvarez-Rodríguez, M.; Jose-Cunilleras, E.; López-Béjar, M. Temporary Relocation during Rest Periods: Relocation Stress and Other Factors Influence Hair Cortisol Concentrations in Horses. Animals 2020, 10, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munk, R.; Jensen, R.B.; Palme, R.; Christensen, J.W. The effect of sex and time of day on testosterone concentrations in equine saliva and serum. Comp. Exerc. Physiol. 2016, 12, 177–182. [Google Scholar] [CrossRef]
- Waddington, B.; Penitente-Filho, J.M.; Neves, J.G.S.; Pinho, R.O.; Chaya, A.Y.; Maitan, P.P.; Silveira, C.O.; Neves, M.G.; Guimarães, S.E.F.; de Carvalho, G.R.; et al. Testosterone serum profile, semen characteristics and testicular biometry of Mangalarga Marchador stallions in a tropical environment. Reprod. Domest. Anim. 2017, 52, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Aurich, J.; Wulf, M.; Ille, N.; Erber, R.; von Lewinski, M.; Palme, R.; Aurich, C. Effects of season, age, sex, and housing on salivary cortisol concentrations in horses. Domest. Anim. Endocrinol. 2015, 52, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Carroll, C.L.; Huntington, P.J. Body condition scoring and weight estimation of horses. Equine Vet. J. 1988, 20, 41–45. [Google Scholar] [CrossRef]
- Buchanan, K.L.; Goldsmith, A.R. Noninvasive endocrine data for behavioural studies: The importance of validation. Anim. Behav. 2004, 67, 183–185. [Google Scholar] [CrossRef]
- Becker-Birck, M.; Schmidt, A.; Lasarzik, J.; Aurich, J.; Möstl, E.; Aurich, C. Cortisol release and heart rate variability in sport horses participating in equestrian competitions. J. Vet. Behav. Clin. Appl. Res. 2013, 8, 87–94. [Google Scholar] [CrossRef]
- Skoluda, N.; Dettenborn, L.; Stalder, T.; Kirschbaum, C. Elevated hair cortisol concentrations in endurance athletes. Psychoneuroendocrinology 2012, 37, 611–617. [Google Scholar] [CrossRef]
- Cymbaluk, N.F.; Christison, G.I. Environmental effects on thermoregulation and nutrition of horses. Vet. Clin. N. Am. Equine Pract. 1990. [Google Scholar] [CrossRef]
- Montillo, M.; Comin, A.; Corazzin, M.; Peric, T.; Faustini, M.; Veronesi, M.C.; Valentini, S.; Bustaffa, M.; Prandi, A. The Effect of temperature, rainfall, and light conditions on hair cortisol concentrations in newborn foals. J. Equine Vet. Sci. 2014, 34, 774–778. [Google Scholar] [CrossRef]
- Schubach, K.M.; Cooke, R.F.; Brandao, A.P.; Lippolis, K.; Hinchliff, M.T.; Bohnert, D.W.; Cerri, R.L.A. Using hair cortisol concentrations to assess the adrenocortical stress response in beef cattle administered corticotrophin-release hormone. J. Anim. Sci. 2016, 94, 109. [Google Scholar] [CrossRef]
- Hannan, M.A.; Murase, H.; Sato, F.; Tsogtgerel, M.; Kawate, N.; Nambo, Y. Age related and seasonal changes of plasma concentrations of insulin-like peptide 3 and testosterone from birth to early-puberty in Thoroughbred male horses. Theriogenology 2019, 132, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, T.; Aurich, J.E. Regulation of seasonal reproductive activity in the stallion, ram and hamster. Anim. Reprod. Sci. 2000, 58, 197–213. [Google Scholar] [CrossRef]
- Roser, J.F. Endocrine basis for testicular function in the stallion. Theriogenology 1997, 48, 883–892. [Google Scholar] [CrossRef]
- Schmidt, K.; Deichsel, K.; de Oliveira, R.A.; Aurich, J.; Ille, N.; Aurich, C. Effects of environmental temperature and season on hair coat characteristics, physiologic and reproductive parameters in Shetland pony stallions. Theriogenology 2017, 97, 170–178. [Google Scholar] [CrossRef]
- Brinkmann, L.; Gerken, M.; Riek, A. Adaptation strategies to seasonal changes in environmental conditions of a domesticated horse breed, the Shetland pony (Equus ferus caballus). J. Exp. Biol. 2012, 215, 1061–1068. [Google Scholar] [CrossRef] [Green Version]
- Boonstra, R.; Lane, J.E.; Boutin, S.; Bradley, A.; Desantis, L.; Newman, A.E.M.; Soma, K.K. Plasma DHEA levels in wild, territorial red squirrels: Seasonal variation and effect of ACTH. Gen. Comp. Endocrinol. 2008, 158, 61–67. [Google Scholar] [CrossRef]
- Hau, M.; Stoddard, S.T.; Soma, K.K. Territorial aggression and hormones during the non-breeding season in a tropical bird. Horm. Behav. 2004, 45, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Arena, I.; Marliani, G.; Sabioni, S.; Gabai, G.; Bucci, D.; Accorsi, P.A. Assessment of horses’ welfare: Behavioral, hormonal, and husbandry aspects. J. Vet. Behav. 2021, 41, 82–90. [Google Scholar] [CrossRef]
- Almeida, P.E.; Weber, P.S.D.; Burton, J.L.; Zanella, A.J. Depressed DHEA and increased sickness response behaviors in lame dairy cows with inflammatory foot lesions. Domest. Anim. Endocrinol. 2008, 34, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.H.; Josephs, R.A. Testosterone and cortisol jointly regulate dominance: Evidence for a dual-hormone hypothesis. Horm. Behav. 2010, 58, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.H.; Prasad, S. The dual-hormone hypothesis: A brief review and future research agenda. Curr. Opin. Behav. Sci. 2015, 3, 163–168. [Google Scholar] [CrossRef]
- Leary, C.J.; Knapp, R. The stress of elaborate male traits: Integrating glucocorticoids with androgen-based models of sexual selection. Anim. Behav. 2014, 89, 85–92. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olvera-Maneu, S.; Carbajal, A.; Gardela, J.; Lopez-Bejar, M. Hair Cortisol, Testosterone, Dehydroepiandrosterone Sulfate and Their Ratios in Stallions as a Retrospective Measure of Hypothalamic–Pituitary–Adrenal and Hypothalamic–Pituitary–Gonadal Axes Activity: Exploring the Influence of Seasonality. Animals 2021, 11, 2202. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11082202
Olvera-Maneu S, Carbajal A, Gardela J, Lopez-Bejar M. Hair Cortisol, Testosterone, Dehydroepiandrosterone Sulfate and Their Ratios in Stallions as a Retrospective Measure of Hypothalamic–Pituitary–Adrenal and Hypothalamic–Pituitary–Gonadal Axes Activity: Exploring the Influence of Seasonality. Animals. 2021; 11(8):2202. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11082202
Chicago/Turabian StyleOlvera-Maneu, Sergi, Anaïs Carbajal, Jaume Gardela, and Manel Lopez-Bejar. 2021. "Hair Cortisol, Testosterone, Dehydroepiandrosterone Sulfate and Their Ratios in Stallions as a Retrospective Measure of Hypothalamic–Pituitary–Adrenal and Hypothalamic–Pituitary–Gonadal Axes Activity: Exploring the Influence of Seasonality" Animals 11, no. 8: 2202. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11082202