
Effects of Dietary Supplementation with Glycerol Monolaurate (GML) or the Combination of GML and Tributyrin on Growth Performance and Rumen Microbiome of Weaned Lambs

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tested Product
2.2. Experimental Animals and Design
2.3. Diet Sampling and Analysis
2.4. Determination of Apparent Digestibility
2.5. Rumen Fluid Sampling and Analysis
2.6. DNA Preparation and Sequencing
2.7. Statistical Analyses
3. Results
3.1. Growth Performance and Apparent Digestibility
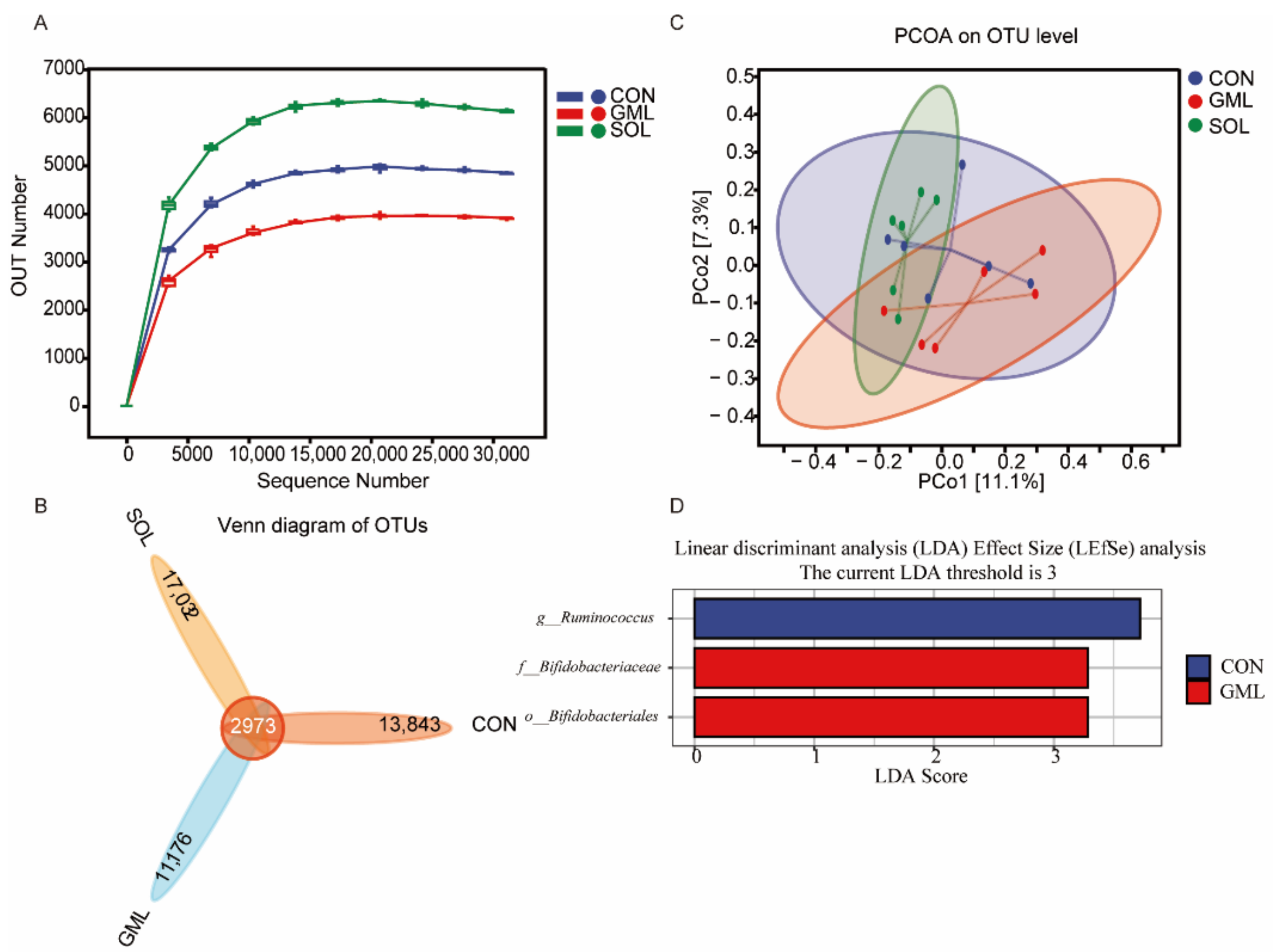
3.2. Rumen Bacterial Diversity
3.3. Rumen Bacterial Composition
4. Discussion
4.1. The Effect of Glycerol Monolaurate (GML) or the Combination (Solider, SOL) of GML and Tributyrin (TB) on the Growth Performance of Weaned Lambs
4.2. The Effect of Glycerol Monolaurate (GML) or the Combination (Solider, SOL) of GML and Tributyrin (TB) on the Rumen Microbiome of Weaned Lambs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ekanayake, L.J.; Corner-Thomas, R.A.; Cranston, L.M.; Kenyon, P.R.; Morris, S.T.; Pain, S.J. Pre-Exposure of Early-Weaned Lambs to a Herb-Clover Mix Does Not Improve Their Subsequent Growth. Animals 2020, 10, 1354. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Castellano, L.E.; Suarez-Trujillo, A.; Martell-Jaizme, D.; Cugno, G.; Arguello, A.; Castro, N. The effect of 357 colostrum period management on BW and immune system in lambs: From birth to weaning. Animal 2015, 9, 1672–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekiz, B.; Kocak, O.; Yalcintan, H.; Yilmaz, A. Effects of suckling duration on growth, slaughtering and carcass quality characteristics of Kivircik lambs. Trop. Anim. Health Prod. 2016, 48, 48–395. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.M.; Mao, S.Y.; Zhu, W.Y.; Liu, J.H. Effect of starter diet supplementation on rumen epithelial morphology and expression of genes involved in cell proliferation and metabolism in pre-weaned lambs. Animal 2018, 12, 2274–2283. [Google Scholar] [CrossRef]
- Li, C.; Zhang, Q.; Wang, G.; Niu, X.; Wang, W.; Li, F.; Li, F.; Zhang, Z. The functional development of the rumen is influenced by weaning and associated with ruminal microbiota in lambs. Anim. Biotechnol. 2020, 8, 1–17. [Google Scholar] [CrossRef]
- Welch, J.L.; Xiang, J.H.; Okeoma, C.M.; Schlievert, P.M.; Stapleton, J.T. Glycerol Monolaurate, an Analogue to a Factor Secreted by Lactobacillus, Is Virucidal against Enveloped Viruses, Including HIV-1. Mbio 2020, 11, e00686-20. [Google Scholar] [CrossRef]
- Wieland, M.; Weber, B.K.; Hafner-Marx, A.; Sauter-Louis, C.; Bauer, J.; Knubben-Schweizer, G.; Metzner, M. A controlled trial on the effect of feeding dietary chestnut extract and glycerol monolaurate on liver function in newborn calves. J. Anim. Physiol. Anim. Nutr. 2015, 99, 190–200. [Google Scholar] [CrossRef]
- Jackman, J.A.; Hakobyan, A.; Zakaryan, H.; Elrod, C.C. Inhibition of African swine fever virus in liquid and feed by medium-chain fatty acids and glycerol monolaurate. J. Anim. Sci. Biotechnol. 2020, 11, 114. [Google Scholar] [CrossRef]
- Valentini, J.; Da Silva, A.S.; Fortuoso, B.F.; Reis, J.H.; Gebert, R.R.; Griss, L.G.; Boiago, M.M.; Lopes, L.Q.S.; Santos, R.C.V.; Wagner, R.; et al. Chemical composition, lipid peroxidation, and fatty acid profile in meat of broilers fed with glycerol monolaurate additive. Food Chem. 2020, 330, 127187. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, H.; Alhamoud, Y.; Chen, Y.; Zhuang, J.; Liu, T.; Cai, L.; Shen, W.; Wu, X.; Zheng, W.; et al. Integrated metabolomic and gene expression analyses to study the effects of glycerol monolaurate on flesh quality in large yellow croaker (Larimichthys crocea). Food Chem. 2022, 367, 130749. [Google Scholar] [CrossRef]
- Fortuoso, B.F.; Dos Reis, J.H.; Gebert, R.R.; Barreta, M.; Griss, L.G.; Casagrande, R.A.; de Cristo, T.G.; Santiani, F.; Campigotto, G.; Rampazzo, L.; et al. Glycerol monolaurate in the diet of broiler chickens replacing conventional antimicrobials: Impact on health, performance and meat quality. Microb. Pathog. 2019, 129, 161–167. [Google Scholar] [CrossRef]
- Zhang, W.X.; Zhang, Y.; Zhang, X.W.; Deng, Z.X.; Liu, J.X.; He, M.L.; Wang, H.F. Effects of Dietary Supplementation with Combination of Tributyrin and Essential Oil on Gut Health and Microbiota of Weaned Piglets. Animals 2020, 10, 180. [Google Scholar] [CrossRef] [Green Version]
- Fortuoso, B.F.; Galli, G.M.; Griss, L.G.; Armanini, E.H.; Silva, A.D.; Fracasso, M.; Mostardeiro, V.; Morsch, V.M.; Lopes, L.Q.S.; Santos, R.C.V.; et al. Effects of glycerol monolaurate on growth and physiology of chicks consuming diet containing fumonisin. Microb. Pathog. 2020, 147, 104261. [Google Scholar] [CrossRef]
- Chan, A.; Lensing, A.W.A.; Kubitza, D.; Brown, G.; Elorza, D.; Ybarra, M.; Halton, J.; Grunt, S.; Kenet, G.; Bonnet, D.; et al. Clinical presentation and therapeutic management of venous thrombosis in young children: A retrospective analysis. Thromb. J. 2018, 16, 29. [Google Scholar] [CrossRef] [Green Version]
- Schlievert, P.M.; Kilgore, S.H.; Kaus, G.M.; Ho, T.D.; Ellermeier, C.D. Glycerol Monolaurate (GML) and a Nonaqueous Five-Percent GML Gel Kill Bacillus and Clostridium Spores. mSphere 2018, 3, e00597-18. [Google Scholar] [CrossRef] [Green Version]
- Strandberg, K.L.; Peterson, M.L.; Lin, Y.C.; Pack, M.C.; Chase, D.J.; Schlievert, P.M. Glycerol monolaurate inhibits Candida and Gardnerella vaginalis in vitro and in vivo but not Lactobacillus. Antimicrob. Agents Chemother. 2010, 54, 597–601. [Google Scholar] [CrossRef] [Green Version]
- Ren, Q.C.; Xuan, J.J.; Wang, L.K.; Zhan, Q.W.; Yin, D.Z.; Hu, Z.Z.; Yang, H.J.; Zhang, W.; Jiang, L.S. Effects of tributyrin supplementation on ruminal microbial protein yield, fermentation characteristics and nutrients degradability in adult Small Tail ewes. Anim. Sci. J. 2018, 89, 1271–1279. [Google Scholar] [CrossRef]
- Vandermeers, A.; Vandermeers-Piret, M.C.; Rathe, J.; Christophe, J. Competitive inhibitory effect exerted by bile salt micelles on the hydrolysis of tributyrin by pancreatic lipase. Biochem. Biophys. Res. Commun. 1976, 69, 790–797. [Google Scholar] [CrossRef]
- Dell’Anno, M.; Reggi, S.; Caprarulo, V.; Hejna, M.; Sgoifo Rossi, C.A.; Callegari, M.L.; Baldi, A.; Rossi, L. Evaluation of 405 Tannin Extracts, Leonardite and Tributyrin Supplementation on Diarrhoea Incidence and Gut Microbiota of Weaned Piglets. Animals 2021, 11, 1693. [Google Scholar] [CrossRef]
- He, J.; Dong, L.; Xu, W.; Bai, K.; Lu, C.; Wu, Y.; Huang, Q.; Zhang, L.; Wang, T. Dietary Tributyrin Supplementation Attenuates Insulin Resistance and Abnormal Lipid Metabolism in Suckling Piglets with Intrauterine Growth Retardation. PLoS ONE 2015, 10, e0136848. [Google Scholar] [CrossRef]
- Guo, W.; Liu, J.; Yang, Y.; Ma, H.; Gong, Q.; Kan, X.; Ran, X.; Cao, Y.; Wang, J.; Fu, S.; et al. Rumen-bypassed tributyrin alleviates heat stress by reducing the inflammatory responses of immune cells. Poult. Sci. 2021, 100, 348–356. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis, 15th ed.; AOAS International Press: Arlington, VA, USA, 2011. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Zhao, Y.; Tang, Z.; Nan, X.; Sun, F.; Jiang, L.; Xiong, B. Effects of Lonicera japonica extract on performance, blood biomarkers of inflammation and oxidative stress during perinatal period in dairy cows. Asian-Australas. J. Anim. Sci. 2020, 33, 1096–1102. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.; Garcia-Santos, S.; Nunes, A.; Rito, S.; Azevedo, J.; Guedes, C.; Silva, S.; Ferreira, L. Introducing Mediterranean Lupins in Lambs’ Diets: Effects on Growth and Digestibility. Animals 2021, 11, 942. [Google Scholar] [CrossRef]
- Sun, Z.; Yu, Z.; Wang, B. Perilla frutescens Leaf Alters the Rumen Microbial Community of Lactating Dairy Cows. Microorganisms 2019, 7, 562. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Gallardo Paffetti, M.; Carcamo, J.; Arias-Darraz, L.; Alvear, C.; Ojeda, J. Effect of Type of Pregnancy on Transcriptional and Plasma Metabolic Response in Sheep and Its Further Effect on Progeny Lambs. Animals 2020, 10, 2290. [Google Scholar] [CrossRef]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Liu, C.; Cui, Y.; Li, X.; Yao, M. microeco: An R package for data mining in microbial community ecology. FEMS Microbiol. Ecol. 2021, 97, fiaa255. [Google Scholar] [CrossRef]
- Ragionieri, L.; Cacchioli, A.; Ravanetti, F.; Botti, M.; Ivanovska, A.; Panu, R.; Righi, F.; Quarantelli, A.; Gazza, F. Effect of the supplementation with a blend containing short and medium chain fatty acid monoglycerides in milk replacer on rumen papillae development in weaning calves. Ann. Anat. 2016, 207, 97–108. [Google Scholar] [CrossRef]
- Liu, T.; Tang, J.; Feng, F. Glycerol monolaurate improves performance, intestinal development, and muscle amino acids in yellow-feathered broilers via manipulating gut microbiota. Appl. Microbiol. Biotechnol. 2020, 104, 10279–10291. [Google Scholar] [CrossRef] [PubMed]
- Machmuller, A.; Soliva, C.R.; Kreuzer, M. Effect of coconut oil and defaunation treatment on methanogenesis in sheep. Reprod. Nutr. Dev. 2003, 43, 41–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reveneau, C.; Ribeiro, C.V.; Eastridge, M.L.; Firkins, J.L. Interaction of unsaturated fat or coconut oil with monensin in lactating dairy cows fed 12 times daily. II. Fatty acid flow to the omasum and milk fatty acid profile. J. Dairy Sci. 2012, 95, 2061–2069. [Google Scholar] [CrossRef] [PubMed]
- Panyakaew, P.; Boon, N.; Goel, G.; Yuangklang, C.; Schonewille, J.T.; Hendriks, W.H.; Fievez, V. Effect of supplementing coconut or krabok oil, rich in medium-chain fatty acids on ruminal fermentation, protozoa and archaeal population of bulls. Animal 2013, 7, 1950–1958. [Google Scholar] [CrossRef] [Green Version]
- Faciola, A.P.; Broderick, G.A. Effects of feeding lauric acid or coconut oil on ruminal protozoa numbers, fermentation pattern, digestion, omasal nutrient flow, and milk production in dairy cows. J. Dairy Sci. 2014, 97, 5088–5100. [Google Scholar] [CrossRef]
- Hu, Q.; Yin, F.; Li, B.; Guo, Y.; Yin, Y. Dietary Tributyrin Administration Improves Intestinal Morphology and Selected Bacterial and Short-Chain Fatty Acid Profiles in Broilers Under an Isocaloric Feeding Regime. Front. Microbiol. 2021, 12, 715712. [Google Scholar] [CrossRef]
- Liu, S.; Ma, J.Y.; Zhou, J.; Wu, J.D.; Li, J.H.; Alugongo, G.M.; Xiao, J.X.; Wang, J.J.; Wang, Y.J.; Wang, W.; et al. Tributyrin supplementation in pasteurized waste milk: Effects on growth performance, health, and blood parameters of dairy calves. J. Dairy Sci. 2021, 104, 12496–12507. [Google Scholar] [CrossRef]
- Shen, H.; Lu, Z.; Chen, Z.; Wu, Y.; Shen, Z. Rapid Fermentable Substance Modulates Interactions between Ruminal Commensals and Toll-Like Receptors in Promotion of Immune Tolerance of Goat Rumen. Front. Microbiol. 2016, 7, 1812. [Google Scholar] [CrossRef]
- Jiang, C.X.; Ding, L.M.; Dong, Q.M.; Wang, X.J.; Wei, H.Y.; Hu, C.S.; Ma, C.F.; Yan, Q.; Zhou, Y.Q.; Degen, A.A. Effects of root extracts of three traditional Chinese herbs as dietary supplements on dry matter intake, average daily gain, rumen fermentation and ruminal microbiota in early weaned yak calves. Anim. Feed Sci. Technol. 2021, 278, 115002. [Google Scholar] [CrossRef]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J.P.; Klenk, H.P.; Clement, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Sulak, M.; Sikorova, L.; Jankuvova, J.; Javorsky, P.; Pristas, P. Variability of Actinobacteria, a minor component of rumen microflora. Folia Microbiol. 2012, 57, 351–353. [Google Scholar] [CrossRef]
- Kovanda, L.; Zhang, W.; Wei, X.; Luo, J.; Wu, X.; Atwill, E.R.; Vaessen, S.; Li, X.; Liu, Y. In Vitro Antimicrobial Activities of Organic Acids and Their Derivatives on Several Species of Gram-Negative and Gram-Positive Bacteria. Molecules 2019, 24, 3770. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wang, R.; Zhang, G.; Chen, F.; Xu, B. In Vitro Antibacterial Activities and Mechanisms of Action of Fatty Acid Monoglycerides against Four Foodborne Bacteria. J. Food Prot. 2020, 83, 331–337. [Google Scholar] [CrossRef]
- Neubauer, V.; Petri, R.; Humer, E.; Kroger, I.; Mann, E.; Reisinger, N.; Wagner, M.; Zebeli, Q. High-grain diets supplemented with phytogenic compounds or autolyzed yeast modulate ruminal bacterial community and fermentation in dry cows. J. Dairy Sci. 2018, 101, 2335–2349. [Google Scholar] [CrossRef]
- Won, M.Y.; Oyama, L.B.; Courtney, S.J.; Creevey, C.J.; Huws, S.A. Can rumen bacteria communicate to each other? Microbiome 2020, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Wang, C.; Mao, H. Effects of leucine addition on the rumen development and rumen bacterial communities of early weaned Hu lambs. Chin. J. Anim. Sci. 2019, 55, 66–71. [Google Scholar]
- Jung, D.H.; Seo, D.H.; Kim, G.Y.; Nam, Y.D.; Song, E.J.; Yoon, S.; Park, C.S. The effect of resistant starch (RS) on the bovine rumen microflora and isolation of RS-degrading bacteria. Appl. Microbiol. Biotechnol. 2018, 102, 4927–4936. [Google Scholar] [CrossRef]


{kind=link}
| Item | Content (%) |
|---|---|
| Ingredients | |
| Silage corn | 15.00 |
| Alfalfa hay | 25.00 |
| Corn | 28.80 |
| Soybean meal | 16.50 |
| Rapeseed meal | 9.50 |
| Wheat bran | 3.00 |
| CaHPO4 | 0.50 |
| Limestone | 0.40 |
| NaCl | 0.30 |
| Premix 1 | 1.00 |
| Total | 100.00 |
| Nutrient levels 2 | |
| ME (MJ/kg) | 11.12 |
| CP | 17.46 |
| EE | 2.32 |
| NDF | 24.56 |
| ADF | 10.45 |
| Ca | 0.66 |
| P | 0.53 |
| Item | Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | GML | SOL | |||
| Initial BW (kg) | 11.56 | 11.11 | 11.70 | 0.23 | 0.28 |
| Final BW (kg) | 20.01 b | 22.26 a | 21.36 ab | 0.60 | 0.04 |
| ADG (kg) | 0.19 b | 0.24 a | 0.22 a | 7.99 | 0.02 |
| DMI (kg/d) | 0.68 | 0.72 | 0.71 | 11.05 | 0.13 |
| F/G | 3.59 x | 3.05 y | 3.21 xy | 0.11 | 0.07 |
| Item | Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | GML | SOL | |||
| DM | 81.73 | 82.60 | 83.35 | 0.45 | 0.37 |
| OM | 82.62 | 84.62 | 83.71 | 0.61 | 0.43 |
| CP | 81.03 y | 89.04 x | 85.13 xy | 1.48 | 0.08 |
| EE | 89.15 | 90.01 | 89.23 | 1.21 | 0.96 |
| NDF | 51.43 b | 55.07 ab | 56.43 a | 0.87 | 0.04 |
| ADF | 49.85 | 52.15 | 53.33 | 0.98 | 0.36 |
| Item | Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | GML | SOL | |||
| Chao1 | 4840.52 b | 3895.97 b | 6123.24 a | 312.45 | 0.01 |
| Shannon | 9.70 ab | 8.96 b | 10.42 a | 0.21 | 0.01 |
| Simpson | 0.99 | 0.98 | 0.99 | 0.01 | 0.15 |
| Item | Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | GML | SOL | |||
| Bacteroidetes | 62.91 | 71.96 | 67.62 | 0.03 | 0.38 |
| Firmicutes | 33.99 | 24.59 | 28.67 | 0.03 | 0.34 |
| Tenericutes | 0.70 | 0.57 | 0.75 | <0.01 | 0.82 |
| Proteobacteria | 0.99 | 0.98 | 0.99 | <0.01 | 0.62 |
| Spirochaetes | 0.33 | 0.54 | 0.59 | <0.01 | 0.76 |
| Actinobacteria | 0.26 ab | 0.56 a | 0.12 b | <0.01 | 0.03 |
| Synergistetes | 0.33 | 0.09 | 0.37 | <0.01 | 0.65 |
| Fibrobacteres | 0.11 | 0.17 | 0.26 | <0.01 | 0.12 |
| TM7 | 0.12 | 0.02 | 0.05 | <0.01 | 0.17 |
| Verrucomicrobia | 0.02 b | 0.03 ab | 0.07 a | <0.01 | 0.05 |
| Item | Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | GML | SOL | |||
| Prevotella | 37.54 | 40.37 | 38.29 | 0.04 | 0.95 |
| Succiniclasticum | 2.99 | 2.96 | 3.66 | <0.01 | 0.44 |
| Ruminococcus | 2.10 a | 1.11 b | 1.82 ab | <0.01 | 0.03 |
| Oscillospira | 1.41 | 1.65 | 0.58 | <0.01 | 0.37 |
| Shuttleworthia | 0.25 | 0.04 | 2.17 | <0.01 | 0.30 |
| Butyrivibrio | 0.44 | 0.62 | 1.23 | <0.01 | 0.16 |
| RFN20 | 0.47 | 0.52 | 0.92 | <0.01 | 0.48 |
| CF231 | 0.88 | 0.34 | 0.44 | <0.01 | 0.22 |
| Anaerovibrio | 0.70 | 0.49 | 0.35 | <0.01 | 0.49 |
| Anaeroplasma | 0.50 | 0.43 | 0.53 | <0.01 | 0.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Wang, H.; Zhang, Y.; Li, X.; Jiang, X.; Ding, H. Effects of Dietary Supplementation with Glycerol Monolaurate (GML) or the Combination of GML and Tributyrin on Growth Performance and Rumen Microbiome of Weaned Lambs. Animals 2022, 12, 1309. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101309
Li Y, Wang H, Zhang Y, Li X, Jiang X, Ding H. Effects of Dietary Supplementation with Glycerol Monolaurate (GML) or the Combination of GML and Tributyrin on Growth Performance and Rumen Microbiome of Weaned Lambs. Animals. 2022; 12(10):1309. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101309
Chicago/Turabian StyleLi, Yi, Heze Wang, Yulei Zhang, Xilong Li, Xianren Jiang, and Hongbiao Ding. 2022. "Effects of Dietary Supplementation with Glycerol Monolaurate (GML) or the Combination of GML and Tributyrin on Growth Performance and Rumen Microbiome of Weaned Lambs" Animals 12, no. 10: 1309. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101309