
Group Reunion in Zoo European Wildcats Using Cat Appeasing Pheromone (CAP) and Gradual Release of the Animals in the Exhibit—A Case Study

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
- To advocate the need for systematic publications on the strategies enacted and on the outcomes, both in the short and long term, of as many (re)union episodes as possible in zoos and other facilities housing animals worldwide.
- To describe the behaviour of a family of zoo European wildcats during one such episode, involving a procedure aimed at the reunion of a female cat with her family social group after seven months of absence due to medical reasons.
2. Materials and Methods
2.1. Subjects and Area
2.2. Preamble
2.3. Procedure
2.4. Behavioural Observations
2.5. Data Analysis
- Between the pre-reunion period and the post-reunion period. In this analysis, only data pertaining to the four wildcats who had been in the enclosure in both periods were included (n = 4, RF was excluded).
- Among post-reunion period A, post-reunion period B and the late-reunion period. The data pertaining to all five cats were included in this analysis (n = 5, RF was included).
3. Results
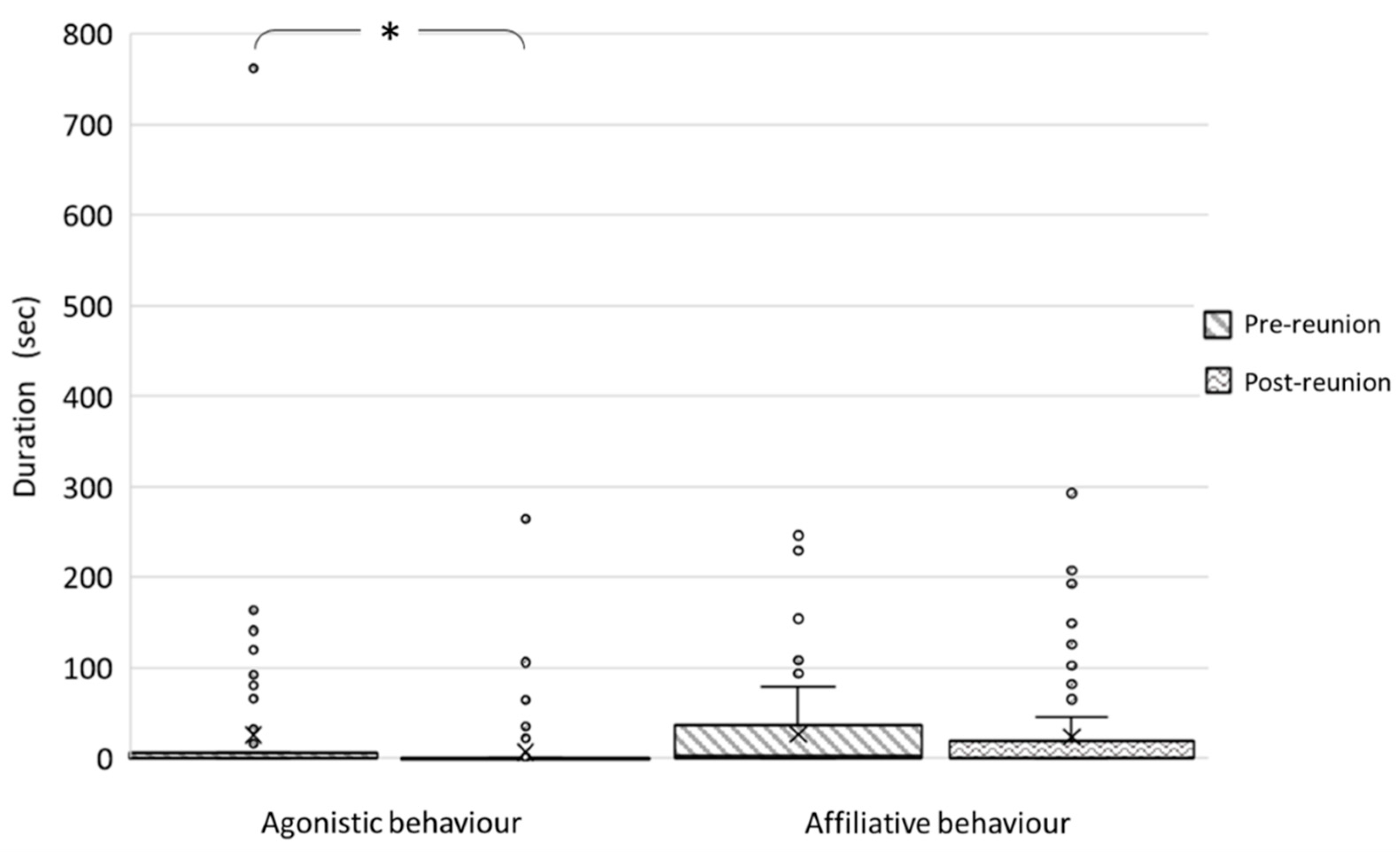
3.1. Pre-Reunion Period vs. Post-Reunion Period
3.1.1. Individual Behaviours, Inactivity and “Not Observed”
3.1.2. Intraspecific and Interspecific Social Behaviour
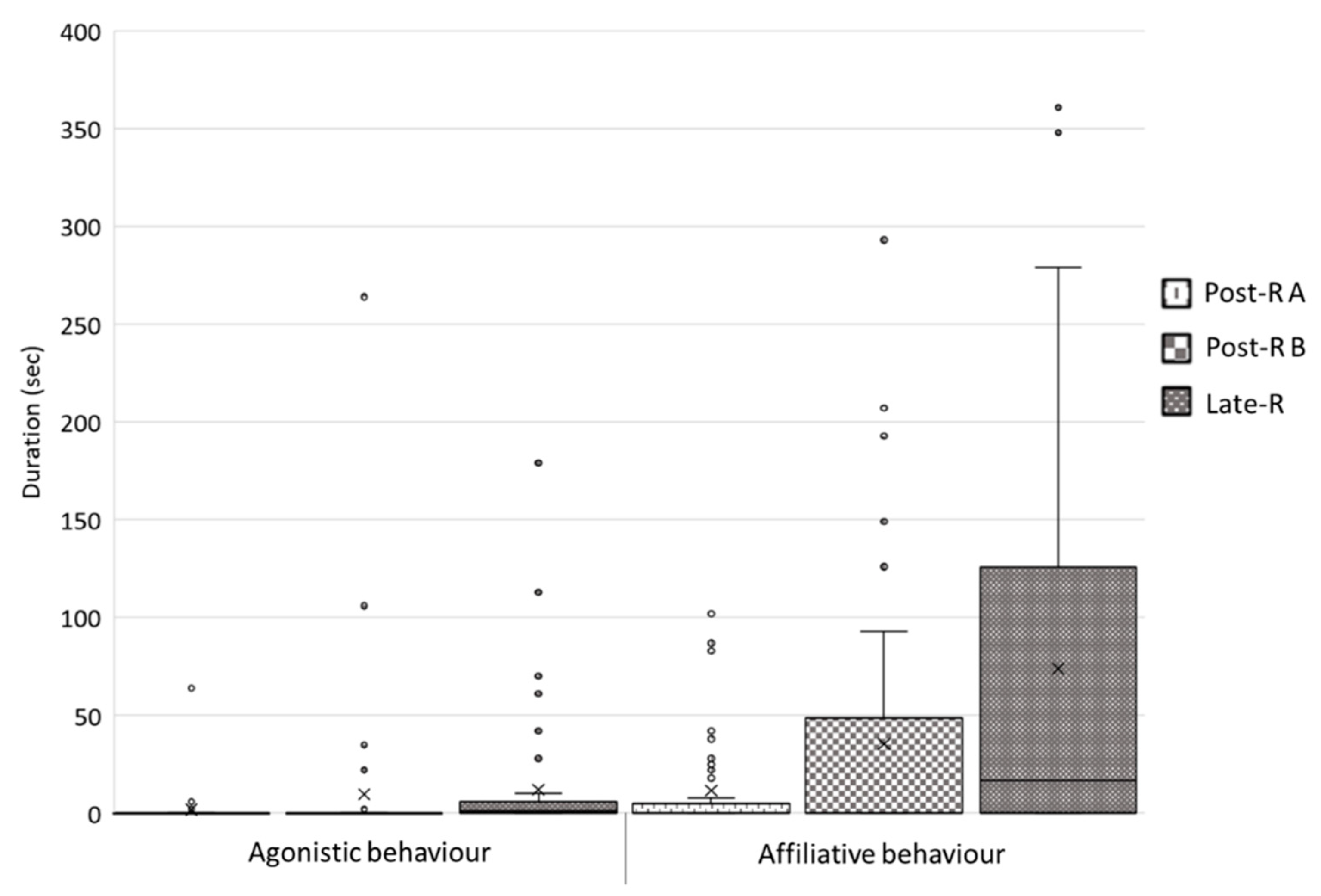
3.2. Post-Reunion Period A vs. Post-Reunion Period B vs. Late-Reunion Period
3.2.1. Individual Behaviours, Inactivity and “Not Observed”
3.2.2. Intraspecific and Interspecific Social Behaviour
Update to Present
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reinhardt, V.; Liss, C.; Stevens, C. Social Housing of Previously Single-Caged Macaques: What are the Options and the Risks? Anim. Welf. 1995, 4, 307–328. [Google Scholar]
- Gaultier, E.; Falewee, C.; Bougrat, L.; Pageat, P. The introduction of a female tiger (Panthera tigris) in a pre-established group of two neutered males: A case study. In Current Issues and Research in Veterinary Behavioural Medicine; Purdue University Press: West Lafayette, IN, USA, 2005; pp. 1–5. [Google Scholar]
- Rox, A.; Kapteijn, C. From solitary to social: The process of resocialisation of rescued primates. Rev. Eubacteria 2012, 30, 1–3. [Google Scholar]
- Moscovice, L.R.; Deschner, T.; Hohmann, G. Welcome Back: Responses of Female Bonobos (Pan paniscus) to Fusions. PLoS ONE 2015, 10, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Den Berghe, F.; Paris, M.C.J.; Sarnyai, Z.; Vlamings, B.; Millar, R.P.; Ganswindt, A.; Cozzi, A.; Pageat, P.; Paris, D.B.B.P. Dog Appeasing Pheromone Prevents the Androgen Surge and May Reduce Contact Dominance and Active Submission After Stressful Interventions in African Wild Dogs (Lycaon pictus). PLoS ONE 2019, 14, e0212551. [Google Scholar] [CrossRef] [PubMed]
- Botreau, R.; Veissier, I.; Butterworth, A.; Bracke, M.B.M.; Keeling, L.J. Definition of Criteria for Overall Assessment of Animal Welfare. Anim. Welf. 2007, 16, 225–228. [Google Scholar]
- Finkle, H.; Terkel, J. Cortisol Levels and Aggression in Neutered and Intact Free-Roaming Female Cats Living in Urban Social Groups. Physiol. Behav. 2010, 99, 343–347. [Google Scholar] [CrossRef]
- Hosey, G.; Melfi, V.; Pankhurst, S. Zoo Animals: Behaviour, Management, and Welfare; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Sackett, D.L.; Rosenberg, W.M.C.; Grey, J.A.M.; Haynes, R.B.; Richardson, W.S. Evidence based medicine: What it is and what it isn’t. BMJ 1996, 312, 71. [Google Scholar] [CrossRef] [Green Version]
- Alligood, N.R.; Lindsay, D.; Mehrkam, R.L.; Leighty, K.A. Applying behavior-analytic methodology to the science and practice of environmental enrichment in zoos and aquariums. Zoo Biol. 2017, 36, 175–185. [Google Scholar] [CrossRef]
- Kroshko, J.; Clubb, R.; Harper, L.; Mellor, E.; Moehrenschlager, A.; Mason, G. Stereotypic route tracing in captive Carnivora is predicted by species—Typical home range sizes and hunting styles. Anim. Behav. 2016, 117, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Gartland, K.; McDonald, M.; Braccini Slade, S.; White, F.; Sanz, C. Behavioral Changes Following Alterations in the Composition of a Captive Bachelor Group of Western Lowland Gorillas (Gorilla gorilla gorilla). Zoo Biol. 2018, 37, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Gusset, M.; Slotow, R.; Somers, M. Divided we Fail: The Importance of Social Integration for the Re-Introduction of Endangered African Wild Dogs (Lycaon pictus). J. Zool. 2006, 270, 502–511. [Google Scholar] [CrossRef]
- Rox, A.; Van Vliet, A.H.; Sterck, E.H.M.; Langermans, J.A.M.; Louwerse, A.L. Factors determining male introduction success and long-term stability in captive rhesus macaques. PLoS ONE 2019, 14, e0219972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hörner, F.; Oerke, S.K.; Müller, D.W.H.; Westerhüs, U.; Azogu-Sepe, I.; Hruby, J.; Preisfeld, G. Monitoring behaviour in African elephants during introduction into a new group: Differences between related and unrelated animals. Animals 2021, 11, 2990. [Google Scholar] [CrossRef] [PubMed]
- De Rouck, M.; Kitchener, A.C.; Law, G.; Nelissen, M. A comparative study of the influence of social housing conditions on the behaviour of captive tigers (Panthera tigris). Anim. Welf. 2005, 14, 229. [Google Scholar]
- Macri, A.M.; Patterson-Kane, E. Behavioural analysis of solitary versus socially housed snow leopards (Panthera uncia), with the provision of simulated social contact. Appl. Anim. Behav. Sci. 2011, 130, 115–123. [Google Scholar] [CrossRef]
- Antonenko, T.V.; Ulitina, O.M.; Pysarev, S.V.; Matsyura, A.V. Different enriched environments for Eurasian lynx in the Barnaul Zoo. Ukr. J. Ecol. 2019, 9, 671–675. [Google Scholar] [CrossRef]
- Alejandro, J.; Yamanashi, Y.; Nemoto, K.; Bercovitch, F.B.; Huffman, M.A. Behavioral changes of solitary housed female pygmy slow lorises (Nycticebus pygmeaus) after introduction into group enclosures. Animals 2021, 11, 2751. [Google Scholar] [CrossRef]
- Szokalski, M.S.; Litchfield, C.A.; Foster, W.K. Enrichment for captive tigers (Panthera tigris): Current knowledge and future directions. Appl. Anim. Behav. Sci. 2012, 139, 1–9. [Google Scholar] [CrossRef]
- Cozzi, A.; Bougrat, L.; Cavicchio, P.; Pageat, P. The use of synthetic cat’s facial pheromones (F3 and F4) to introduce a female and a male tiger in the same enclosure. In Proceedings of the AZA Conference, Phoenix, AZ, USA, 8–13 September 2012. [Google Scholar]
- Wyatt, T.D. Pheromones and Animal Behaviour: Communication by Smell and Taste; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Wyatt, T.D. Pheromones and Animal Behaviour: Chemical Signals and Signatures; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Wyatt, T. Semiochemicals: Pheromones, signature mixtures and behaviour. In Olfaction in Animal Behaviour and Welfare; CAB International: Wallingford, UK, 2017; pp. 26–38. [Google Scholar]
- Pageat, P.; Gaultier, E. Current research in canine and feline pheromones. Vet. Clin. Small Anim. Pract. 2003, 33, 187–211. [Google Scholar] [CrossRef]
- Montague, M.J.; Li, G.; Gandolfi, B.; Khan, R.; Aken, B.L.; Searle, S.M.J.; Minx, P.; Hillier, L.W.; Koboldt, D.C.; Davis, B.W.; et al. Comparative analysis of the domestic cat genome reveals genetic signatures underlying feline biology and domestication. Proc. Natl. Acad. Sci. USA 2014, 111, 17230–17235. [Google Scholar] [CrossRef] [Green Version]
- Wells, D.L. Sensory stimulation as environmental enrichment for captive animals: A review. Appl. Anim. Behav. Sci. 2009, 118, 1–11. [Google Scholar] [CrossRef]
- Dehnhard, M. Mammal semiochemicals: Understanding pheromones and signature mixtures for better zoo-animal husbandry and conservation. Int. Zoo Yearb. 2011, 45, 55–79. [Google Scholar] [CrossRef]
- Nace, A.; Swanson, W.F.; Graham, L.H. Assessing the ability of the synthetic cat pheromone ‘Feliway’ to mitigate post-operative stress in tigers (Panthera tigris). In Campbell Centre for the Study of Animal Welfare Research SympOsium; University of Guelph: Guelph, ON, Canada, 2015. [Google Scholar]
- Spielman, J.S. An Evaluation of the Function of Scent-Marking in Carnivores with a Specific Study into the Effects of Pheromone Enrichment for Captive Tigers (Panthera tigris) and Lions (Panthera leo); University of Edinburgh: Edinburgh, UK, 2000. [Google Scholar]
- Martínez-Macipe, M.; Lafont-Lecuelle, C.; Manteca, X.; Pageat, P.; Cozzi, A. Evaluation of an innovative approach for sensory enrichment in zoos: Semiochemical stimulation for captive lions (Panthera leo). Anim. Welf. 2015, 24, 455–461. [Google Scholar] [CrossRef]
- Costa, A.L.M.D.; Teixeira, R.H.F.; Ribeiro, V.L.; Kokubun, H.S.; Riva, H.G. Uso do feromônio facial felino fração F3 no tratamento de dermatite psicogênica em gatos-maracajá (Leopardus wiedii) cativos-relato de casos. Clín. Vet. 2016, 21, 60–64. [Google Scholar] [CrossRef]
- Pageat, P.; Mengoli, M.; Cozzi, A. Using Feline pheromones: From pet cats to wild Felids. In Proceedings of the International Society of Chemical Ecology Annual Congress, Urbana Champaign, IL, USA, 8–12 July 2014. [Google Scholar]
- Pageat, P. Cat Appeasing Pheromone. Google Patents WO2015/140631 A1, 24 September 2015. [Google Scholar]
- Cozzi, A.; Monneret, P.; Lafont-Lecuelle, C.; Bougrat, L.; Gaultier, E.; Pageat, P. The maternal cat appeasing pheromone: Exploratory study of the effects on aggressive and affiliative interactions in cats. J. Vet. Behav. Clinic. Appl. Res. 2010, 5, 37–38. [Google Scholar] [CrossRef]
- DePorter, T.L.; Bledsoe, D.L.; Beck, A.; Ollivier, E. Evaluation of the efficacy of an appeasing pheromone diffuser product vs placebo for management of feline aggression in multi-cat households: A pilot study. J. Feline Med. Surg. 2018, 21, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, D.W.; Yamaguchi, N.; Kitchener, A.C.; Daniels, M.; Kilshaw, K.; Driscoll, D. The Scottish wildcat: On the way to cryptic extinction through hybridisation: Past history, present problem, and future conservation. In Biology and Conservation of Wild Felids; Macdonald, D.W., Loveridge, A.J., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 471–491. [Google Scholar]
- Lozano, J.; Malo, A.F. Conservation of European Wildcat (Felis silvestris) in Mediterranean Environments: A Reassessment of Current Threats. Mediterranean Ecosystems: Dynamics, Management and Conservation; Nova Science Publishers: Hauppauge, NY, USA, 2012; pp. 1–31. [Google Scholar]
- Yamaguchi, N.; Kitchener, A.; Driscoll, C.; Nussberger, B. Felis Silvestris; The IUCN Red List of Threatened Species: Cambridge, UK, 2015. [Google Scholar]
- Corbett, L.K. Feeding Ecology and Social Organization of Wildcats (Felis silvestris) and Domestic Cats (Felis catus) in Scotland. Ph.D. Thesis, University of Aberdeen, Aberdeen, UK, 1979. [Google Scholar]
- Nowell, K.; Jackson, P. The Wild Cats: Status Survey and Conservation Action Plan; International Union for Nature Conservation—Cat Specialist Group: Gland, Switzerland, 1996. [Google Scholar]
- Sunquist, M.; Sunquist, F. Wild Cats of the World; University of Chicago Press: Chicago, IL, USA, 2002. [Google Scholar]
- Berteselli, G.V.; Regaiolli, B.; Normando, S.; De Mori, B.; Zaborra, C.A.; Spiezio, C. European wildcat and domestic cat: Do they really differ? J. Vet. Behav. 2017, 22, 35–40. [Google Scholar] [CrossRef]
- Stanton, L.A.; Sullivan, M.S.; Fazio, J.M. A standardized ethogram for the Felidae: A tool for behavioural researchers. Appl. Anim. Behav. Sci. 2015, 173, 3–16. [Google Scholar] [CrossRef]
- Altmann, J. Observational study of behaviour: Sampling methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef] [Green Version]
- Regaiolli, B.; Sandri, C.; Rose, P.E.; Vallarin, V.; Spiezio, C. Investigating parental care behaviour in same-sex pairing of zoo greater flamingo (Phoenicopterus roseus). PeerJ 2018, 6, e5227. [Google Scholar] [CrossRef] [Green Version]
- Kratochwill, T.R.; Hitchcock, J.; Horner, R.H.; Levin, J.R.; Odom, S.L.; Rindskopf, D.M.; Shadish, W.R. Single-Case Designs Technical Documentation 2010. Available online: http://ies.ed.gov/ncee/wwc/pdf/wwc_scd.pdf (accessed on 3 June 2016).
- Siegel, S.; Castellan, N.J. Statistica Non Parametrica; McGraw-Hill: New York, NY, USA, 1992. [Google Scholar]
- Nemenyi, P. Distribution-Free Multiple Comparisons. Ph.D. Thesis, Princeton University, Princeton, NJ, USA, 1963. [Google Scholar]
- Pohlert, T. The pairwise multiple comparison of mean ranks package (PMCMR). R Package 2014, 27, 9. [Google Scholar]
- Biró, Z.; Szemethy, L.; Heltai, M. Home range sizes of wildcats (Felis silvestris) and feral domestic cats (Felis silvestris f. catus) in a hilly region of Hungary. Mamm. Biol. 2004, 69, 302–310. [Google Scholar] [CrossRef]
- Elzerman, A.L.; DePorter, T.L.; Beck, A.; Collin, J.F. Conflict and affiliative behavior frequency between cats in multi-cat households: A survey-based study. J. Feline Med. Surg. 2020, 22, 705–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Forkman, B.; Dimitrov, I.; Langbein, J.; et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef]
- Rault, J.L. Friends with benefits: Social support and its relevance for farm animal welfare. Appl. Anim. Behav. Sci. 2012, 136, 1–14. [Google Scholar] [CrossRef]
- Rault, J.L. Be kind to others: Prosocial behaviours and their implications for animal welfare. Appl. Anim. Behav. Sci. 2019, 210, 113–123. [Google Scholar] [CrossRef]
- Clark, F.E.; Fitzpatrick, M.; Hartley, A.; King, A.J.; Lee, T.; Routh, A.; Walker, S.L.; George, K. Relationship between behaviour, adrenal activity, and environment in zoo-housed western lowland gorillas (Gorilla gorilla gorilla). Zoo Biol. 2012, 31, 306–321. [Google Scholar] [CrossRef]
- Larsen, M.J.; Sherwen, S.L.; Rault, J.L. Number of nearby visitors and noise level affect vigilance in captive koalas. Appl. Anim. Behav. Sci. 2014, 154, 76–82. [Google Scholar] [CrossRef]
- Quadros, S.; Goulart, V.D.L.; Passos, L.; Vecci, M.A.M.; Young, R.J. Zoo visitor effect on mammal behaviour: Does noise matter? Appl. Anim. Behav. Sci. 2014, 156, 78–84. [Google Scholar] [CrossRef]
- Sherwen, S.L.; Hemsworth, P.H. The visitor effect on zoo animals: Implications and opportunities for zoo animal welfare. Animals 2019, 9, 366–393. [Google Scholar] [CrossRef] [Green Version]
- Heckstein, R.A.; Hart, B.L. The organization and control of grooming in cats. Appl. Anim. Behav. Sci. 2000, 68, 131–140. [Google Scholar] [CrossRef]
- Regaiolli, B.; Rizzo, A.; Ottolini, G.; Miletto Petrazzini, M.E.; Spiezio, C.; Agrillo, C. Motion illusions as environmental enrichment for zoo animals: A preliminary investigation in lions (Panthera leo). Front. Psychol. 2019, 10, 2220. [Google Scholar] [CrossRef] [Green Version]
- Dawkins, M.S. From an animal’s point of view: Motivation, fitness, and animal welfare. Behav. Brain Sci. 1990, 13, 1–61. [Google Scholar] [CrossRef]
- Van Den Bos, R. Post-conflict stress-response in confined group-living cats (Felis silvestris catus). J. Appl. Anim. Behav. Sci. 1998, 59, 323–330. [Google Scholar] [CrossRef]
- Virga, V. Behavioural dermatology. Vet. Clin. N. Am. Small Anim. Pract. 2003, 33, 231–251. [Google Scholar] [CrossRef]
- Wells, D.L. A note on the influence of visitors on the behaviour and welfare of zoo-housed gorillas. Appl. Anim. Behav. Sci. 2005, 93, 13–17. [Google Scholar] [CrossRef]
- Carlstead, K.; Brown, J.L.; Strawn, W. Behavioural and physiological correlates of stress in laboratory cats. Appl. Anim. Behav. Sci. 1993, 38, 143–158. [Google Scholar] [CrossRef]
- Mallapur, A.; Chellam, R. Environmental influences on stereotypy and the activity budget of Indian leopards (Panthera pardus) in four zoos in Southern India. Zoo Biol. 2002, 21, 585–595. [Google Scholar] [CrossRef]
- Sellinger, R.L.; Ha, J.C. The effects of visitor density and intensity on the behaviour of two captive jaguars (Panthera onca). J. Appl. Anim. Welf. Sci. 2005, 8, 233–244. [Google Scholar] [CrossRef]
- Davey, G. Visitors’ effect on the welfare of animals in the zoo: A review. Appl. Anim. Behav. Sci. 2007, 10, 169–183. [Google Scholar] [CrossRef]
- Morgan, K.N.; Tromborg, C.T. Sources of Stress in Captivity. Appl. Anim. Behav. Sci. 2007, 102, 262–302. [Google Scholar] [CrossRef]
- Powell, D. Preliminary evaluation of environmental enrichment techniques for African lions. Anim. Welf. 1995, 4, 361–370. [Google Scholar]
- Bashaw, M.J.; Bloomsmith, M.A.; Marr, M.; Maple, T.L. To hunt or not to hunt? A feeding enrichment experiment with captive large felids. Zoo Biol. 2003, 22, 189–198. [Google Scholar] [CrossRef]
- Green, T.; Mellor, D. Extending ideas about animal welfare assessment to include ‘quality of life’ and related concepts. N. Z. Vet. J. 2011, 59, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, P.H.; Mellor, D.J.; Cronin, G.M.; Tilbrook, A.J. Scientific assessment of animal welfare. N. Z. Vet. J. 2015, 63, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, J.W.S. Sociality in cats: A comparative review. J. Vet. Behav. 2016, 11, 11–113. [Google Scholar] [CrossRef]
- Sunquist, M. What is a Tiger? Ecology and Behaviour. Tigers of the World, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Normando, S.; Gelli, D.; Meers, L.; Ulitina, O.; Bernardini, B. Study on the Lynx (Lynx lynx & Lynx pardinus) and the European wildcat (Felis silvestris) in Italian zoos. In Proceedings of the XI Convegno Nazionale della Ricerca nei Parchi “Conoscere per proteggere”, Bussolengo, Italy, 1–3 October 2021; p. 36. [Google Scholar]



{kind=link}
{kind=link}
| PR1 | PR2 | PR3 | PR4 | PR5 | PR6 | PR7 | PR8 | PR9 | PR10 | R1 | R2 | POR1 | POR2 | POR3 | POR4 | POR5 | POR6 | POR7 | POR8 | POR9 | POR10 | Pause from 29/07 to 10/09 | LR1 | LR2 | LR3 | LR4 | LR5 | |
| Pre-reunion period | Reunion period | Post-reunion period | Pause 44 days | Late-reunion period | ||||||||||||||||||||||||
| Spot-on | All cats together | Post-reunion period A | Post-reunion period B | |||||||||||||||||||||||||
| Behavioural Categories | Definition |
|---|---|
| Individual behaviours | |
| Attention | A wildcat is alert and stares at a specific point with straight ears or with ears backwards. |
| Maintenance | A wildcat defecates and then covers faeces, urinates, eats and drinks. Includes stretching, body shake, individual play, manipulation of plants and other objects and yawning after waking up. |
| Exploration | A wildcat explores the environment visually or olfactorily (sniffing the ground or any object). |
| Self-grooming | A wildcat cleans its fur by licking, scratching, biting, chewing or by licking a paw and swiping it on the head with the apparent intent to clean the head. |
| Locomotion | A wildcat walks, runs or jumps inside the enclosure. |
| Territorial behaviours | A wildcat marks the environment by urine spray with vertical tail and horizontal urine jet, clawing, rubbing the head against an object, defecating without covering the faeces and patrolling. |
| Social behaviours | |
| Affiliative | A wildcat observes, sniffs or licks another subject or rubs the head and nose against the body of another wildcat. |
| Agonistic | A wildcat stares at another subject, with ears forward of the head. It can move the tail with fast movements or can have ears flat. Includes agonistic displays, such as hissing, piloerection, raised paw and bared teeth. |
| Social interspecific | A wildcat observes zookeepers, visitors or animals belonging to species other than its own. |
| Inactivity | |
| Individual inactivity | A wildcat sleeps or rests alone. |
| Social inactivity | A wildcat sleeps or rests in contact with another subject. |
| Not observed | A wildcat is hiding or is not distinctly visible. |
| Behavioural Categories | Pre-Reunion | Post-Reunion | V-Value | p-Value |
|---|---|---|---|---|
| Individual behaviours | ||||
| Attention | 8 (72.5) | 0 (19) | 3.268 | 0.001 * |
| Maintenance | 6 (18.75) | 0 (13.25) | 1.347 | 0.178 |
| Exploration | 104.5 (399.75) | 12.5 (339.25) | 1.320 | 0.187 |
| Self-grooming | 29 (128) | 0 (64.25) | 2.605 | 0.009 * |
| Locomotion | 0 (33.75) | 0 (22.75) | −0.076 | 0.939 |
| Territorial behaviours | 0 (2.25) | 0 (2.25) | −0.033 | 0.974 |
| Inactivity | ||||
| Individual inactivity | 696.5 (1060.25) | 492.5 (1178) | 0.787 | 0.431 |
| Social inactivity | # | # | # | # |
| Not observed | 0 (4.5) | 0 (126.75) | −2.651 | 0.008 * |
| Periods | Post-Reunion Period A vs. Post-Reunion Period B | Post-Reunion Period A vs. Late-Reunion Period | Post-Reunion Period B vs. Late-Reunion Period | |||
|---|---|---|---|---|---|---|
| Behavioural Categories | Post-Reunion Period A | Post-Reunion Period B | Late-Reunion Period | p-Value | p-Value | p-Value |
| Individual behaviours | ||||||
| Attention | 0 (10.75) | 7.5 (45.25) | 29.5 (94.5) | 0.343 | 0.002 * | 0.114 |
| Maintenance | 0 (6.25) | 3 (18.5) | 7.5 (14.5) | 0.270 | 0.073 | 0.793 |
| Exploration | 9.5 (186.5) | 97(439.75) | 205.5 (438) | 0.011 * | 0.001 * | 0.793 |
| Self-grooming | 0 (15.5) | 0 (61) | 6.5 (57.25) | 0.793 | 0.248 | 0.609 |
| Locomotion | 0 (4.25) | 0 (70.25) | 5.5 (89.25) | 0.189 | 0.082 | 0.916 |
| Territorial behaviour | 0 (0) | 0 (9.5) | 0 (6) | 0.871 | 0.995 | 0.916 |
| Inactivity | ||||||
| Individual inactivity | 1026 (1108.25) | 129.5 (1155.5) | 235 (972.5) | 0.065 | 0.127 | 0.952 |
| Social inactivity | 0 (0) | 0 (0) | 0 (0) | 0.988 | 0.641 | 0.734 |
| Not observed | 0 (79.25) | 12 (124.25) | 0 (66.75) | 0.454 | 0.952 | 0.293 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertoni, V.; Spiezio, C.; Regaiolli, B.; Cozzi, A.; Valsecchi, P.; Normando, S. Group Reunion in Zoo European Wildcats Using Cat Appeasing Pheromone (CAP) and Gradual Release of the Animals in the Exhibit—A Case Study. Animals 2022, 12, 1302. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101302
Bertoni V, Spiezio C, Regaiolli B, Cozzi A, Valsecchi P, Normando S. Group Reunion in Zoo European Wildcats Using Cat Appeasing Pheromone (CAP) and Gradual Release of the Animals in the Exhibit—A Case Study. Animals. 2022; 12(10):1302. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101302
Chicago/Turabian StyleBertoni, Valentina, Caterina Spiezio, Barbara Regaiolli, Alessandro Cozzi, Paola Valsecchi, and Simona Normando. 2022. "Group Reunion in Zoo European Wildcats Using Cat Appeasing Pheromone (CAP) and Gradual Release of the Animals in the Exhibit—A Case Study" Animals 12, no. 10: 1302. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101302