
Delay of Feed Post-Hatch Causes Changes in Expression of Immune-Related Genes and Their Correlation with Components of Gut Microbiota, but Does Not Affect Protein Expression

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Husbandry Protocols
2.2. Bird Sampling
2.3. RNA Isolation and Reverse Transcription-Quantitative PCR
2.4. Protein Extraction and ELISA
2.5. DNA Isolation, Library Preparation, and Microbiota Analysis
2.6. Statistical Analysis
3. Results
3.1. Gene Expression
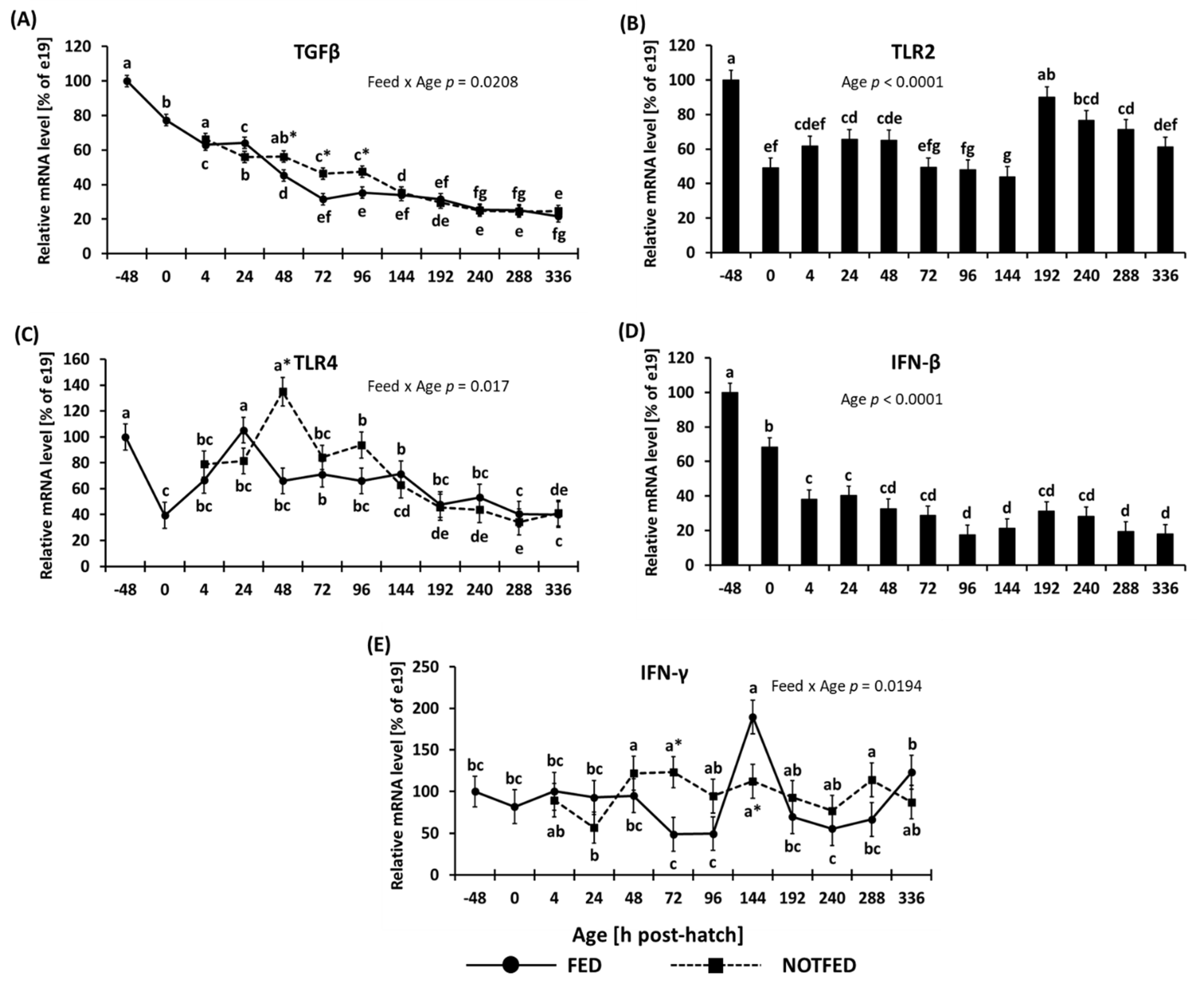
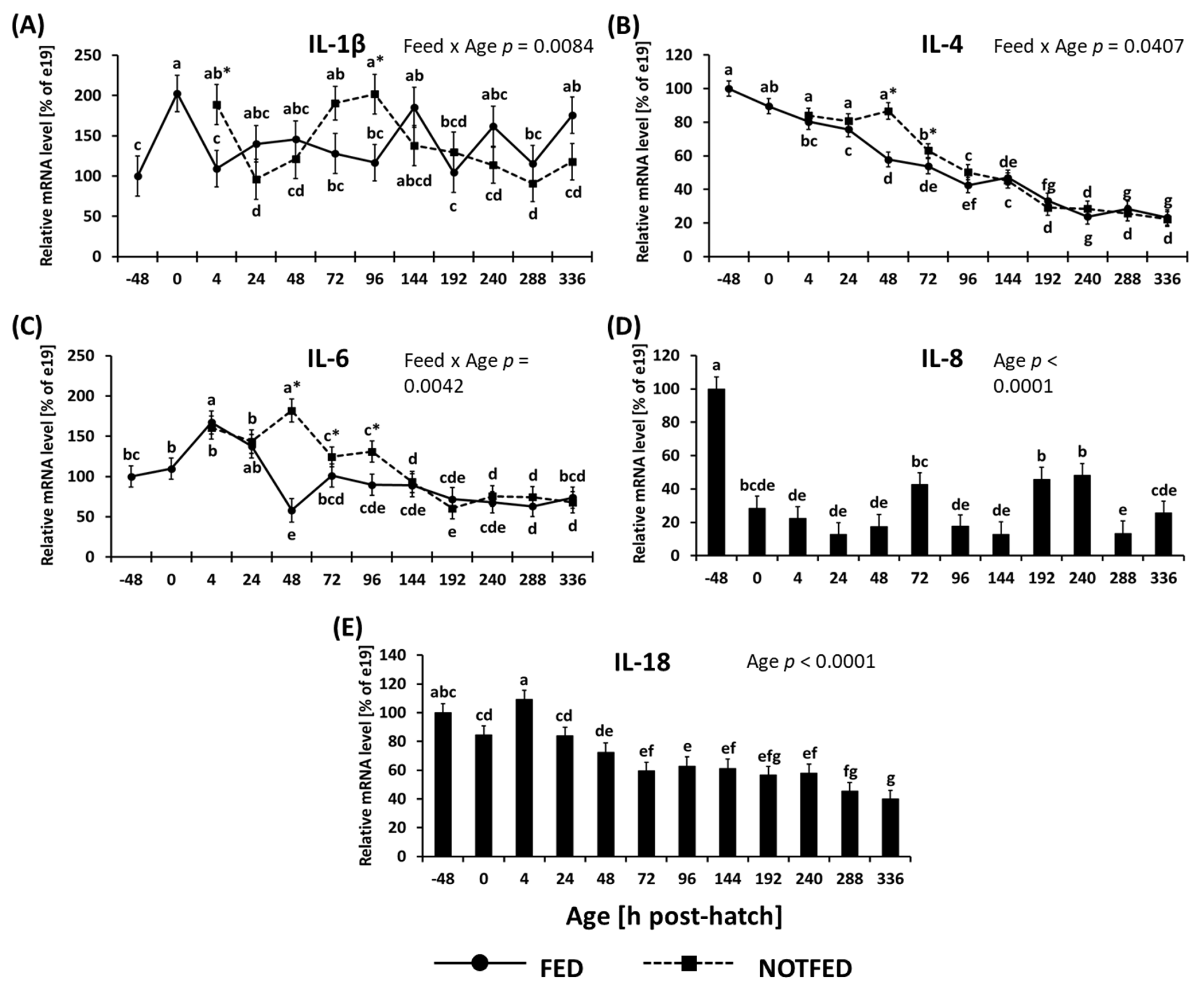
3.1.1. Cytokines and Toll-Like Receptors
3.1.2. Avian Defensins
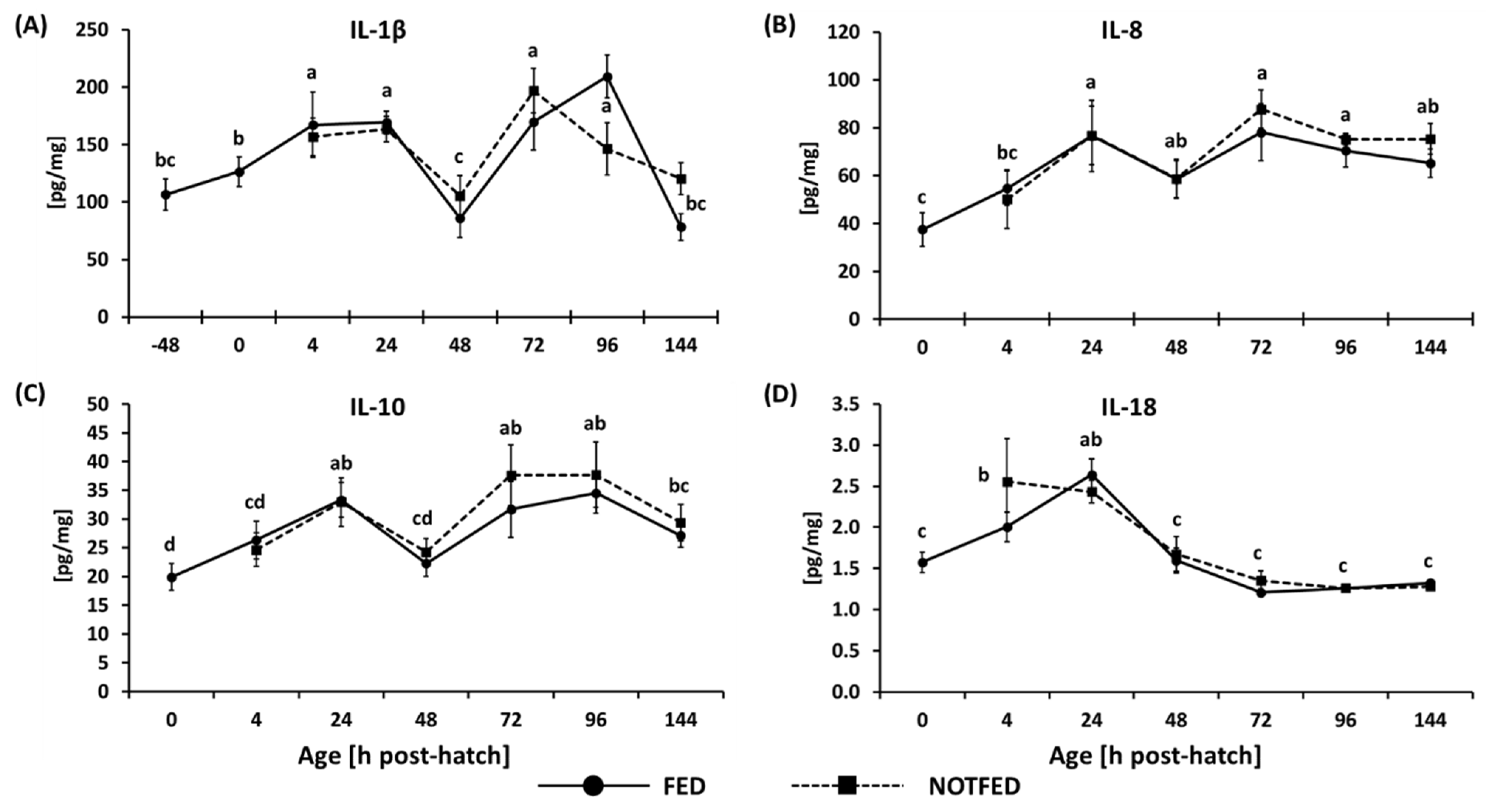
3.2. Protein Expression of Cytokines
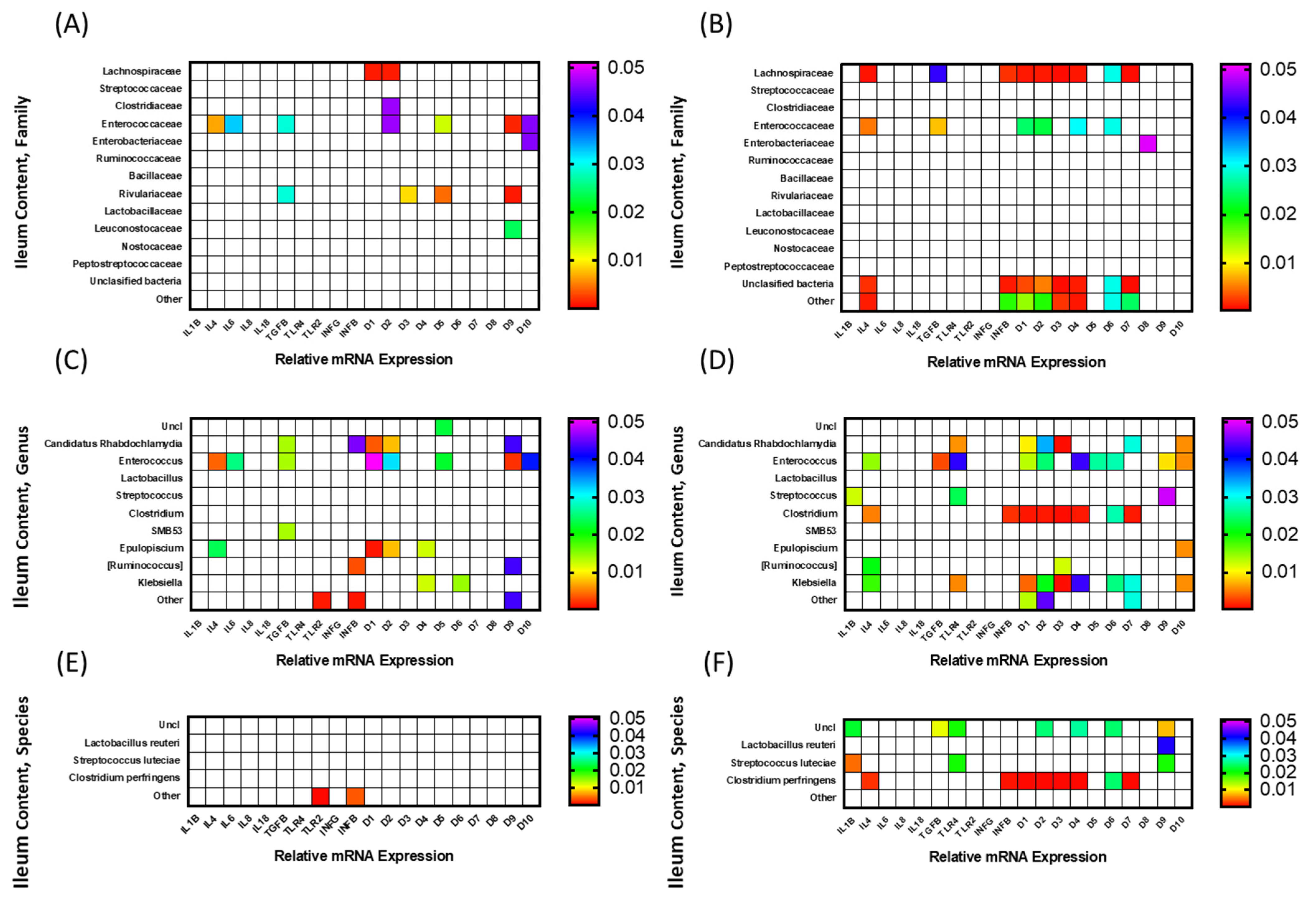
3.3. Correlation among mRNA Expression of Immune-Related Genes and Microbiota Composition
3.3.1. Ileal Luminal Microbiota
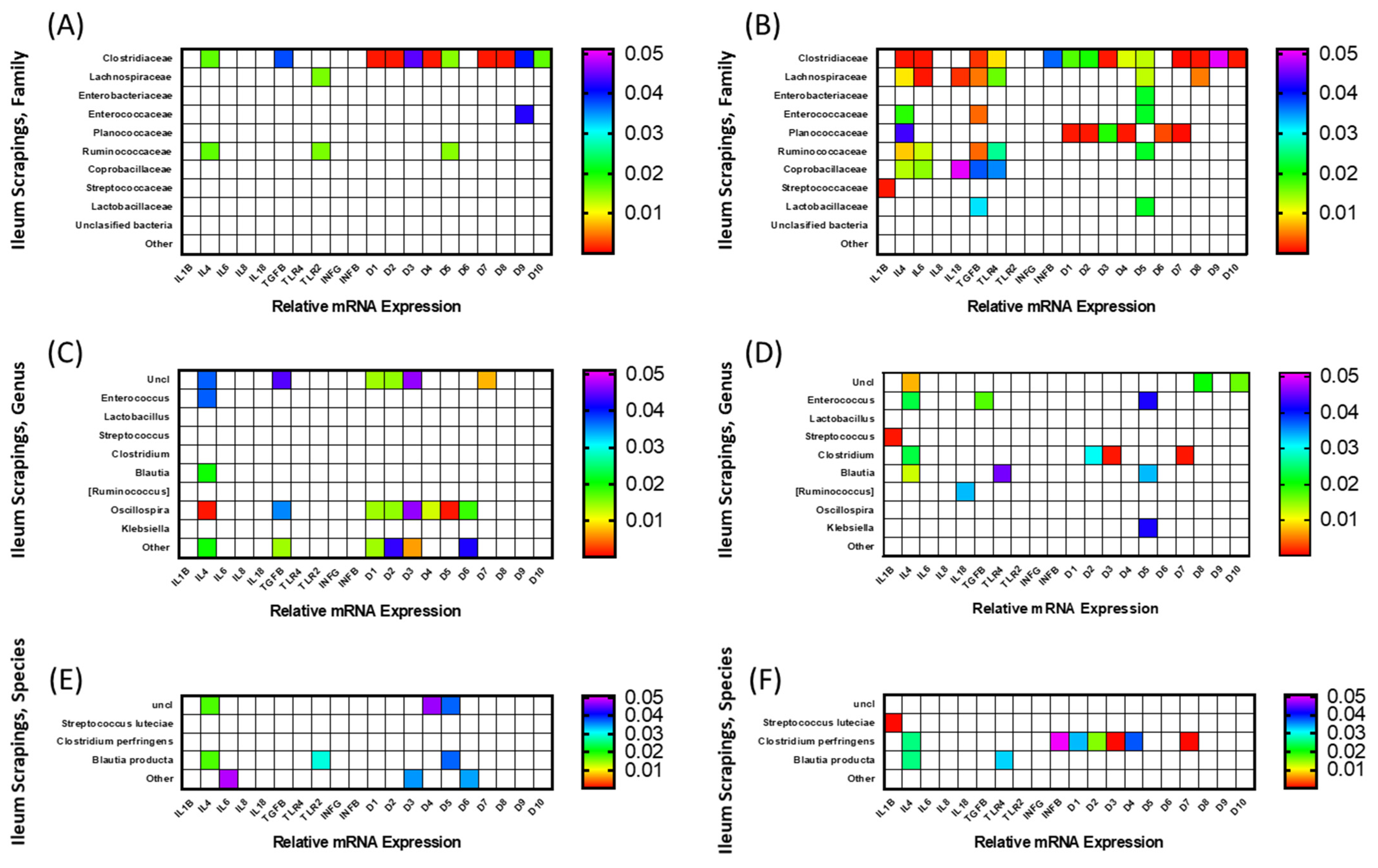
3.3.2. Ileal Scrapings
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noy, Y.; Sklan, D. Yolk utilisation in the newly hatched poult. Br. Poult. Sci. 1998, 39, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Noy, Y.; Sklan, D. Yolk and exogenous feed utilization in the posthatch chick. Poult. Sci. 2001, 80, 1490–1495. [Google Scholar] [CrossRef]
- Careghi, C.; Tona, K.; Onagbesan, O.; Buyse, J.; Decuypere, E.; Bruggeman, V. The effects of the spread of hatch and interaction with delayed feed access after hatch on broiler performance until seven days of age. Poult. Sci. 2005, 84, 1314–1320. [Google Scholar] [CrossRef] [PubMed]
- Bigot, K.; Mignon-Grasteau, S.; Picard, M.; Tesseraud, S.J.P.s. Effects of delayed feed intake on body, intestine, and muscle development in neonate broilers. Poult. Sci. 2003, 82, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, E.; Kondo, N.; Saldanha, E.; Loddy, M.; Careghi, C.; Decuypere, E. Performance and physiological parameters of broiler chickens subjected to fasting on the neonatal period. Poult. Sci. 2003, 82, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Proszkowiec-Weglarz, M.; Schreier, L.L.; Miska, K.B.; Angel, R.; Kahl, S.; Russell, B. Effect of early neonatal development and delayed feeding post-hatch on jejunal and ileal calcium and phosphorus transporter genes expression in broiler chickens. Poult. Sci. 2019, 98, 1861–1871. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Jia, M.; Wong, E. Delayed access to feed affects broiler small intestinal morphology and goblet cell ontogeny. Poult. Sci. 2020, 99, 5275–5285. [Google Scholar] [CrossRef]
- Uni, Z.; Ganot, S.; Sklan, D. Posthatch development of mucosal function in the broiler small intestine. Poult. Sci. 1998, 77, 75–82. [Google Scholar] [CrossRef]
- Geyra, A.; Uni, Z.; Sklan, D. The effect of fasting at different ages on growth and tissue dynamics in the small intestine of the young chick. Br. J. Nutr. 2001, 86, 53–61. [Google Scholar] [CrossRef]
- Dibner, J.; Knight, C.; Kitchell, M.; Atwell, C.; Downs, A.; Ivey, F. Early feeding and development of the immune system in neonatal poultry. J. Appl. Poult. Res. 1998, 7, 425–436. [Google Scholar] [CrossRef]
- Taha-Abdelaziz, K.; Hodgins, D.C.; Lammers, A.; Alkie, T.N.; Sharif, S. Effects of early feeding and dietary interventions on development of lymphoid organs and immune competence in neonatal chickens: A review. Vet. Immunol. Immunopathol. 2018, 201, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Golonka, R.M.; Xiao, X.; Abokor, A.A.; Joe, B.; Vijay-Kumar, M. Altered nutrient status reprograms host inflammation and metabolic health via gut microbiota. J. Nutr. Biochem. 2020, 80, 108360. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Egan, M.; Ryan, C.A.; Boyaval, P.; Dempsey, E.M.; Ross, R.P.; Stanton, C. A good start in life is important—perinatal factors dictate early microbiota development and longer term maturation. FEMS Microbiol. Rev. 2020, 44, 763–781. [Google Scholar] [CrossRef] [PubMed]
- Nishio, J.; Honda, K.J.C.; Sciences, M.L. Immunoregulation by the gut microbiota. Cell. Mol. Life Sci. 2012, 69, 3635–3650. [Google Scholar] [CrossRef]
- Kogut, M.H. The gut microbiota and host innate immunity: Regulators of host metabolism and metabolic diseases in poultry? J. Appl. Poult. Res. 2013, 22, 637–646. [Google Scholar] [CrossRef]
- Yu, K.; Choi, I.; Yun, C.-H. Immunosecurity: Immunomodulants enhance immune responses in chickens. Anim. Biosci. 2021, 34, 321–337. [Google Scholar] [CrossRef]
- Hasselquist, D.; Nilsson, J.-Å. Physiological mechanisms mediating costs of immune responses: What can we learn from studies of birds? Anim. Behav. 2012, 83, 1303–1312. [Google Scholar] [CrossRef]
- Lowenthal, J.W.; Staeheli, P.; Schultz, U.; Sekellick, M.J.; Marcus, P.I. Nomenclature of avian interferon proteins. J. Interferon Cytokine Res. 2001, 21, 547–549. [Google Scholar] [CrossRef]
- Sick, C.; Schultz, U.; Staeheli, P. A family of genes coding for two serologically distinct chicken interferons. J. Biol. Chem. 1996, 271, 7635–7639. [Google Scholar] [CrossRef] [Green Version]
- Digby, M.R.; Lowenthal, J.W. Cloning and expression of the chicken interferon-γ gene. J. Interferon Cytokine Res. 1995, 15, 939–945. [Google Scholar] [CrossRef]
- Haq, K.; Elawadli, I.; Parvizi, P.; Mallick, A.I.; Behboudi, S.; Sharif, S. Interferon-γ influences immunity elicited by vaccines against very virulent Marek’s disease virus. Antivir. Res. 2011, 90, 218–226. [Google Scholar] [CrossRef]
- Garrido, D.; Alber, A.; Kut, E.; Chanteloup, N.K.; Lion, A.; Trotereau, A.; Dupont, J.; Tedin, K.; Kaspers, B.; Vervelde, L. The role of type I interferons (IFNs) in the regulation of chicken macrophage inflammatory response to bacterial challenge. Dev. Comp. Immunol. 2018, 86, 156–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weining, K.C.; Sick, C.; Kaspers, B.; Staeheli, P. A chicken homolog of mammalian interleukin-1β: cDNA cloning and purification of active recombinant protein. Eur. J. Biochem. 1998, 258, 994–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.-T.; Huang, W.-Y.; Chen, T.; Salawu, E.O.; Wang, D.; Lee, Y.-Z.; Chang, Y.-Y.; Yang, L.-W.; Sue, S.-C.; Wang, X. Structure and function of chicken interleukin-1 beta mutants: Uncoupling of receptor binding and in vivo biological activity. Sci. Rep. 2016, 6, 27729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avery, S.; Rothwell, L.; Degen, W.D.; Schijns, V.E.; Young, J.; Kaufman, J.; Kaiser, P. Characterization of the first nonmammalian T2 cytokine gene cluster: The cluster contains functional single-copy genes for IL-3, IL-4, IL-13, and GM-CSF, a gene for IL-5 that appears to be a pseudogene, and a gene encoding another cytokinelike transcript, KK34. J. Interferon Cytokine Res. 2004, 24, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, A.A.; Kim, W.H.; Lillehoj, H.S. Interleukin-4 (IL-4) may regulate alternative activation of macrophage-like cells in chickens: A sequential study using novel and specific neutralizing monoclonal antibodies against chicken IL-4. Vet. Immunol. Immunopathol. 2018, 205, 72–82. [Google Scholar] [CrossRef] [PubMed]
- de Waal Malefyt, R.; Hans, Y.; Roncarolo, M.-G.; Spits, H.; de Vries, J.E. Interleukin-10. Curr. Opin. Immunol. 1992, 4, 314–320. [Google Scholar] [CrossRef]
- Rothwell, L.; Young, J.R.; Zoorob, R.; Whittaker, C.A.; Hesketh, P.; Archer, A.; Smith, A.L.; Kaiser, P. Cloning and characterization of chicken IL-10 and its role in the immune response to Eimeria maxima. J. Immunol. 2004, 173, 2675–2682. [Google Scholar] [CrossRef] [Green Version]
- Schneider, K.; Klaas, R.; Kaspers, B.; Staeheli, P. Chicken interleukin-6: cDNA structure and biological properties. Eur. J. Biochem. 2001, 268, 4200–4206. [Google Scholar] [CrossRef]
- Lynagh, G.R.; Bailey, M.; Kaiser, P. Interleukin-6 is produced during both murine and avian Eimeria infections. Vet. Immunol. Immunopathol. 2000, 76, 89–102. [Google Scholar] [CrossRef]
- Baggiolini, M. Chemokines and leukocyte traffic. Nature 1998, 392, 565–568. [Google Scholar] [CrossRef] [PubMed]
- Sugano, S.; Stoeckle, M.Y.; Hanafusa, H. Transformation by Rous sarcoma virus induces a novel gene with homology to a mitogenic platelet protein. Cell 1987, 49, 321–328. [Google Scholar] [CrossRef]
- Okamura, H.; Tsutsui, H.; Komatsu, T.; Yutsudo, M.; Hakura, A.; Tanimoto, T.; Torigoe, K.; Okura, T.; Nukada, Y.; Hattori, K. Cloning of a new cytokine that induces IFN-γ production by T cells. Nature 1995, 378, 88–91. [Google Scholar] [CrossRef]
- Schneider, K.; Puehler, F.; Baeuerle, D.; Elvers, S.; Staeheli, P.; Kaspers, B.; Weining, K.C. cDNA cloning of biologically active chicken interleukin-18. J. Interferon Cytokine Res. 2000, 20, 879–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Göbel, T.W.; Schneider, K.; Schaerer, B.; Mejri, I.; Puehler, F.; Weigend, S.; Staeheli, P.; Kaspers, B. IL-18 stimulates the proliferation and IFN-γ release of CD4+ T cells in the chicken: Conservation of a Th1-like system in a nonmammalian species. J. Immunol. 2003, 171, 1809–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogut, M.H.; Arsenault, R.J. A role for the non-canonical Wnt-β-catenin and TGF-β signaling pathways in the induction of tolerance during the establishment of a Salmonella enterica serovar enteritidis persistent cecal infection in chickens. Front. Vet. Sci. 2015, 2, 33. [Google Scholar] [CrossRef] [Green Version]
- Moustakas, A.; Souchelnytskyi, S.; Heldin, C.-H. Smad regulation in TGF-β signal transduction. J. Cell Sci. 2001, 114, 4359–4369. [Google Scholar] [CrossRef]
- van Dijk, A.; Veldhuizen, E.J.; Haagsman, H.P. Avian defensins. Vet. Immunol. Immunopathol. 2008, 124, 1–18. [Google Scholar] [CrossRef]
- Chertov, O.; Michiel, D.F.; Xu, L.; Wang, J.M.; Tani, K.; Murphy, W.J.; Longo, D.L.; Taub, D.D.; Oppenheim, J.J. Identification of defensin-1, defensin-2, and CAP37/Aurocidin as T-cell chemoattractant proteins released from interleukin-8-stimulated neutrophils. J. Biol. Chem. 1996, 271, 2935–2940. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wong, E.A. Expression of avian β-defensin mRNA in the chicken yolk sac. Dev. Comp. Immunol. 2019, 95, 89–95. [Google Scholar] [CrossRef]
- Proszkowiec-Weglarz, M.; Schreier, L.L.; Kahl, S.; Miska, K.B.; Russell, B.; Elsasser, T.H. Effect of delayed feeding post-hatch on expression of tight junction- and gut barrier-related genes in the small intestine of broiler chickens during neonatal development. Poult. Sci. 2020, 99, 4714–4729. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Poultry, 9th ed.; National Academies Press: Washington, DC, USA, 1994. [Google Scholar]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S. Primer3—new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, V.L.; Mowbray, C.A.; Cadwell, K.; Niranji, S.S.; Bailey, R.; Watson, K.A.; Ralph, J.; Hall, J. Effects of rearing environment on the gut antimicrobial responses of two broiler chicken lines. Vet. Immunol. Immunopathol. 2016, 178, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Campos, P.M.; Miska, K.B.; Kahl, S.; Jenkins, M.C.; Shao, J.; Proszkowiec-Weglarz, M. Effects of Eimeria tenella on cecal luminal and mucosal microbiota in broiler chickens. Avian Dis. 2022, 66, 39–52. [Google Scholar] [CrossRef]
- Shira, E.B.; Sklan, D.; Friedman, A. Impaired immune responses in broiler hatchling hindgut following delayed access to feed. Vet. Immunol. Immunopathol. 2005, 105, 33–45. [Google Scholar] [CrossRef]
- Bar-Shira, E.; Sklan, D.; Friedman, A. Establishment of immune competence in the avian GALT during the immediate post-hatch period. Dev. Comp. Immunol. 2003, 27, 147–157. [Google Scholar] [CrossRef]
- Eren, U.; Kum, S.; Nazligul, A.; Gules, O.; Aka, E.; Zorlu, S.; Yildiz, M. The several elements of intestinal innate immune system at the beginning of the life of broiler chicks. Microsc. Res. Tech. 2016, 79, 604–614. [Google Scholar] [CrossRef]
- Rehman, M.S.U.; Rehman, S.U.; Yousaf, W.; Hassan, F.U.; Ahmad, W.; Liu, Q.; Pan, H. The potential of Toll-Like Receptors to modulate avian immune system: Exploring the effects of genetic variants and phytonutrients. Front. Genet. 2021, 12, 671235. [Google Scholar] [CrossRef]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-gamma: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef]
- He, H.; Genovese, K.J.; Kogut, M.H. Modulation of chicken macrophage effector function by T(H)1/T(H)2 cytokines. Cytokine 2011, 53, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Konkel, J.E.; Chen, W. Balancing acts: The role of TGF-β in the mucosal immune system. Trends Mol. Med. 2011, 17, 668–676. [Google Scholar] [CrossRef] [Green Version]
- Simon, K.; de Vries Reilingh, G.; Kemp, B.; Lammers, A. Development of ileal cytokine and immunoglobulin expression levels in response to early feeding in broilers and layers. Poult. Sci. 2014, 93, 3017–3027. [Google Scholar] [CrossRef]
- Lamot, D.; Van der Klein, S.; Van De Linde, I.; Wijtten, P.; Kemp, B.; Van Den Brand, H.; Lammers, A. Effects of feed access after hatch and inclusion of fish oil and medium chain fatty acids in a pre-starter diet on broiler chicken growth performance and humoral immunity. Animal 2016, 10, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Tang, D.; Yan, S.; Fan, H.; Li, G.; Shahid, M.S.; Mahmood, T.; Guo, Y. Effects of age on immune function in broiler chickens. J. Anim. Sci. Biotechnol. 2021, 12, 42. [Google Scholar] [CrossRef] [PubMed]
- Yacoub, H.A.; Elazzazy, A.M.; Abuzinadah, O.A.; Al-Hejin, A.M.; Mahmoud, M.M.; Harakeh, S.M. Antimicrobial activities of chicken β-defensin (4 and 10) peptides against pathogenic bacteria and fungi. Front. Cell. Infect. Microbiol. 2015, 5, 36. [Google Scholar] [CrossRef]
- Lee, M.O.; Jang, H.-J.; Rengaraj, D.; Yang, S.-Y.; Han, J.Y.; Lamont, S.J.; Womack, J.E. Tissue expression and antibacterial activity of host defense peptides in chicken. BMC Vet. Res. 2016, 12, 231. [Google Scholar] [CrossRef] [Green Version]
- Terada, T.; Nii, T.; Isobe, N.; Yoshimura, Y. Changes in the expression of avian β-defensins (AvBDs) and proinflammatory cytokines and localization of AvBD2 in the intestine of broiler embryos and chicks during growth. J. Poult. Sci. 2018, 55, 0180022. [Google Scholar] [CrossRef] [Green Version]
- Lyu, W.; Zhang, L.; Gong, Y.; Wen, X.; Xiao, Y.; Yang, H. Developmental and Tissue Patterns of the Basal Expression of Chicken Avian β-Defensins. Biomed Res. Int. 2020, 2020, 2567861. [Google Scholar] [CrossRef]
- Meade, K.G.; O’Farrelly, C. B-Defensins: Farming the microbiome for homeostasis and health. Front. Immunol. 2019, 9, 3072. [Google Scholar] [CrossRef] [PubMed]
- Hofacre, C.L.; Smith, J.A.; Mathis, G.F. An optimist’s view on limiting necrotic enteritis and maintaining broiler gut health and performance in today’s marketing, food safety, and regulatory climate. Poult. Sci. 2018, 97, 1929–1933. [Google Scholar] [CrossRef] [PubMed]
- Broom, L.J.; Kogut, M.H. The role of the gut microbiome in shaping the immune system of chickens. Vet. Immunol. Immunopathol. 2018, 204, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Hughes, R.J.; Geier, M.S.; Moore, R.J. Bacteria within the gastrointestinal tract microbiota correlated with improved growth and feed conversion: Challenges presented for the identification of performance enhancing probiotic bacteria. Front. Microbiol. 2016, 7, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The Genus Enterococcus: Between Probiotic Potential and Safety Concerns-An Update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Luo, L.; Zhang, Y.; Wang, Z.; Xia, Z. Effects of the Dietary Probiotic, Enterococcus faecium NCIMB11181, on the Intestinal Barrier and System Immune Status in Escherichia coli O78-Challenged Broiler Chickens. Probiotics Antimicrob. Proteins 2019, 11, 946–956. [Google Scholar] [CrossRef] [Green Version]
- Shehata, A.M.; Paswan, V.K.; Attia, Y.A.; Abdel-Moneim, A.E.; Abougabal, M.S.; Sharaf, M.; Elmazoudy, R.; Alghafari, W.T.; Osman, M.A.; Farag, M.R.; et al. Managing gut microbiota through in ovo nutrition influences early-life programming in broiler chickens. Animals 2021, 11, 3491. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene b | GenBank AccessionNo. c | Forward Primer (5′→3′) | Reverse Primer (5′→3′) | Amplicon Size (bp) |
|---|---|---|---|---|
| IL-1β | NM_204524 | GCATCAAGGGCTACAAGCTC | CAGGCGGTAGAAGATGAAGC | 131 |
| IL-4 | NM_001007079 | GAATGACATCCAGGGAGAGG | AACAATTGTGGAGGCTTTGC | 112 |
| IL-6 | NM_204628 | GGCATTCTCATTTCCTTCTAGG | CCGTAAGAAATGTAACAGGTGTTTT | 135 |
| IL-8 | DQ393272 | ATGTGAAGCTGACGCCAAG | GGCCATAAGTGCCTTTACGA | 131 |
| IL-18 | NM_204608 | TGAAATCTGGCAGTGGAATG | CAACCATTTTCCCATGCTCT | 144 |
| TGF-β | NM_001318450 | CGACCTCGACACCGACTACT | CCACTTCCACTGCAGATCCT | 135 |
| TLR2 | NM_204278 | TCACAGGCAAAATCACGGTG | GATTTGGTTGGACTGCAGCA | 116 |
| TLR4 | NM_001030693 | TTCCTGCTGAAATCCCAAAC | TATGGATGTGGCACCTTGAA | 132 |
| IFN-β | NM_001024836 | GTGCTTGTACCTGGGACCAT | GGATGAGGCTGTGAGAGGAG | 107 |
| IFN-γ | NM_205149 | GCCGCACATCAAACACATATCT | TGAGACTGGCTCCTTTTCCTT | 207 |
| AvBD1 | NM_204993 | TGTGCATTTCTGAAGTGCCC | TTGGGATGTCTGGCTCTTCA | 104 |
| AvBD2 | NM_204992 | GTTCCGTTCCTGCTGCAAAT | TGAGAGGGGTCTTCTTGCTG | 133 |
| AvBD3 | NM_204650 | GATTCTGTCGTGTTGGGAGC | TCCTCACAGAATTCAGGGCA | 114 |
| AvBD5 | NM_001001608 | ATTACCCCAGGACTGTGAGC | ACGTGAAGGGACATCAGAGG | 147 |
| AvBD6 | NM_001001193 | GCCCTACTTTTCCAGCCCTA | CCTGTTCCTCACACAGCAAG | 133 |
| AvBD7 | NM_001001194 | CTCTTGCTGTGCAAGGGGAT | GGAGTGCCAGAGAAGCCATT | 91 |
| AvBD8 | NM_001001781 | ATGCTCCAAGGATCACTGCT | CTGCTTAGCTGGTCTGAGGT | 122 |
| AvBD9 | NM_001001611 | GACGCTGACACCTTAGCATG | CCCATTTGCAGCATTTCAGC | 118 |
| AvBD10 | NM_001001609 | CACTTTTCCCTGACACCGTG | AAAGCCTTTCCTTACTGCGC | 148 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miska, K.B.; Kahl, S.; Schreier, L.L.; Russell, B.; Kpodo, K.R.; Proszkowiec-Weglarz, M. Delay of Feed Post-Hatch Causes Changes in Expression of Immune-Related Genes and Their Correlation with Components of Gut Microbiota, but Does Not Affect Protein Expression. Animals 2022, 12, 1316. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101316
Miska KB, Kahl S, Schreier LL, Russell B, Kpodo KR, Proszkowiec-Weglarz M. Delay of Feed Post-Hatch Causes Changes in Expression of Immune-Related Genes and Their Correlation with Components of Gut Microbiota, but Does Not Affect Protein Expression. Animals. 2022; 12(10):1316. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101316
Chicago/Turabian StyleMiska, Katarzyna B., Stanislaw Kahl, Lori L. Schreier, Beverly Russell, Kouassi R. Kpodo, and Monika Proszkowiec-Weglarz. 2022. "Delay of Feed Post-Hatch Causes Changes in Expression of Immune-Related Genes and Their Correlation with Components of Gut Microbiota, but Does Not Affect Protein Expression" Animals 12, no. 10: 1316. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101316